

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Neurodegenerative diseases such as Alzheimer’s disease (AD) and Parkinson’s disease are frequently observed in older populations. AD is a complex disease characterized by cognitive dysfunction in learning and memory [1]. Pathological characteristics of AD include astrogliosis and microglial activation, which are associated with inflammatory responses and cholinergic dysfunction. These pathological processes lead to neurodegeneration and synaptic and neuronal loss [2].
Inflammation is a harmful stimulus that causes tissue damage, including the release of inflammatory mediators, such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) [3]. Neuroinflammation has been observed in AD brains due to the release of pro-inflammatory factors [4]. Astrocytes in the central nervous system form an integral part of the blood-brain barrier (BBB) [5]. Astrocytes interact with other cells including neurons and play a role in maintaining tissue homeostasis, supporting neuronal development, and inflammatory modulation [56]. Glial fibrillary acidic protein (GFAP) is an astrocyte marker [7]. High level of GFAP release was related to worse memory in AD animal studies [8]. Thus, the inhibition of the inflammatory response may protect against memory loss.
Acetylcholinesterase (AChE) inhibitors have been developed based on the cholinergic hypothesis, which explains the pathogenesis of AD associated with cholinergic activities [910]. In patients with AD, cholinergic neurons are lost with a reduction in M2 muscarinic receptors, which regulate the release of acetylcholine [11]. Enhancement of cholinergic signaling in the central nervous system improves memory function [9]. Moreover, the regulation of acetylcholine by AChE can attenuate inflammation [12]. Thus, controlling cholinergic activity may prevent AD.
Scopolamine (Sco) is a muscarinic acetylcholine receptor antagonist that reduces neurotransmission in the central nervous system and induces cognitive dysfunction including memory loss [1314]. Sco can readily permeate the BBB and induce cognitive disorders. In addition, Sco can mimic AD conditions and has been considered a potent model of AD [915]. Thus, Sco has been widely used as an experimental model of cognitive impairment in neuroscience-related studies [16].
Herbal medicines are marked by multiple components and effects, and play important roles in the development of new therapies with several advantages over conventional chemical compound-based medications [17]. Both safflower (Carthamus tinctorius L.) and dandelion (Taraxacum coreanum, TC) belong to the Asteraceae family and have been widely used as folk medicines, exerting strong anti-inflammatory, antioxidant, hepatoprotective, and antimicrobial activities [18]. Seeds of safflower (Carthamus tinctorius L. seeds, CTS) are used to treat osteoporosis and rheumatism in Korea [19], and TC is a plant native to Korea with antioxidative and anticancer activities [2021]. In addition, it has been reported that the pharmacological activities of these herbs are attributed to their various phytochemical components, such as phenolic acids, polyphenolic compounds, polysaccharides, and lignans [18]. According to the previous studies, herbal medicines are often used in compatibility rather than being used in single, to promote effects and to reduce adverse reactions [22]. The most common and highest frequency of compatibility among herbal pairs is 1:1 [2223]. A proportion test design method, called the increase-decrease baseline geometric proportion design method, is commonly used for studying the ratio between 2 herbal medicines to explore whether the combination has a synergistic effect or to screen the best ratio for the combination [242526]. According to this method, when one herb is combined with another, herb A increases by 10–30% while herb B decreases by 10–30% until the maximum compatibility of these 2 herbs is reached, which means herb A alone and vice versa [24]. Therefore, in this study, the ratio of the CTS–TC combination was set to 1:4, 1:1, and 4:1, and the limit of the ratio was CTS and TC alone, to investigate whether these ratios have protective effects against memory deficits induced by Sco. Additionally, we determined the potential active components of the CTS–TC combination using high-performance liquid chromatography (HPLC). Previous studies have identified various phytochemicals in safflower and TC and have indicated the pharmacological activities of each plant. For example, hydroxysafflor yellow A from the flower of safflower has been found to exert anti-inflammatory, anti-cancer, vascular, brain, and liver disease effects, as well as cell proliferation and inhibition effects [27]. TC is rich in compounds such as chicoric acid, chlorogenic acid, caffeic acid, and luteolin, which show hepatoprotective, anti-inflammatory, antiviral, and antidepressive effects [2829]. Recent studies have demonstrated that serotonin derivatives from CTS may exert antioxidant and cholinergic improvement effects [30]. Therefore, we chose serotonin and its derivatives (N-feruloylserotonin and N-(p-coumaroyl) serotonin) from CTS and the abundant compounds from TC (chicoric acid and chlorogenic acid) for HPLC analysis to explore the contents of these compounds in combination with CTS and TC.
