

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Thyroid-associated ophthalmopathy (TAO) is a significant extrathyroidal manifestation and comorbidity, predominantly associated with Graves’ disease (GD) and, to a lesser extent, found in euthyroid or hypothyroid patients who have thyroid autoantibodies. Approximately half of the patients with GD exhibit symptoms and signs indicative of TAO, including abnormal ocular sensations, increased lacrimation, eyelid edema or retraction, and conjunctival injection [1]. Imaging techniques such as magnetic resonance imaging or computed tomography scans detect TAO in about 70% of GD patients, encompassing both subclinical and clinical cases [1]. TAO is more prevalent among women than men and is more common in the White population than in the Asian population [2]. While mild cases of TAO typically do not require treatment beyond that for GD, moderate to severe cases, which represent 3% to 5% of TAO instances, necessitate specific targeted therapies [1]. In the most severe cases of TAO, complications such as optic neuropathy or corneal injury may occur, potentially leading to vision loss.
The underlying mechanism for TAO primarily involves autoimmune processes [1,3]. The presence of thyrotropin receptor (TSH-R) in orbital fibroblasts (OFs) and their interactions with infiltrating immune cells, including T cells, B cells, and macrophages, play a crucial role in the development and progression of TAO. In the early phase of the disease, the main pathological features include inflammation with adipose tissue expansion due to adipogenic differentiation of OFs and extraocular muscle expansion through the accumulation of glycosaminoglycans. Tissue fibrosis may occur in the later stages of TAO. Currently, glucocorticoid treatment, which modulates inflammation and immune responses, is the primary therapy for TAO. Recent studies have implicated the insulin-like growth factor (IGF)-1 receptor, in conjunction with TSH-R, in the pathogenesis of TAO, and a monoclonal antibody targeting the IGF-1 receptor has demonstrated clinical benefits [4]. However, the molecular and cellular mechanisms of TAO remain incompletely understood, and existing treatment strategies are inadequate to halt the progression of the disease, despite the significant systemic adverse effects associated with these medications.
In addition to autoimmune processes that initiate and exacerbate TAO, tissue hypoxia has also been suggested as a potential link between smoking and TAO, according to some studies [5-7]. The development of a hypoxic microenvironment in TAO can be explained by factors such as inflammation [8] and the expansion of adipose tissue, which may induce hypoxia [9]. Given the anatomical constraints of the orbit’s bony structure, any expansion of tissue within this limited space can lead to vascular compression and reduced blood flow, as observed in TAO [10,11]. If the specific roles of the hypoxic microenvironment in the progression of TAO are identified, it could lead to the development of new management strategies that complement existing treatments, which primarily focus on immune suppression.
Hypoxia-inducible factor (HIF)-1α, a crucial molecule in hypoxic conditions, significantly influences adipogenesis and fibrosis by regulating cellular metabolism and the extracellular matrix, both of which are involved in the progression of TAO [1,2]. Therefore, we explore the impact of hypoxia on OFs from TAO patients, specifically examining the role of HIF-1α in adipogenesis during the progression of TAO.
METHODS
Cell cultures
OFs were obtained from the Department of Ophthalmology at Seoul St. Mary’s Hospital, College of Medicine, The Catholic University of Korea. TAO OFs were collected during surgeries on patients, while non-TAO OFs were derived from upper lid blepharoplasties in individuals without any history of immune, inflammatory, or thyroid diseases. The study’s protocol, including the acquisition of specimens, received approval from the Institutional Review Board of College of Medicine, The Catholic University of Korea (KC17SESI0504) and written informed consent was obtained from all patients. This research was conducted in accordance with the ethical standards of the Declaration of Helsinki for medical research involving human participants. The clinical characteristics of the primary cells are presented in Supplemental Table S1.
Primary cultures of OFs were grown in Dulbecco’s Modified Eagle’s Medium (Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (Gibco) and 1% penicillin-streptomycin (Thermo Fisher Scientific, Waltham, MA, USA).
Oil Red O stain
OFs were seeded at a density of 5×103 cells per well in 6-well plates. They were cultured in adipogenesis medium, specifically Human Adipose-derived Mesenchymal Stem Cells Complete Medium (Cyagen, Santa Clara, CA, USA). Following a 14-day incubation, the media were discarded, and the wells were washed three times with phosphate-buffered saline (PBS). Each well was then fixed with 10% formaldehyde for 10 minutes, followed by a rinse with distilled water. Subsequently, each well was stained with a 0.3% Oil Red O solution in distilled water and isopropanol for 20 minutes. After staining, the wells were washed with a 60% isopropanol solution. Finally, the wells were left to dry overnight on a clean bench and were examined under a microscope (Nikon TS-100F) at 400× magnification.
