

 PDF
PDF ePub
ePub Citation
Citation Print
Print



서론
비만은 전세계적으로 증가추세에 있으며 세계보건기구에서 새로운 유행병으로 규정하기에 이르렀다 [1]. 비만의 원인은 유전과 신체활동 저하, 서구화된 식생활 등의 환경적 요인에 기인되는데 개인 차원의 건강문제 뿐 아니라 비만인구가 늘어나게 되면 국가 차원의 사회경제적 비용이 증가되기 때문에 국가차원에서의 비만 관리를 위한 다각적 노력이 필요하다. 비만은 비만 그 자체뿐만 아니라 비만으로 야기될 수 있는 사회적 장애와, 2차적인 합병증인 심혈관계 질환이나 당뇨, 지방간, 관절이상, 암의 발생 비율을 현저하게 증가시킬 수 있기 때문에 전세계적으로 비만 예방과 치료를 위한 많은 다양한 연구에 관심이 모아지고 있다 [23].
비만인구의 감소를 유도하기 위하여 운동과 생활습관과 관련된 많은 정책이 시행되고 있으나, 비만율은 지속적으로 증가되고 있는 실정이다. 비만 유병률을 낮추기 위해서는 신체활동을 늘리고 식생활의 개선뿐만 아니라 체지방 감소에 유용한 생리활성 물질을 함유한 식물자원의 활용도 중요하다. 이에 체지방 감소에 도움을 주는 기능성 성분에 대한 천연물 연구가 활발히 진행되고 있다. 그 중 강력한 항산화물질인 플라보노이드를 다량 함유한 베리류, 감귤류, 코코아, 녹차 등에 관한 연구들이 진행 중에 있다[456]. 베리류의 일종인 댕댕이나무 (Lonicera caerulea) 열매는 허니베리로 많이 알려진 열매로 보라색의 납작한 모양으로 높은 산의 초지 또는 숲 가장자리에 서식하는데 러시아, 중국 만주 지방과 일본, 몽골 등지에서 볼 수 있고 전통 의료 목적으로도 사용되어 왔다 [7]. 국내에서는 강원도와 함경도, 그리고 한라산 능선 지역에서 생산되고 있다. 댕댕이나무에 관한 연구로는 항산화 활성 및 성분 분석에 대한 연구가 진행 되었고 [89] 알코올성 손상 간기능 개선 효과에 대한 연구 [10] 면역 [11], 항암 [12]등에 효과적이며 특히 항산화 효과는 방사선 치료 시 회복에 도움을 주는 것으로 보고되었다 [1314]. 따라서 댕댕이나무 열매는 질병의 예방뿐만 아니라 치료 측면까지 활용 가능할 것으로 사료된다.
항비만 in vitro 실험으로 지방전구세포인 3T3-L1이 많이 사용되고 있으나 최근 연구에서는 다양한 조직으로 분화할 수 있는 지방유래줄기세포 (ex vivo)를 이용하여 갈색지방으로 의 분화 유도에 관한 연구가 진행되고 있다[15]. 지방유래줄기세포는 많은 양을 얻을 수 있고 안정적이며 분화 유도 시 다양한 세포로의 분화가 가능하기 때문에 세포 치료용으로의 가능성도 높아지고 있다 [16]. 지방전구세포에 isobutylmethylxanthine (IBMX), dexamethasone(DEX), insulin을 처리하면 지방세포로 분화가 진행되는데[17], 지방조직의 분화는 다른 조직세포분화와는 달리 여러 호르몬과 다양한 전사인자들의 상호작용을 통해 이루어진다 [18]. 지방전구세포가 지방세포로 분화되면 지방세포 특이적인 유전자들의 활성과 함께 지방구 형성을 촉진하여 세포 내 지방의 축적을 초래하게 된다 [18].