Go to : 
MATERIALS AND METHODS
Sample preparation
Water extracts of CTS, TC, and the CTS–TC combination at ratios of 4:1, 1:1, and 1:4 were obtained through a series of processes including pulverization, extraction, concentration, and drying. Briefly, CTS and the aerial part of TC were dried, pulverized, and then mixed according to the design proportions (CTS:TC at 4:1, 1:1, and 1:4). Subsequently, CTS, TC, and their mixtures were placed in an extraction tank, and purified water was added 20 times. Heat extraction was conducted at 90°C for 8 h and then filtered using a 200-mesh filter. The extracts were concentrated under a temperature at 60°C and then freeze-drying for 48 h. The water extracts of CTS and TC yielded 8.4% and 30.6%, respectively, of the original herbs, and the water extracts of the CTS–TC combination at 4:1, 1:1, and 1:4 ratios yielded 16.0%, 26.3%, and 34.0%, respectively, of the original mixtures.
HPLC analysis
An aliquot of the aqueous extract (10 mg) of CTS–TC combination (at the ratio of 4:1, 1:1, and 1:4, respectively) was dissolved in 10 mL of 50% methanol and passed through a Dismic-25 JP membrane filter (Advantec Toyo, Tokyo, Japan; pore diameter: 5 μm). Next, 10 μL of the sample was injected into a reverse-phase HPLC system with a YMC-Pack ODS-AM (4.6 × 250 mm, 5 μm pore size, YMC, Kyoto, Japan) column at 35°C. The mobile phase consisted of a 0.1% aqueous solution of formic acid and acetonitrile and the following gradient conditions were used: 0 min, 0% B; 4 min, 0% B; 10 min, 6% B; 14 min, 22% B; 60 min, 28% B; and 70 min, 100% B. The flow rate was 0.8 mL/min. UV absorbance at 280 nm was monitored using an Agilent 1260 series diode array detector (Agilent Technologies, Waldbronn, Germany). All peaks were assigned through co-injection tests with authentic samples and by comparison with the UV spectral data. All measurements were performed in triplicate.
Animal experiment
Five-week-old male ICR mice (Orient Inc., Seongnam, Korea) that weighed 26–28 g were housed in an environment with a 12 h light/dark cycle, a constant temperature of 22 ± 2°C, and humidity of 50 ± 10%. The animal experimental procedures followed the Animal Research: Reporting of In Vivo Experiments guidelines 2.0, and the experimental protocol was approved by the Pusan National University Institutional Animal Care and Use Committee (approval number: PNU-2018-1873). The mice were fed a standard laboratory diet and had free access to water.
The experimental schedule is illustrated in Fig. 1. After one week of acclimatization, the mice were randomly divided into 7 groups of similar average weights. The experiment groups are as follows: 1) Normal (Nor) = water + 0.9% NaCl; 2) Sco = water + Sco; 3) CTS = CTS + Sco; 4) TC = TC + Sco; 5) 4:1 = CTS–TC combination at a ratio of 4:1 + Sco; 6) 1:1 = CTS–TC combination at a ratio of 1:1 + Sco; 7) 1:4 = CTS–TC combination at a ratio of 1:4 + Sco. Samples (100 mg/kg, 100 µL/day) dissolved in water were orally administered to mice by using an oral gavage needle for 14 consecutive days, while those in the Sco group received water on the same schedule. To induce memory deficits, mice (Sco, CTS, TC, and CTS–TC combination groups) were received 1 mg/kg of Sco injection (intraperitoneal injection, 100 µL/day) 30 min before each behavioral test, while the Nor group received 0.9% NaCl solution [31]. The animals were sacrificed on the day of behavioral tests. The whole brains of the mice were quickly removed and stored at −80°C.
 | Fig. 1Time schedule of the animal experiment. Mice were acclimatized for 1 week. Samples including CTS, TC, and CTS–TC combination (100 mg/kg/day) were orally administered to the mice for 14 days. Scopolamine (1 mg/kg) was intraperitoneal injected to the mice 30 min before each behavioral test.CTS, Carthamus tinctorius L. seeds; TC, Taraxacum coreanum.
|
T-maze test
To investigate spatial working memory, the T-maze test was conducted using the method described by Montgomery [32], with a slight modification. The black-painted T-shaped equipment contained 3 regions: start arm, left arm, and right arm (70 cm total width of the maze, 65 cm total length, and 7 cm height). A clapboard separated the left and right arms. Behavioral experiments were conducted over 2 days. On the first day, during the training session, each mouse explored from the start arm to the left arm for 4 min while the right arm was blocked. After the training, the mice rested in their cages for 24 h. The next day, during the test session, the arms were opened, and the mice were allowed to explore both arms for 4 min. The frequency of exploring each arm was recorded, and the space perceptive ability (%) was calculated as the ratio of the frequency of exploring the left or right arm to the total number of explorations, multiplied by 100.