Western blot analysis
OFs were washed three times with PBS and lysed using radioimmunoprecipitation assay (RIPA) lysis buffer (Bio-Rad, Hercules, CA, USA) supplemented with a protease inhibitor cocktail (Bio-Rad). Protein samples, each containing 50 μg, were prepared using the bicinchoninic acid method with a bovine serum albumin standard and subsequently loaded onto 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels. After electrophoresis, the proteins were transferred to membranes (Millipore, Bedford, MA, USA), and Ponceau S staining (Abcam, Cambridge, UK) was employed to assess the efficiency of the transfer. The membranes were then blocked overnight in a solution of 5% skim milk and 5% bovine serum albumin in PBS containing 0.2% Tween 20 (Bio-Rad). This was followed by incubation with the primary antibody. After the primary antibody incubation, the membranes were probed with a horseradish peroxidase-conjugated secondary antibody and developed using SuperSignal West Pico PLUS Chemiluminescent Substrate (Thermo Fisher). Protein detection was performed with the ChemiDoc MP system (Bio-Rad).
IDF-11774 preparation and the IC50 cell viability assay
IDF-11774 (provided by Ildong Pharmaceutical Co. Ltd., Seoul, Korea), previously known as Lw6, is an HIF-1α inhibitor that has demonstrated effectiveness in treating colorectal, lung, and thyroid cancers [12-14]. Cell viability was evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma, St. Louis, MO, USA) assay. Cells were treated with varying concentrations of IDF-11774 (0, 0.5, 1.0, 1.5, 2.0, and 2.5 μM) and incubated at 37°C in a 5% CO2 atmosphere for 48 hours. Following this, 10 μL of MTT solution was added to each well. After incubation for 4 hours in a 5% CO2 environment at 37˚C, the medium was removed from all wells. Isopropyl alcohol was added to each well, followed by a 20-minute incubation period, after which distilled water was added. The absorbance at 570 nm was measured using a Tecan Infinite M200 Pro microplate reader (Tecan Life Sciences, Männedorf, Switzerland). The half-maximal inhibitory concentration (IC50) was determined as the concentration that reduced the absorbance in normal cells by 50% in the MTT assay, indicating a 50% reduction in cell viability.
Hypoxia chamber
A hypoxia chamber was utilized to create hypoxic conditions, under which the cultured dishes were handled and treated. The cells were incubated at 37°C in an atmosphere containing 5% CO2. Hypoxic conditions were established using a gas mixture of 1% O2, 5% CO2, and 94% N2 at a constant temperature of 37°C over a 48-hour period in an incubator chamber (Baker Ruskinn, Bridgend, UK). OFs were not used beyond passage number 8 in the initial culture.
Statistical analyses
We categorized experimental conditions into six groups to distinguish between normoxic and hypoxic environments. Further subdivisions were based on IDF-11774 treatment, allowing for a detailed analysis of the effects on adipogenesis. To enable focused analyses, we created data subsets based on specific samples aligned with each sample group and condition. To visually depict the distribution of protein expression values across various conditions, we employed boxplots demonstrating the median, quartiles, and outliers. For comparing protein expression levels in normoxic versus hypoxic conditions across each sample group, we utilized the Wilcoxon signed-rank test. A nonparametric test for paired data was applied to assess proteins like HIF, CCAAT/enhancer binding protein (CEBP), and peroxisome proliferator-activated receptor γ (PPARγ) in each sample. Protein expression values were represented using the median and interquartile range. We performed two-way analysis of variance (ANOVA) to evaluate the effects of group (TAO vs. non-TAO), condition (normoxic vs. hypoxic), and their interaction on protein expression values. To strengthen the robustness of our findings, we used bootstrapping analysis to validate the F values and P values obtained from two-way ANOVA. Additionally, we conducted one-way ANOVA to evaluate the variation in protein expression and cytokines across different treatment conditions. After ANOVA, the Tukey honest significant difference post hoc test was conducted to evaluate the significance of differences between groups. Statistical analyses were performed using R version 4.3.0 (R Foundation for Statistical Computing, Vienna, Austria), and statistical significance was defined as a P value less than 0.05.