연구방법
실험 재료
댕댕이나무 열매는 강원도 정선에서 5월 말에 수확한 열매를 구입하여 사용하였고, 시료를 블랜더 (Hanil, Seoul, Korea)로 파쇄하여 60℃ 수조 (Chang Shin Co., Seoul, Korea)에서 에탄올 25%가 함유된 증류수로 3시간 동안 환류 추출하였다. 용매 추출액은 여과지 No. 2 filter paper (Toyo Roshi Kaisha, Tokyo, Japan)로 필터하여 60–65℃의 온도에서 회전 농축기 (EYELA, N-N series, Shanghai, China)에서 감압 농축시켜 용매를 완전히 제거하여 시료를 얻었다. 얻어진 시료는 −20℃에서 보관하였다. 수율은 추출 전 시료 중량에 대한 추출 후 농축액의 백분율로, 25% 에탄올 추출물에서 14.44%로 나타났다.
3T3-L1 지방전구세포
3T3-L1 지방전구세포는 American Type Culture Collection (Manassas, VA, USA)에서 구입하였다. 10% bovine calf serum (BCS), 1% penicillin 및 streptomycin (Gibco BRL, NY, USA)이 함유된 DMEM을 사용하여 37℃, 5% CO2 조건에서 배양하였다. 세포수의 증식에 따른 과밀도 현상을 해소하기 위하여 48시간마다 배지 교환을 실시하여 적정수의 세포를 유지하고 지방세포 분화 및 지방 관련 바이오마커로 peroxisome proliferator-activated receptor-gamma(PPARγ), CCAAT/enhancer-binding protein-alpha (C/EBPα), sterol regulatory element-binding transcription factor 1(SREBP1)를 측정하였다.
지방에서 추출한 지방유래줄기세포 (mouse adiposederived stem cells, MADSCs)
지방유래줄기세포는 4주령의 C57BL/6 mouse를 구입하여 경추 탈골 후 지방 (subcutaneous adipose tissue (SAT), epididymal adipose tissue (EAT)와 retroperitoneal adipose tissue (RAT))을 채취하여 사용하였다. 채취한 지방조직은 collagenase II를 처리하여 지방분해 후 원심 분리하여 stromal vascular fraction (SVF)를 획득 후 10% BCS를 첨가한 DMEM에 풀어 10 mm dish에 37℃, 5% CO2 조건에서 배양하였다. 이틀 후 6 well plate에 well당 1 × 105 cells의 농도로 분주 후 다음 날 분화를 유도하였다.
동물실험은 가천대학교의 심의·승인을 획득 (GIACUCR2018016) 후, 실험을 진행하였다.
세포독성 평가 (CCK-8 assay)
지방전구세포인 3T3-L1 cell을 10% BCS가 포함된 DMEM 배지에 2 × 104 cells/well로 96-well plate에 well 당 100 µL씩 분주하였다. 24시간 후 LCE를 농도별 (0, 0.05, 0.5, 1.25, 2,5, 5, 10 mg/mL)로 처리한 뒤 각각 24, 48, 72시간 동안 배양하였다. 각 well에 CCK-8 용액 10 µL를 가하고 알루미늄 호일로 밀폐하여 2시간 동안 다시 배양한 후 450 nm의 파장에서 흡광도를 측정하였다. 모든 실험은 3회이상 반복하였다.
세포분화 유도
세포를 1 × 105 cells/well로 희석시켜 6 well plate (SPL, Korea)에 분주한 후 세포가 80% 경과 시 0.5 mM IBMX(Sigma, Missouri, USA), 1 µM Dex (Sigma, Missouri, UAS), 5 µg/mL Insulin (Sigma, Missouri, USA)이 포함된 DMEM (10% FBS, 1% penicillin/streptomycin)으로 분화를 유도하였고 3일후 5 µg/mL insulin이 포함된 DMEM(10% FBS, 1% penicillin/streptomycin)으로 교체하였다. 세포분화 유도기간은 총 7일로 실험군은 분화 시작부터 마지막까지 분화유도 medium내 LCE각각의 농도 (0, 0.25, 0.5, 1 mg/mL)로 처리하였고, 분화만 유도하고 LCE를 처리하지 않은 군 (0 mg/mL)을 대조군 (Control)으로 삼았다. 같은 실험을 3회이상 반복하였다.