Novel object recognition test
The recognition memory of mice was determined using the novel object recognition test [33]. The test was performed using a black-painted open-field square apparatus (40 cm × 40 cm × 40 cm). In the training session, 2 identical objects (A and A) were placed at a fixed distance in the apparatus, and the mice were allowed to move and touch each object for 4 min. The test session started after 24 h and allowed the mice to explore the objects for 4 min; however, one of the objects used in the training session was replaced with a completely different object (A and B). Cognitive function in mice was measured as object cognitive ability (%), which describes the ratio of the number of touches of the familiar object (training session) to the novel object (test session) over the entire exploration time.
Western blot analysis
To observe protein expression changes, mouse brain tissue was homogenized in lysis buffer (radioimmunoprecipitation assay buffer mixed with protease inhibitor cocktail) and the tissue homogenate was centrifuged at 12,000 rpm at 4°C for 20 min. Protein concentration was determined using the Bradford Protein Assay Kit (Bio-Rad, Hercules, CA, USA) in accordance with the manufacturer’s instructions. Several dilutions of protein standard range 0–500 µg/mL was prepared using a known concentration of bovine serum albumin (1 mg/mL), to generate a standard curve. The 200 µL of Bradford working reagent (Bio-Rad) was added to standards and samples, mixed and then recorded the absorbances at 595 nm. The amount of protein in each sample was determined using a standard curve. Equal amounts (15 µg/lane) of samples were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis. Proteins were then transferred onto polyvinylidene fluoride membranes. A skim milk solution (5%) was used to block nonspecific binding sites on the membranes. Then the membranes were incubated with primary antibodies including AChE (sc-373901; Santa Cruz Biotechnology, Santa Cruz, CA, USA), iNOS (#13120; Cell Signaling, Danvers, MA, USA), COX-2 (#12282; Cell Signaling), GFAP (#12389; Cell Signaling), and β-actin (#8457; Cell Signaling) overnight at 4°C. The membranes were washed and incubated with secondary antibodies (anti-rabbit and anti-mouse, #7074 and #7076, respectively; Cell Signaling Technology). An enhanced chemiluminescence solution mix was used to visualize the bands using a chemiluminescent imaging system (Davinch Chemi, Seoul, Korea).
Statistical analysis
Data were presented as the mean ± SD. Statistical analyses were conducted using SPSS 23.0 (SPSS Inc., Chicago, IL, USA). For comparisons among groups, data were analyzed using a 1-way analysis of variance, followed by Duncan’s post-hoc test. The comparison between the old route (familiar object) and the new route (novel object) was conducted using the Student’s t-test. Statistical significance was set at P < 0.05 or less.
Go to :
RESULTS
Effect of CTS–TC combination on spatial working memory in Sco-injected mice
As shown in Fig. 2, no significant difference was observed between mice approaching the old and new routes in the Sco group, indicating that a cognitive deficit was induced by Sco. In contrast, significant differences were observed between the old and new routes in the groups of TC (46.2% vs. 53.8%, P = 0.008), 1:1 (41% vs. 59%, P = 0.001), and 1:4 (43% vs. 57%, P < 0.001). In the new route, ratios of 1:1 and 1:4 showed more frequent entries after training compared to CTS or TC treatment alone.
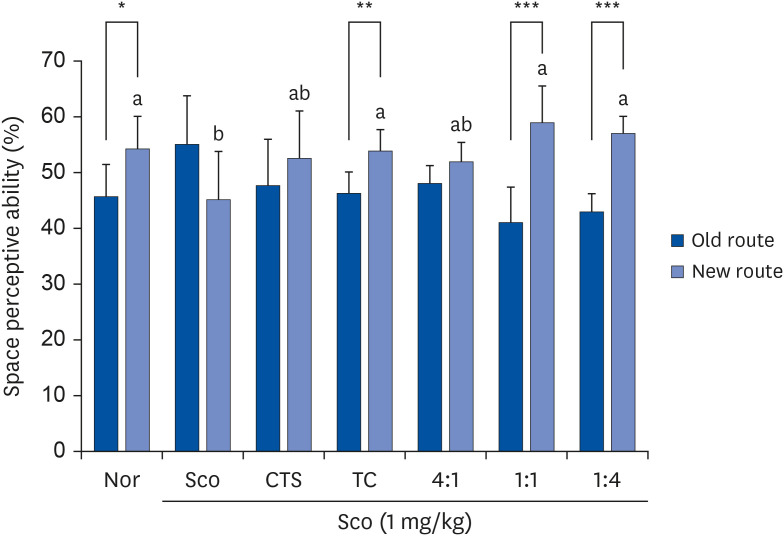 | Fig. 2Effects of CTS–TC combination on T-maze test in Sco-induced mice. Mice were allowed to explore one arm (one arm was blocked) for 4 min and then returned to their home cage. One day after training, 2 arms were opened and allowed the mice to explore for 4 min. The times for the mice to enter each arm were recorded.Values are expressed as mean ± SD (n = 6/group).