RESULTS
Expression of HIF-1 α in TAO and non-TAO OFs
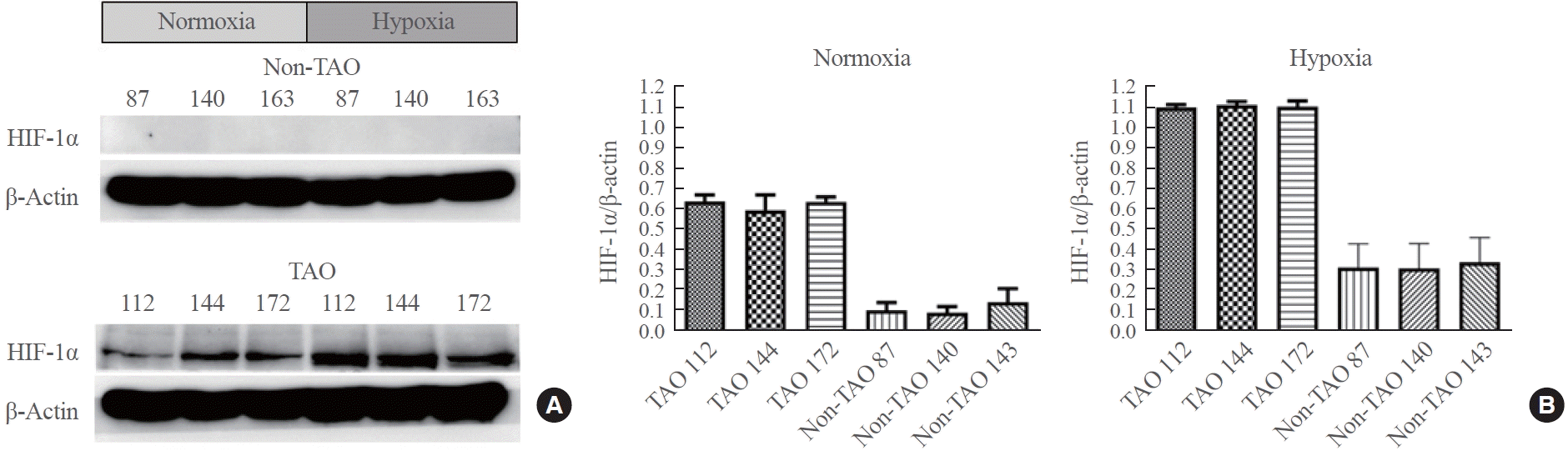
To investigate HIF-1α expression in OFs, primary cells from TAO patients and non-TAO controls were cultured under normoxic (5% CO2 in air) and hypoxic conditions. Fig. 1 illustrates the HIF-1α expression levels in each sample under both normoxia and hypoxia. Two-way ANOVA revealed significant main effects for both group and condition, as well as a significant interaction effect. TAO OFs exhibited significantly higher HIF-1α expression than non-TAO OFs (F(1, 32)=332.012, P<0.001, η2=0.735). Additionally, hypoxia led to a significant increase in HIF-1α expression compared to normoxia (F(1, 32)=93.189, P<0.001, η2=0.206). The interaction effect indicated that the increase in HIF-1α expression under hypoxia was more pronounced in TAO OFs than in non-TAO OFs (F(1, 32)=14.779, P<0.001, η2=0.033). Bootstrapped F values confirmed the robustness of these findings, with mean values of 403.0745 for the group effect, 128.1350 for the condition effect, and 16.92281 for the interaction effect, further supporting the observation of significant differences. These results suggest that HIF-1α expression is significantly higher in TAO OFs, especially under hypoxic conditions.
Adipogenic phenotype changes under normoxia and hypoxia
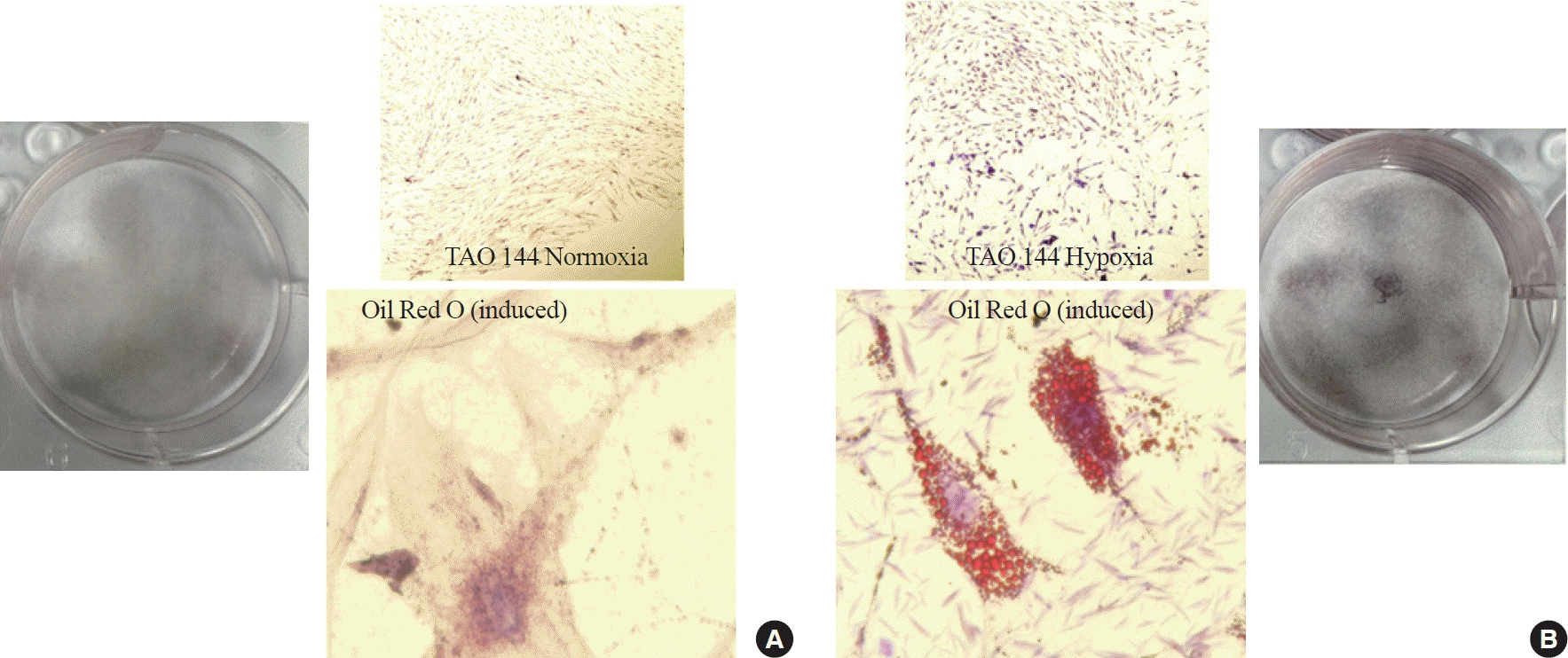
Phenotypically, TAO OFs cultured in adipogenic medium for 14 days exhibited adipogenic differentiation (Fig. 2). Under hypoxic conditions, more prominent lipid droplets were observed compared to those under normoxic conditions.