Oil Red O 염색 및 지방축적 평가
LCE를 농도별로 함께 처리하여 분화시킨 세포에 4% formaldehyde를 처리하여 30분간 상온에서 고정하였다. 고정한 세포에 0.4 μm filter로 여과한 Oil Red O 염색시약을 20분간 처리한 뒤 증류수로 Oil Red O 염색시약을 충분히 세척하고 위상차 도립현미경 (Nikon Eclipse microscope, Tokyo, Japan)에서 × 100배로 확대하여 사진촬영을 하였다. 사진촬영 후 isopropanol을 처리하여 염색된 세포 지방구에서 Oil Red O 염색약을 추출한 뒤 500 nm에서 흡광도를 측정하였다. 동일한 실험을 3회이상 반복하여 측정하였다.
지방 분화 관련 유전자의 mRNA 발현량 측정
LCE를 농도별 함께 처리하여 분화시킨 세포를 PBS로 2회 세척한 후 세포를 수득하여 RNA extraction kit (17221, iNtRON Biotechnology, Gyeonggi-do, Korea)을 이용하여 total RNA를 분리하였다. RNA 50 ng을 cDNA로 합성한 후 SYBR Green Master Mix (TaKaRa Bio, Otsu, Japan)를 사용하여 ABI QuantStudio 3 (Applied Biosystems, California, USA) 장비로 qRT-PCR 분석을 수행하였다. 프라이머는 PPARγ, C/EBPα, and SREBP1 특이적인 프라이머를 사용하였다 (Table 1). 모든 실험은 3회 이상 반복 실험하였다.
지방 분화 유전자들의 단백질 발현량 측정
분화가 완료된 세포를 ice-cold 1xPBS로 두 번 세척한 후 PRO-PREP Protein Purification Kit (iNtRON, Gyeonggi-do, Korea)를 넣어 세포를 용해시켰다. 단백질 농도는 BCA Protein Assay Kit (Thermo Fisher Scientific, Massachusetts, USA)로 측정하였고, 세포용해액들을 단백질량이 30 µg 되도록 10% SDS-PAGE gel에 loading하여 electrophoresis 하여 세포 내 단백질을 분리하였으며, PVDF western blotting membrane (Life Science, CA, USA) 에 부착하였다. PPAR γ, SREBP, C/EBPα, β-actin의 1차 항체는 abcam (Abcam, Cambridge, USA)에서 구입하여 사용하였고 4℃에서 밤새 incubation 한 후, horseradish peroxidase-conjugated 2차 항체 (Promega, Wisconsin, USA)들을 부착시켜 발생되는 enhanced chemiluminescence를 ImageQuant LAS500 (GE Healthcare, Massachusetts, USA)로 하였다. 모든 실험은 3회 이상 반복 실험하였다.
결과
세포독성평가
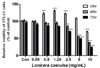
LCE를 3T3-L1 세포에 처리하여 세포 독성과 세포 증식능을 확인하였다. 24, 48, 72시간이 경과하여도 1.25 mg/mL 이하의 농도에서 세포 독성은 관찰되지 않았고 80% 이상의 생존율을 보였다. 5 mg/mL 이상의 농도에서는 48시간 이후로 세포증식능이 급격히 떨어졌다. 그러나 24시간 경과 시에는 5 mg/mL이하의 농도에서 세포 분화가 활발히 일어난 것을 관찰하였다 (Fig. 1). 위의 결과를 바탕으로 실험에 사용할 농도는 1 mg/mL이하인 0.25, 0.5, 1 mg/mL으로 진행하였다.