The experiment groups are as follows: Nor = 0.9% NaCl + water; Sco = Sco + water; CTS = CTS + Sco; TC = TC + Sco; 4:1 = CTS–TC combination at 4:1 ratio + Sco; 1:1 = CTS–TC combination at 1:1 ratio + Sco; 1:4 = CTS–TC combination at 1:4 ratio + Sco.
Nor, normal; Sco, scopolamine; CTS, Carthamus tinctorius L. seeds; TC, Taraxacum coreanum.
*P < 0.05, **P < 0.01, ***P < 0.001 are significantly different between old and new routs by Student’s t-test.
a,bSignificantly different (P < 0.05) among the groups by Duncan’s multiple range test.
|
Effect of CTS–TC combination on object recognition memory in Sco-injected mice
No significant differences were observed between the groups during the training sessions. In the test session (Fig. 3) using 2 different objects (A and B), the Nor group showed higher exploration of the novel object than the familiar object. However, Sco significantly induced cognitive deficits in the Sco group, with no significant difference between the novel and familiar objects. Significant differences between familiar and novel objects were found for CTS (45.3% vs. 54.8%, P = 0.046), TC (43.7% vs. 56.3%, P = 0.032), 1:1 (41.5% vs. 58.5%, P < 0.001), and 1:4 (39.2% vs. 60.8%, P < 0.001).
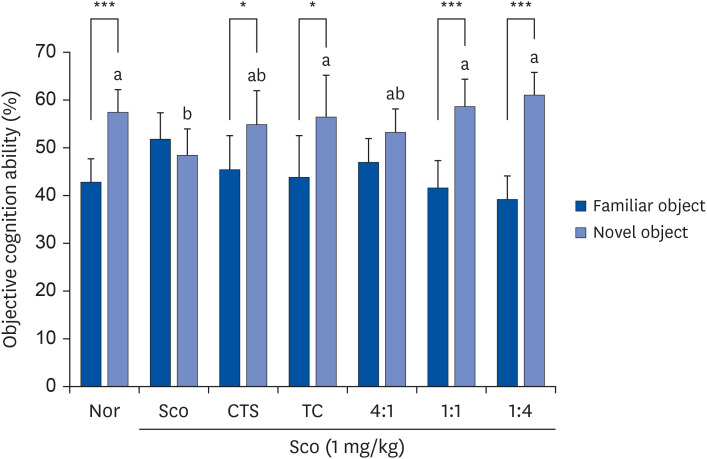 | Fig. 3Effects of CTS–TC combination on novel object recognition test in Sco-induced mice. Mice were allowed to explore the same objects (A and A’) for 4 min and then returned to their home cage. One day after training, one of the objects was replaced (A and B) and allowed the mice to explore for 4 min. The times for the mice to contact each object were recorded.Values are expressed as mean ± SD (n = 6/group).
The experiment groups are as follows: Nor = 0.9% NaCl + water; Sco = Sco + water; CTS = CTS + Sco; TC = TC + Sco; 4:1 = CTS–TC combination at 4:1 ratio + Sco; 1:1 = CTS–TC combination at 1:1 ratio + Sco; 1:4 = CTS–TC combination at 1:4 ratio + Sco.
Nor, normal; Sco, scopolamine; CTS, Carthamus tinctorius L. seeds; TC, Taraxacum coreanum.
*P < 0.05, ***P < 0.001 are significantly different between familiar and novel objects by Student’s t-test.
a,bSignificantly different (P < 0.05) among the groups by Duncan’s multiple range test.
|
Effect of CTS–TC combination on AChE protein expression in Sco-injected mice
As shown in Fig. 4, the highest AChE protein expression was observed in the Sco group after Sco injection. However, the administration of CTS or TC alone and the CTS–TC combination significantly downregulated the expression of AChE. Interestingly, the CTS–TC combination at all ratios downregulated AChE protein expression more effectively than CTS or TC alone, especially at a 1:1 ratio, and decreased AChE protein expression by approximately 2-fold compared to CTS or TC alone.