Expression of adipogenic markers: effects of hypoxia and HIF-1α inhibitor treatment
Baseline expression under hypoxia
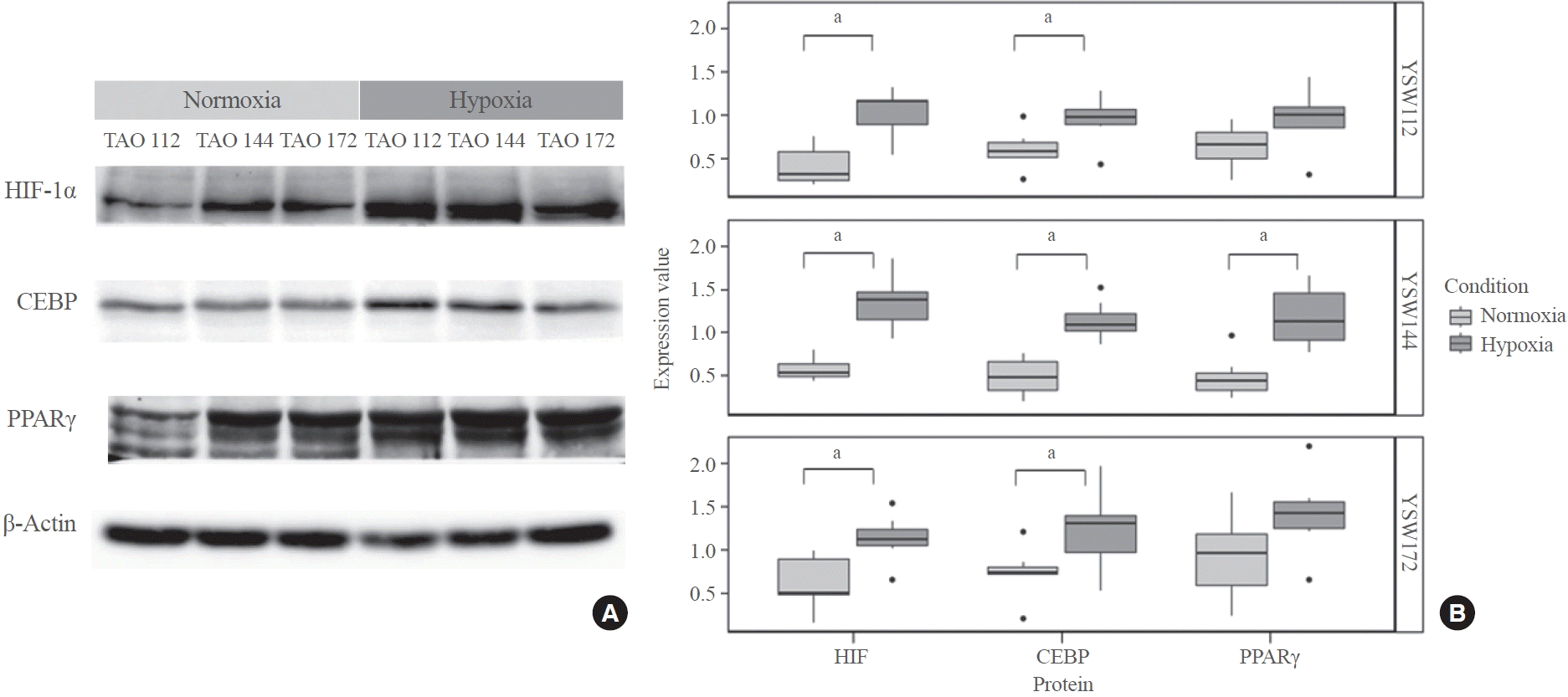
As illustrated in Fig. 3, all TAO samples exhibited higher HIF1α expression under hypoxic conditions. Additionally, CEBP and PPARγ expression levels were elevated in hypoxic conditions compared to normoxic conditions. Notably, TAO 144 was the sample that demonstrated the most significant increase in protein expression in response to hypoxic stimuli, with statistical significance.