Oil Red O 염색 및 지방축적 평가
지방전구세포 와 지방유래줄기세포의 분화 및 lipid droplet 생성 정도를 확인하기 위하여 LCE를 0, 0.25, 0.5와 1 mg/mL로 희석 처리하여 분화를 유도한 후 Oil Red O 염색을 실시하였고, Fig. 2 & 3과 같이 댕댕이나무 열매 추출물을 처리하지 않고 지방세포분화를 유도한 대조군(Con; 0 mg/mL)에 비해 LCE를 처리한 군에서 농도의존적으로 지방구 형성이 억제되는 것을 관찰할 수 있었다. Fig. 2A는 3T3-L1 지방전구세포의 LCE 농도별 지방분화유도 결과이다. 분화를 시키지 않은 Pre군에 비해 Con군은 세포 크기가 커지고 Oil Red O 염색에 의해 빨갛게 염색된 지방구를 확인할 수 있다. 이 지방구는 LCE농도가 높아질수록 지방구의 크기가 작아지며 색깔도 옅어지는 것을 볼 수 있다. 이는 흡광도를 측정한 결과 (Fig. 2B)에서 더욱 더 잘 나타난다. 특히 LCE를 처리하지 않고 지방 분화를 유도한 대조군에 비해 1 mg/mL 농도에서 lipid droplet 형성이 가장 많이 억제되어 82% 수준으로 유의미한 결과를 얻었다 (p < 0.001). Fig. 3은 마우스 지방에서 MADSCs를 분화시켜 얻은 결과이다. 3T3-L1 지방전구세포의 결과와 마찬가지로 분화 유도 후 LCE농도 의존적으로 지방구의 크기가 감소하고 색깔이 옅어졌다 (Fig. 3A). 흡광도 측정 결과 모든 농도에서 p < 0.001의 유의미한 결과를 얻었다(Fig. 3B). 이 결과는 LCE는 3T3-L1과 지방유래줄기세포의 지방 분화유도 시 지방구의 크기와 색깔 감소에 영향을 미치는 것으로 확인되었다.
지방 분화 유전자들의 단백질 발현량 측정
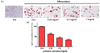
3T3-L1 뿐만 아니라 MADSCs에서도 LCE를 농도별로 처리하면서 지방세포분화를 유도하였다. Fig. 5의 western blot의 결과에서 보는 것처럼 LCE를 처리하지 않은 대조군은 PPARγ, CEBP/α, SREBP-1의 발현이 증가하였다. LCE를 농도별로 처리하였을 때 유의성 있게 감소하는 것을 3T3-L1 세포뿐만 아니라 MADSCs에서도 확인하였다. 대조군에 비해 LCE의 농도의존적으로 MADSCs에서의 PPARγ, C/EBPα, SREBP1 단백질 발현은 모두 감소하였고 SREBP1 단백질에 비해 PPARγ, C/EBPα 단백질의 발현은 눈에 띄게 줄어든 것을 알 수 있다 (Fig. 5A, B, and D). PPARγ, C/EBPα, SREBP1 단백질 발현이 증가할수록 지방세포 분화가 촉진되며 지방산 대사관련 유전자의 발현이 증가하게 된다 [2223]. 위의 결과로 보아 MADSCs에서 LCE처리군은 농도의존적으로 지방세포 분화를 억제하는 것으로 나타났다.
고찰
비만과 그에 따른 대사성 질환의 발병률이 전세계적으로 심각한 보건 문제로 대두되고 있고 국내에서도 최근 들어 일반적인 비만보다는 고도비만과 초고도비만을 중심으로 일어나고 있다 [24]. 비만은 단순한 과체중과는 달리 체내 지방의 과잉축적을 의미하며 세포수의 변화없이 세포가 커지는 세포 비대형 비만 (hypertrophic obesity)과 지방세포 수와 크기가 모두 같이 증가한 세포 과다 증식 및 세포 비대형 비만 (hyperplastic-hypertrophic obesity)등이 있다 [25]. 비만은 지방전구세포가 지방세포로 분화되는 정상적인 지방세포의 발달 이상 뿐만 아니라 과잉분화와 지방세포 내 지방의 축적 과잉으로 인해 비만 증상이 더욱 심해지는 것으로 알려져 있다 [2].
3T3-L1은 지방전구세포로 성장이 지속되며 배양액에 첨가한 분화유도인자에 의해 성장이 멈추고 지방세포로 분화가 이루어지게 된다 [26]. 분화 시 포유류의 지방조직 성장과 생화학적, 생태학적으로 유사하기 때문에 비만에 대한 효과 검증에 주로 사용된다 [27]. 뿐만 아니라 최근에는 마우스 지방조직에서 직접 추출한 줄기세포도 활발하게 이용되고 있다 [2829]. 지방은 많은 양의 조직 채취가 용이하여 줄기세포를 얻기에 유리하고 지방유래줄기세포는 배양 시 안정적인 성장과 증식이 가능하고 분화를 유도하였을 때 골수줄기세포와 같이 지방세포, 연골, 신경 등 다양한 세포로의 분화가 가능하다 [1630]. 지방조직에서 지방을 분리하여 성숙한 지방세포, 적혈구 등을 제거한 나머지 기질세포 (stromal vascular fraction, SVF)를 분리시켜 지방세포 분화실험에 활용하고 있다 [31].