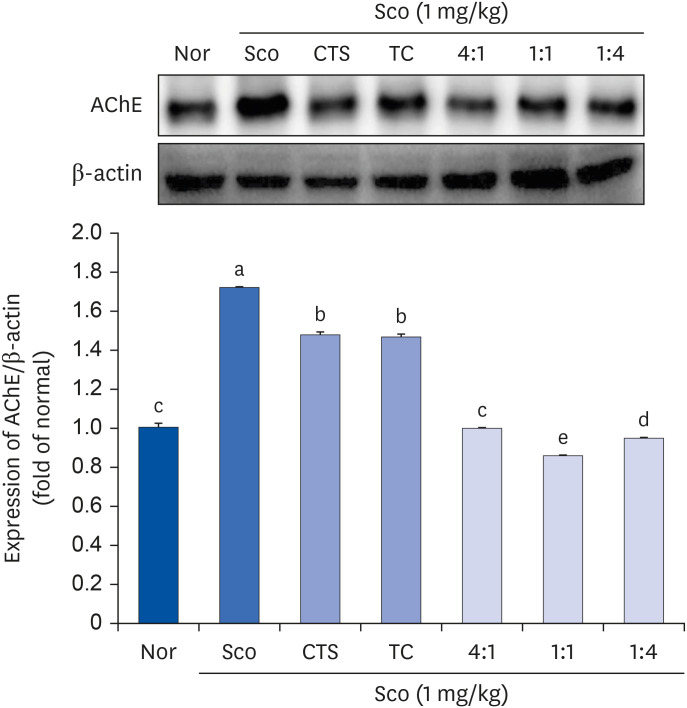 | Fig. 4Effect of CTS–TC combination on AChE protein expression in Sco-induced mice. The whole brain lysate was electrophoresed and immunoblotted with AChE antibody. The protein level was quantitated using Image J and normalized with β-actin.Values are expressed as mean ± SD.
The experiment groups are as follows: Nor = 0.9% NaCl + water; Sco = Sco + water; CTS = CTS + Sco; TC = TC + Sco; 4:1 = CTS–TC combination at 4:1 ratio + Sco; 1:1 = CTS–TC combination at 1:1 ratio + Sco; 1:4 = CTS–TC combination at 1:4 ratio + Sco.
Nor, normal; Sco, scopolamine; CTS, Carthamus tinctorius L. seeds; TC, Taraxacum coreanum; AChE, acetylcholinesterase.
a-eSignificantly different (P < 0.05) among the groups by Duncan’s multiple range test.
|
Effect of the CTS–TC combination on iNOS, COX-2, and GFAP protein expressions in Sco-injected mice
The effect of the CTS–TC combination on inflammation-related factors was evaluated (Fig. 5). iNOS and COX-2 protein expression was significantly upregulated by Sco compared to the Nor group. However, the groups administered CTS or TC alone and the CTS–TC combination significantly downregulated the expression of iNOS and COX-2. The expression of GFAP was increased in the Sco group; however, it was reversed after the administration of CTS and TC, as well as the CTS–TC combination. Moreover, the CTS–TC combination at all ratios effectively downregulated the expression of these proteins compared with CTS or TC alone.
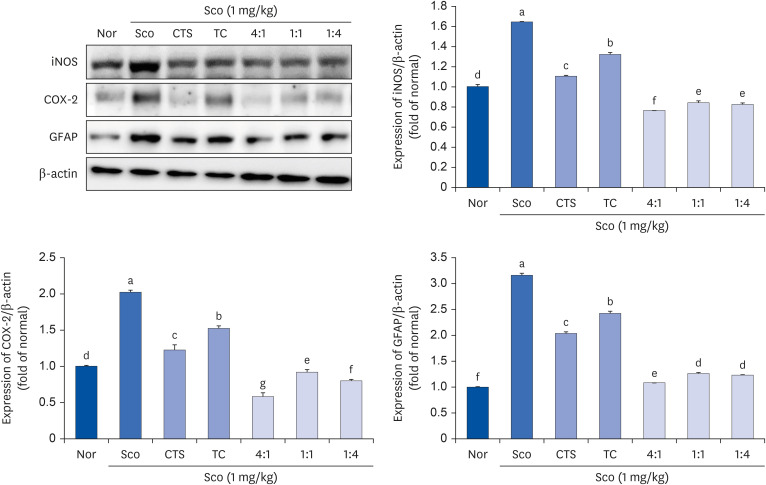 | Fig. 5Effect of CTS–TC combination on iNOS, COX-2, and GFAP protein expression in Sco-induced mice. The whole brain lysate was electrophoresed and immunoblotted with iNOS, COX-2, and GFAP antibodies. The protein level was quantitated using Image J and normalized with β-actin.Values are expressed as mean ± SD.