Impact of HIF-1α inhibitor treatment
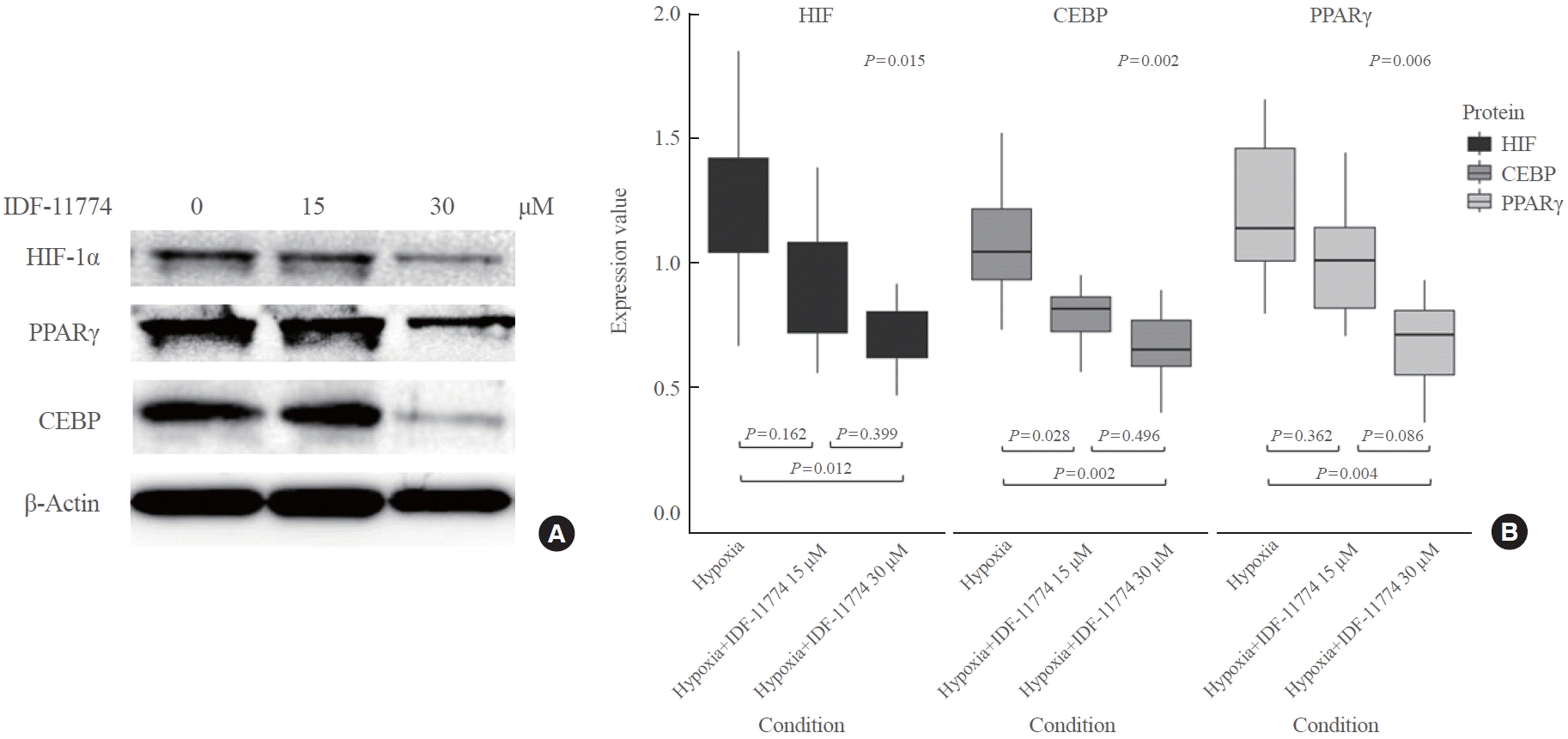
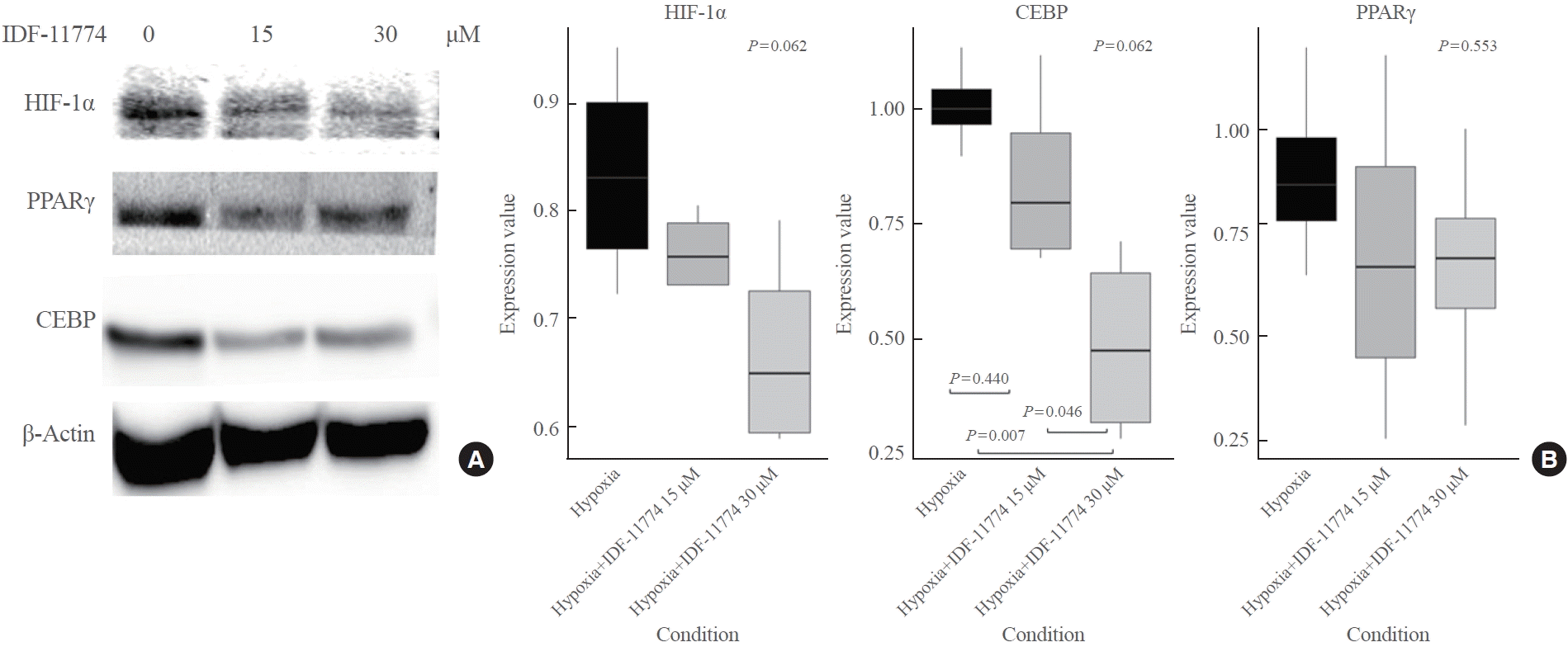
Next, we investigated whether the expression of HIF-1α, CEBP, and PPARγ could be affected by inhibiting HIF-1α using IDF-11774 (IC50=58.6 and 51.5 μM in normoxia and hypoxia, respectively) (Supplemental Fig. S1). We observed a decrease in the expression of HIF-1α, CEBP, and PPARγ with increasing concentrations of IDF-11774 (15 and 30 μM) as shown in Fig. 4. There was a statistically significant difference in the expression levels of all proteins among the no-treatment group, the IDF-11774 15 μM group, and the IDF-11774 30 μM group. In post hoc analysis, a significant decrease in the expression of HIF-1α, CEBP, and PPARγ was noted between the no-treatment group and the IDF-11774 30 μM group (P=0.011, P=0.002, and P=0.004, respectively).
Effects of IGF-1 stimulation and HIF-1α inhibition
To further investigate whether the HIF-1α inhibitor elicited a similar response under IGF-1 stimulation as it did under hypoxia alone, TAO OFs were exposed to IGF-1 (20 mM) under hypoxic conditions, followed by treatment with the HIF-1α inhibitor. Stimulation with IGF-1 resulted in increased expression of HIF-1α, CEBP, and PPARγ. However, the presence of the HIF1α inhibitor significantly reduced the expression of these markers, even in the presence of IGF-1 (Fig. 5). Specifically, the expression of HIF-1α showed a borderline significant decrease (P=0.06), CEBP expression demonstrated a significant reduction (P=0.007), and PPARγ expression did not exhibit a significant change (P=0.5530).