지방세포의 분화 조절은 PPARγ, C/EBPα, SREBP1라고 불리는 세가지 전사인자가 중추적인 역할을 담당하고 있다. 이들 세가지 전사인자는 지방세포 분화 과정 중 각기 다른 시점에서 발현이 유도되고 상호작용을 통해 지방세포 분화를 점진적으로 유도해 나간다 [32]. PPARγ는 지방세포분화와 지방 대사조절에서 중요한 기능을 담당하고 있으면서 PPARγ의 발현 증가는 C/EBPα의 지방세포 분화 후기 발현을 유도한다 [33]. SREBP1은 지방전구세포의 분화 초기에 매우 빠르게 유도되는 전사인자로서, PPARγ와 함께 지방세포 분화를 촉진시키는 역할을 하며 지방 대사에 관련된 많은 유전자의 발현을 증가시키는 것으로 알려져 있다 [22]. 지방조직의 분화는 다양한 전사인자들의 상호작용을 통해 복잡하게 이뤄지는데, 지방전구세포의 지방 분화 시에는 유전자 발현 양상의 변화 뿐만 아니라 세포의 형태적인 변화 또한 동시에 일어난다 [34].
댕댕이나무 (Lonicera caerulea)는 인동과 인동덩굴속에 속하는 낙엽관목으로 북유럽, 북아메리카, 북동아시아 등에 분포하며 9 ~ 60여종에 이른다 [3536]. 댕댕이나무 열매는 blue honeysuckle로 불리우나 honey berry, sweetberry, honeysuckle, edible honeysuckle로 불리기도 한다 [37]. 열매는 당뇨병이나 심혈관질환과 같은 질환을 예방하는 효과와 함께 항산화 작용을 도와준다 [13]. 간 기능 개선에 효과를 보이는 것으로 나타났으며, 베리류의 항염증 효과 등에 관한 연구는 보고된 바가 있다 [38]. 하지만, 댕댕이나무 열매를 이용한 항비만 관련 연구는 미흡하며 현재까지 지방전구세포의 분화 및 지방 생성 억제에 관한 연구는 거의 없는 실정이다.
3T3-L1세포에 댕댕이나무 열매 추출물을 7일간 처리 후 8일째 지방 분화관련 전사인자들 및 그에 따른 단백질 발현을 살펴본 결과, PPARγ, C/EBPα, SREBP1의 mRNA발현과 단백질 발현 모두 농도 의존적으로 대조군에 비해 댕댕이나무 열매 추출물을 처리한 군에서 감소하였는데 이는 댕댕이나무 열매 추출물을 처리한 마우스 지방유래줄기세포에서도 동일한 결과를 얻었다. 이러한 결과는 지방분화에 핵심적인 PPARγ, C/EBPα, SREBP1의 mRNA발현 저해가 단백질 발현 저해를 일으키고 나아가 3T3-L1과 마우스 지방유래줄기세포내 지방구 형성을 억제시키는 결과를 일으킨 것으로 사료된다. 즉, 댕댕이나무 열매 추출물이 3T3-L1세포에서 지방 합성 전사인자 및 단백질 발현을 감소시켜 지방구 크기를 감소시키는 것으로 나타났는데 이는 다른 베리류의 지방합성 억제 효과 연구 결과들과 비슷하였다 [53940]. 본 연구의 지방세포분화 감소 및 억제는 댕댕이나무 열매 추출물이 다량으로 함유하고 있는 안토시아닌의 효과인 것으로 사료된다 [20].