The experiment groups are as follows: Nor = 0.9% NaCl + water; Sco = Sco + water; CTS = CTS + Sco; TC = TC + Sco; 4:1 = CTS–TC combination at 4:1 ratio + Sco; 1:1 = CTS–TC combination at 1:1 ratio + Sco; 1:4 = CTS–TC combination at 1:4 ratio + Sco.
iNOS, inducible nitric oxide synthase; COX-2, cyclooxygenase-2; GFAP, glial fibrillary acidic protein; Nor, normal; Sco, scopolamine; CTS, Carthamus tinctorius L. seeds; TC, Taraxacum coreanum.
a-fSignificantly different (P < 0.05) among the groups by Duncan’s multiple range test.
|
Identification and quantification of the compounds of CTS–TC combination
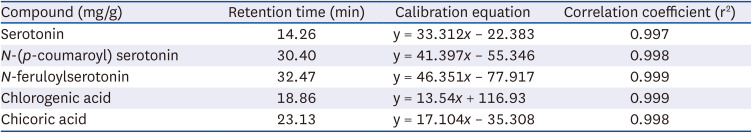
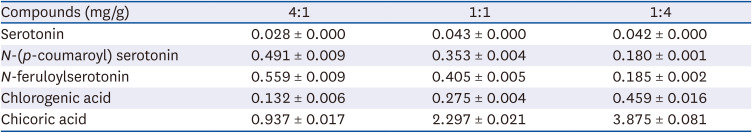
The 5 major components, chlorogenic acid, chicoric acid, serotonin, N-(p-coumaroyl) serotonin, and N-feruloylserotonin, in the CTS–TC combination were qualitatively analyzed (Fig. 6). The order of elution was as follows: serotonin > chlorogenic acid > chicoric acid > N-(p-coumaroyl) serotonin > N-feruloylserotonin. A calibration equation for each standard compound was constructed by plotting the peak area (y) against the concentration of the compound (x) (Table 1). The concentrations of these components differed depending on the ratio of CTS to TC. At 4:1 ratio, chicoric acid (0.937 ± 0.017 mg/g) was the most abundant, followed by N-feruloylserotonin, N-(p-coumaroyl)serotonin, chlorogenic acid, and serotonin (0.559 ± 0.009, 0.491 ± 0.009, 0.132 ± 0.006, and 0.028 ± 0.000 mg/g, respectively); at 1:1 ratio, chicoric acid (2.297 ± 0.021 mg/g) was the most abundant, followed by N-feruloylserotonin, N-(p-coumaroyl)serotonin, chlorogenic acid, and serotonin (0.405 ± 0.005, 0.353 ± 0.004, 0.275 ± 0.004, and 0.043 ± 0.000 mg/g, respectively); and at 1:4 ratio, chicoric acid (3.875 ± 0.081 mg/g) also the most abundant, followed by chlorogenic acid, N-feruloylserotonin, N-(p-coumaroyl)serotonin, and serotonin (0.459 ± 0.016, 0.185 ± 0.002, 0.180 ± 0.001, and 0.042 ± 0.000 mg/g, respectively) (Table 2).
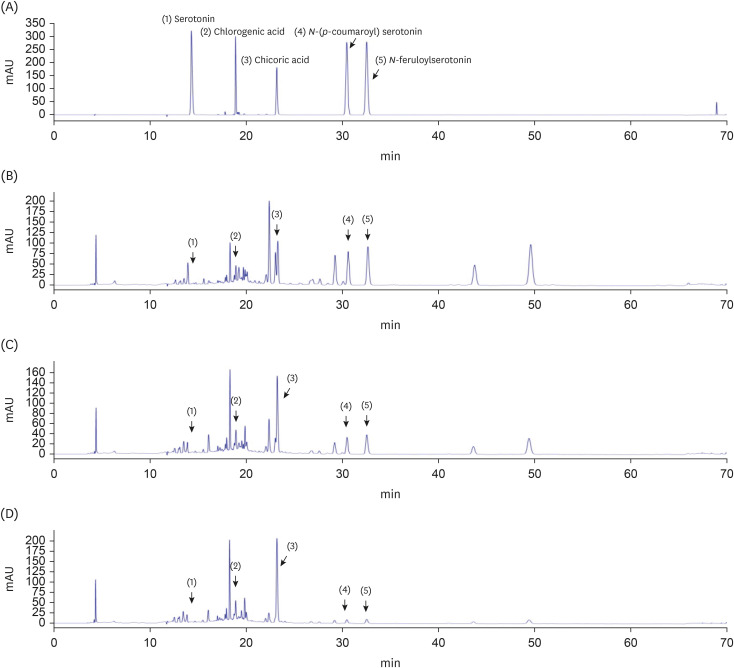 | Fig. 6High-performance liquid chromatography chromatograms of CTS–TC combination at different ratios. Serotonin (1), chlorogenic acid (2), chicoric acid (3), N-(p-coumaroyl) serotonin (4), and N-feruloylserotonin (5) in mixed standards (A), CTS–TC combination at 4:1 ratio (B), CTS–TC combination at 1:1 ratio (C), and CTS–TC combination at 1:4 ratio (D).CTS, Carthamus tinctorius L. seeds; TC, Taraxacum coreanum.
|
Table 1
Retention time, calibration equation, and correlation coefficient values
![]()
Table 2
Contents of the active compounds in CTS–TC combination
Values are presented as the mean ± SD.