Effects of hypoxia and HIF-1α inhibition on inflammatory cytokines
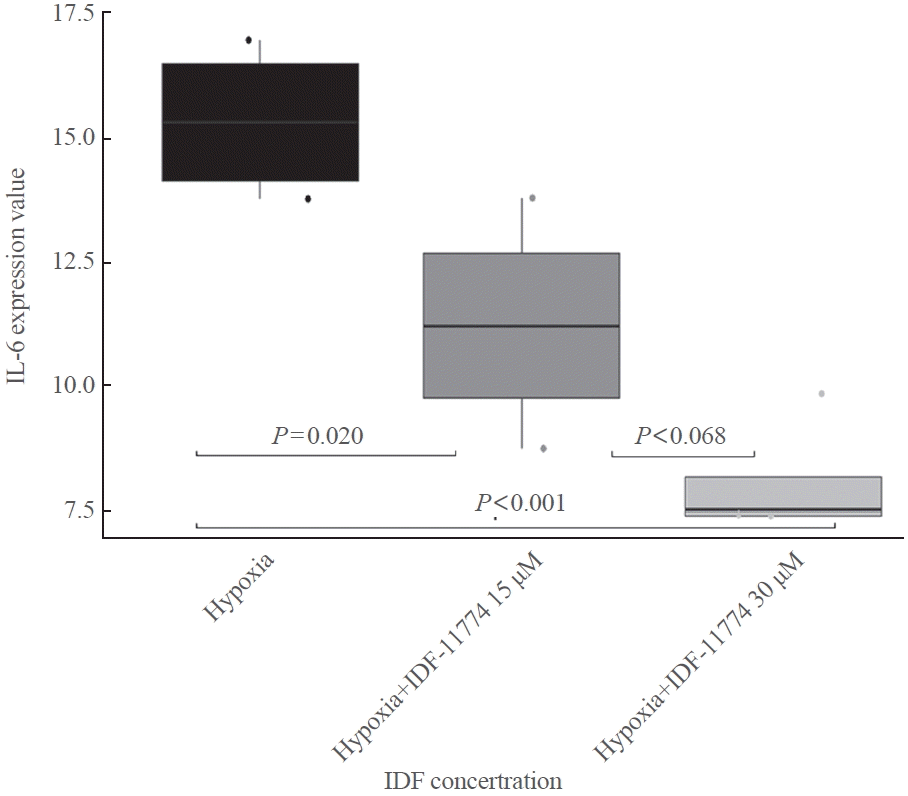
We analyzed the effects of hypoxia and HIF-1α inhibition on inflammatory cytokines, including tumor necrosis factor α, interleukin 1β (IL-1β), IL-2, and IL-6. Hypoxia significantly increased IL-6 levels (Supplemental Fig. S2), and this elevation was substantially reduced by the HIF-1α inhibitor (Supplemental Fig. S3). Additionally, the IL-6 response to the HIF-1α inhibitor was evident when IGF-1 stimulation was introduced under hypoxic conditions (Fig. 6). ANOVA revealed a significant effect of IDF-11774 concentration on IL-6 expression levels (P<0.001). In the post hoc analysis, IL-6 expression at an IDF-11774 concentration of 15 μM was significantly lower than that of the control (P=0.020). Similarly, IL-6 expression at an IDF-11774 concentration of 30 μM was significantly lower than that of the control (P<0.001). However, there was no significant difference in IL-6 expression between IDF-11774 concentrations of 30 μM and 15 μM (P=0.068).
DISCUSSION
In this study, we observed prominent HIF-1α expression and a robust increase in response to hypoxic stimuli in TAO OFs. Under hypoxic conditions, TAO OFs exhibited changes to an adipogenic phenotype. Additionally, the expressions of adipogenic differentiation markers in TAO OFs, specifically CEBP and PPARγ, were more pronounced under hypoxia than under normoxic conditions. Most importantly, inhibition of HIF-1α significantly downregulated adipogenic markers (CEBP and PPARγ) and the inflammatory cytokine IL-6, highlighting its potential role in modulating adipogenesis and inflammation in TAO OFs.
HIF-1α is a main regulator in response to hypoxic stimuli under both physiological and pathological conditions. In normoxia, HIF-1α is located in the cytoplasm and is rapidly degraded by the proteasome, resulting in undetectable levels of HIF-1α [15]. In contrast, under hypoxic conditions, HIF-1α stabilizes and translocates from the cytoplasm to the nucleus. It then dimerizes with HIF-1β, initiating gene transcription that enables cells to adapt and survive under hypoxic conditions [16]. HIF1α activates the transcription of genes involved in various cellular processes, including angiogenesis (vascular endothelial growth factor [VEGF]) [17,18], epithelial fibrosis (plasminogen activator inhibitor-1) [19], glucose metabolism (oxygen-dependent tricarboxylic acid cycle and glucose transporters) [20], iron metabolism and erythropoiesis (transferrin and transferrin receptor) [21], cell proliferation (IGF and transforming growth factor-α) [22], and apoptosis (p53 and p21) [23]. Consequently, tissues must adapt to changes in oxygen concentration, and various targets of HIF-1α are regulated in a tissue-specific manner [16]. Although HIF-1α is typically rapidly degraded under normoxic conditions, it can still be detected in certain situations. Basal cellular activities may result in low levels of HIF-1α even in the presence of normal oxygen levels. Moreover, specific cell types, such as cancer cells [24,25], or external conditions, such as inflammation [26,27], can stabilize or accumulate HIF-1α despite adequate oxygen availability.