3T3-L1세포에 댕댕이나무 열매 추출물을 7일간 처리 후 8일째 지방 분화관련 전사인자들 및 그에 따른 단백질 발현을 살펴본 결과, PPARγ, C/EBPα, SREBP1의 mRNA발현과 단백질 발현 모두 농도 의존적으로 대조군에 비해 댕댕이나무 열매 추출물을 처리한 군에서 감소하였는데 이는 댕댕이나무 열매 추출물을 처리한 마우스 지방유래줄기세포에서도 동일한 결과를 얻었다. 이러한 결과는 지방분화에 핵심적인 PPARγ, C/EBPα, SREBP1의 mRNA발현저해가 단백질 발현 저해를 일으키고 나아가 3T3-L1과 마우스 지방유래줄기세포내 지방구 형성을 억제시키는 결과를 일으킨 것으로 사료된다. 즉, 댕댕이나무 열매 추출물이 3T3-L1세포에서 지방 합성 전사인자 및 단백질 발현을 감소시켜 지방구 크기를 감소시키는 것으로 나타났는데 이는 다른 베리류의 지방합성 억제 효과 연구 결과들과 비슷하였다 [53940]. 본 연구의 지방세포분화 감소 및 억제는 댕댕이나무 열매 추출물이 다량으로 함유하고 있는 안토시아닌의 효과인 것으로 사료된다 [20].
지방분화에 대한 안토시아닌의 효과에 대해서는 최근 많이 연구가 되고 있다. Kim 등 [41]에 따르면 검정콩의 안토시아닌이 3T3-L1의 분화 과정 중 지방 축적과 지방분화에 관련된 주요한 유전자인 PPARγ의 발현을 억제시킨다고 발표하였고 산딸나무 (Cornus kousa)잎에서 에탄올로 추출한 안토시아닌이 3T3-L1의 지방 분화 뿐만 아니라 지방질 합성에 관여하는 유전자들의 발현을 조절하여 지방 축적을 저해시키는 효과에 대해 보고하였다. 안토시아닌이 풍부한 분획물이 AMPK의 인산화과정을 증가시킨다고 보고하였고 이는 안토시아닌의 antiangiogenic activities와도 연관이 있는 것으로, 비만을 조절할 수 있는 기능식품으로 산딸나무를 제안하였다 [42].
본 연구에서는 머루에서 추출한 안토시아닌이 지방세포의 분화를 억제한다는 연구 [5]에서와 유사하게, 댕댕이나무 열매 추출물을 처리한 모든 농도에서 지방구의 형성이 억제된 것으로 나타났다. 지방 분화가 완료된 3T3-L1뿐만 아니라 마우스 지방유래줄기세포에서 현미경 사진으로 보이는 빨간 지방구들이 농도의존적으로 작아지거나 옅어지는 것을 확인하였는데, 이는 흡광도를 측정하였을 때와 같이 농도의존적으로 감소한 것으로 나타났다. 결론적으로, 이는 지방세포분화에 있어서 반드시 필요한 전사인자들의 발현을 댕댕이나무 열매 추출물이 저해시켜 이로 인하여 단백질 발현이 낮아지고 지방구 형성을 억제 및 감소시키는 것으로 사료된다.
요약
본 연구에서는 댕댕이나무 열매 추출물이 3T3-L1과 마우스 지방유래줄기세포의 지방 분화유도 및 지방생성에 미치는 영향을 살펴보았다. 3T3-L1에 댕댕이나무 열매 추출물을 처리하였을 때, 농도의존적으로 지방구의 생성을 줄였고 지방세포 분화에 있어서 중요한 전사인자인 PPARγ, C/EBPα, SREBP1의 발현을 억제시켜 지방 합성이 감소됨을 확인하였다. 또한, 마우스 지방에서 분리한 줄기세포의 지방 분화과정에서도 댕댕이나무 열매 추출물이 PPARγ, C/EBPα, SREBP1의 단백질 발현을 감소시켜 지방 축적을 농도 의존적으로 억제하였다.
이상의 결과로 댕댕이나무 열매 추출물은 세포독성이 없는 농도에서 지방 세포의 분화를 억제 하는 것으로 확인되어 항비만 기능성 소재로서의 활용 가능성이 있을 것으로 사료된다.




 XML Download
XML Download