The experiment groups are as follows: 4:1 = CTS: TC combination at 4:1 ratio; 1:1 = CTS:TC combination at 1:1 ratio; 1:4 = CTS:TC combination at 1:4 ratio.
CTS, Carthamus tinctorius L. seeds; TC, Taraxacum coreanum.
![]()
Go to :
DISCUSSION
While previous studies have reported the various pharmacological effects of CTS and TC extracts and have documented the effects of different parts of safflower on the nervous system, the neuroprotective effects of CTS have not yet been fully investigated. One of the most widely used forms of CTS is CTS oil, which has been shown to have positive effects on hypothalamic proliferation and neurogenesis in young adult mice [34]. A recent study by Liang and Wang [35] has reviewed the benefits of safflower, including CTS and its active constituents, for AD treatment. Kim et al. [30] previously reported that CTS contains various cholinergic-improving compounds, such as serotonin and its derivatives. CTS extract-administered mice have been shown to improve memory function by suppressing AChE expression, with an improvement in cholinergic dysfunction [30]. The combination of CTS and TC improves cognitive dysfunction [36]. Moreover, dandelion, a good candidate for food and medical use, has been reported to show neuroprotective effects indirectly due to its antioxidant and anti-inflammatory activities [3738]. Both the aerial and root parts of TC have been shown to exert neuroprotective effects in glutamate-induced mouse hippocampal HT22 cells by increasing the expression of heme oxygenase-1 and nuclear factor erythroid 2 like 2 [39]. In an AD mouse model, TC extract has been shown to prevent oxidative stress on amyloid-β (Aβ)-induced memory function by reducing lipid peroxidation level and NO production [40]. Sohn and Shin [41] have demonstrated that TC extract had an inhibitory effect on AChE activity, and significant behavioral changes were observed in passive avoidance, Y-maze, and water maze tests in a Sco-induced amnesia mouse model. In an Aβ-injected mouse model, administration of CTS and TC combination attenuated amyloidogenesis by promoting Aβ clearance and degradation, as well as improved mouse behavior, as evaluated in T-maze, novel object recognition, and Morris water maze tests [36]. Therefore, CTS and TC extracts alone or in combination may have protective effects within the nervous system, and the CTS–TC combination might exert synergistic effects on neuroprotection.
The present study demonstrated the protective effect of the CTS–TC combination against Sco-induced memory impairment through the regulation of pathways related to inflammatory and cholinergic signaling. The components in the extract of the CTS–TC combination were chromatographed and quantified, suggesting that these compounds may be the major phytochemicals in the CTS–TC combination responsible for memory protection.
The T-maze test is commonly used to evaluate the spatial working memory [42]. It has been reported that working memory performance is sensitive to central muscarinic blockade induced by Sco [43]. A previous study using the Arena task revealed that Sco disrupts spatial memory in humans using an Arena task [44]. Memory loss and cognitive dysfunction also have been observed in the Sco-injected mice [45]. Consistent with a previous study, Sco decreased space perceptive ability, which was reversed by the CTS–TC combination. Moreover, it was found that the greater the amount of TC in the combination, the higher the space perceptive ability. In addition to the T-maze test, a novel object recognition test was conducted to assess non-spatial object memory [46]. A previous study reported that Sco-induced central cholinergic dysfunction contributes to memory impairment in rodents; thus, Sco induction is a useful model for evaluating object recognition memory [47]. The ability to discriminate an item that has been previously exposed depends on the integrity of the hippocampus and the medial temporal lobe [4648]. In this study, the CTS-TC combination increased novel object recognition ability, which was reduced by Sco. These results suggest that the CTS–TC combination potentially improves spatial memory dysfunction and object cognition impairment induced by Sco.
Activation of AChE has been observed in the brains of patients with AD [49]. AChE blocks cholinergic transmission by hydrolyzing acetylcholine, which has been implicated in learning and memory [50]. Inhibiting AChE is thought to block the hydrolysis of acetylcholine to protect cognitive functions. Choi et al. [51] indicated that the CTS extract significantly decreased alcohol-induced AChE activity in the brain. Moreover, the inhibited AChE activity in mouse whole brain has been shown by the water extract of Taraxacum platycarpum [52]. In the present study, CTS, TC, and the CTS–TC combination decreased Sco-induced upregulation of AChE protein expression. In particular, the CTS–TC combination at all ratios (4:1, 1:1, and 1:4) synergistically reduced the AChE protein level compared with CTS or TC alone, indicating that the CTS–TC combination has a better effect on AChE suppression. These results suggest that the cholinergic system can be modulated by CTS–TC through suppression of AChE protein expression.