The complex interplay between the immune system and OFs is known to play a crucial role in the initiation and progression of TAO [1]. Recent studies have also implicated the IGF-1 pathway in the pathogenesis of TAO [28]. Moreover, based on clinical and laboratory evidence, hypoxia is likely to contribute to the progression of TAO. Firstly, smoking, which can induce tissue hypoxia or create a hypoxic microenvironment [6], is a well-known risk factor for both the development and exacerbation of TAO. Secondly, the expansion of tissue within the confined bony orbit in TAO could lead to a hypoxic microenvironment. Lastly, hypoxia has been shown to play a significant role in the adipogenic differentiation of fibroblasts observed in OFs of TAO patients [6,7,29]. In light of these findings, we hypothesize that HIF-1α, a primary regulator in response to hypoxic conditions, plays a significant role and could be an effective target for treatment.
To date, limited data demonstrate the role of hypoxia in the progression of TAO. Metcalf and Weetman [5] showed that OF responded to cytokine (IL-1 and interferon gamma) stimulation under hypoxic conditions. However, their experiments were conducted only on OFs from healthy subjects. Another study identified differences in adipogenesis under hypoxic conditions between TAO OFs and controls [6]. Gortz et al. [7] investigated the HIF-1-dependent VEGF and adipogenic differentiation in TAO OFs, noting increases associated with smoking. Recent research has furthered this understanding by demonstrating that the interaction between macrophages and OFs under hypoxic conditions not only perpetuates inflammation but also significantly enhances adipogenic differentiation, suggesting a compounding effect on the pathogenesis of TAO that could inform new therapeutic strategies [29]. Van Regemorter et al. [30] found that the expression of HIF-1α protein was increased in TAO adipose tissues but not in TAO OFs. We also observed that HIF-1α expression levels in TAO OFs were higher under normoxia than those in non-TAO OFs and significantly increased under hypoxia only in TAO OFs. In our study, adipogenesis, measured by Oil Red O staining, was increased in TAO OFs under hypoxia. Adipogenesis is associated with the activation of several adipocyte-specific genes such as adiponectin and leptin [31]. PPARγ and CEBPβ stimulate adiponectin and act as factors for terminating the differentiation of fibroblasts into adipocytes [32]. A previous study reported that PPARγ is expressed in the orbital adipose tissue of five TAO patients and is stimulated in cultures of TAO with treatment of a PPARγ agonist (thiazolidinediones) [33]. Our study also confirmed a consistent result of increased PPARγ and CEBP levels under hypoxia in TAO OFs, while those adipogenic marker expressions decreased after a dose-dependent IDF-11774 treatment. Additionally, among inflammatory cytokines, IL-6 responded to hypoxic stimuli and HIF-1α inhibition. However, while the effect of HIF-1α inhibition on IL-6 was maintained with IGF-1 stimulation under hypoxia, the effect on HIF-1α expression was attenuated and that on PPARγ expression was not significant. Therefore, HIF-1α inhibition might not be a good candidate for single therapy of TAO.
Our findings have several clinical implications. First, in conjunction with prior studies [5-7,29], HIF-1α, which is induced by microenvironmental changes such as hypoxia, could be a new target for preventing the progression of TAO. Second, the microenvironment, which is known to play a significant role in cancer progression, could also be crucial in benign diseases such as TAO [20]. Therefore, manipulating aspects of the microenvironment beyond hypoxia might represent another strategic approach for managing benign diseases. Lastly, since the HIF-1α-related mechanism in TAO progression operates independently from immune processes, combining these treatments could be a viable option. This approach could enhance treatment effectiveness, as demonstrated in rheumatoid arthritis [34]. By introducing a new treatment target apart from the immune system, it may be possible to reduce steroid doses and thereby decrease the risk of complications. Furthermore, a strength of this study lies in the fact that all laboratory procedures under hypoxic conditions were conducted within a hypoxic chamber. In contrast, previous studies did not consistently perform all procedures within such a chamber, potentially exposing samples to oxygen.
However, there are several limitations of our study. As this study utilized primary cells from several TAO patients, further studies using various types of TAO OFs are required to generalize the findings. Additionally, it is important to note that primary cells can undergo phenotypic changes at higher passage numbers. Therefore, in our experiments, OFs were not used beyond passage number 8 in the initial culture. This precaution should be considered in future TAO studies to maintain consistent cellular characteristics. The proposed mechanism suggests that HIF-1α appears to be involved in the progression of TAO, but not in its initial development.
In conclusion, our study has elucidated the pivotal role of HIF-1α in the progression of TAO, especially under hypoxic conditions. Importantly, the application of a HIF-1α inhibitor effectively decreased the expression of adipogenic markers, underscoring the potential of targeting HIF-1α as a therapeutic strategy in TAO management.
 XML Download
XML Download