As indicated by clinical and preclinical studies, Sco can damage learning and memory functions in humans and animal models, disrupting cholinergic neurotransmission and triggering inflammatory responses in the brain [5354]. Brain inflammation has been linked to the overproduction of pro-inflammatory molecules, including nitric oxide (NO), which can initiate neurodegeneration [55]. Moreover, NO production is largely regulated by iNOS in inflammatory conditions as demonstrated previously [56]. Intraperitoneal administration of Sco (1 mg/kg) increased the total nitrite content by approximately 2-fold in the brains of Swiss mice compared to that in the control [57]. In specific pathogen-free C57BL/6N male mice, Sco injection at 2 mg/kg increased hippocampal NO levels and iNOS gene expression by approximately 1.5-fold and 1.6-fold, respectively [58]. COX, including 2 isoforms (COX-1 and COX-2), is an important enzyme in prostaglandins (PGs) synthesis. COX-1 plays a role in PGs production with a homeostatic function, whereas COX-2 promotes the production of pro-inflammatory PGs [59]. Previous studies have shown a significant elevation in COX-1 and COX-2 activities after treatment with 1 mg/kg Sco in the brains of rats [53]. It has been indicated that COX-2 was barely detected in normal conditions but highly expressed during inflammatory responses [60]. The administration of Sco (1 mg/kg) increased COX-2 mRNA levels by approximately 3-fold in ICR male mice [61]. Cho et al. [62] demonstrated that hot water extracts of TC significantly inhibited lipopolysaccharide-induced COX-2 protein expression in mice. GFAP is a biomarker of activated astrocytes that links neuroinflammatory responses and neurodegeneration [6364]. Many reactive astrocytes are associated with cognitive dysfunction in neurodegenerative diseases [6]. Sco increased the proliferation of reactive astrocytes and GFAP expression in the cerebellum and hippocampus of rats [64]. A previous study has shown increased GFAP expression in mouse brains injected with Sco [65]. In the present study, the CTS–TC combination (4:1, 1:1, and 1:4) significantly inhibited the protein expression of iNOS, COX-2, and GFAP, showing a synergistic inhibitory effect compared to CTS or TC alone. Interestingly, it was found that the greater the amount of CTS in combination, the greater the downregulation of these proteins. These results suggest a potential anti-inflammatory effect of the CTS–TC combination in Sco-induced mice. Focusing on iNOS, COX-2, and GFAP expression after Sco treatment may be helpful in developing targeted therapies for inflammation-related diseases. In addition, recent studies have suggested that plasma GFAP levels can be used as an early marker related to Aβ pathology in AD [66]. Serum GFAP has been used to diagnose and predict mild cognitive impairment-AD [67]. Therefore, GFAP levels in body fluids, including blood, could serve as a valuable biomarker for AD, reflecting the underlying neuroinflammatory processes. However, in this study, GFAP protein levels were only observed in the brain tissue. Regarding this limitation, GFAP levels will be detected in the brain tissue, plasma, serum, and human blood in the near future.
In the present study, 5 components of the CTS–TC pair were analyzed by chromatography and quantitation. High concentrations of chicoric acid and N-feruloylserotonin were found in the CTS–TC combination; N-(p-coumaroyl) serotonin and chlorogenic acid were second, and serotonin was present in a small amount. Many studies have indicated that chicoric and chlorogenic acids from the genus Taraxacum exert anti-inflammatory and neuroprotective effects [6869]. Liu et al. [68] demonstrated that chicoric acid prevents inflammation-induced memory impairment by reducing the levels of inflammatory mediators and increasing acetylcholine levels and choline acetyltransferase activity in the mouse brain, suggesting a potential anti-inflammatory effect of chicoric acid. Chlorogenic acid suppresses AChE activity in the hippocampus and frontal cortex [70]. Serotonin is a well-known neurotransmitter with multiple functions, including the regulation of mood, memory, cognition, and sleep [71]. N-feruloylserotonin and N-(p-coumaroyl) serotonin have been isolated and identified as 2 bioactive conjugated serotonins in CTS [72]. Previous studies reported that N-feruloylserotonin and N-(p-coumaroyl) serotonin negatively affect neuronal oxidative stress [73], inflammation [74], and adipogenesis [75]. Kim et al. [74] reported that both N-feruloylserotonin and N-(p-coumaroyl) serotonin inhibit lipopolysaccharide-induced inflammatory responses in macrophages by reducing the levels of NO and pro-inflammatory cytokines. N-feruloylserotonin suppresses the inflammatory response by neutralizing the activity of the chemokine CCL22 and inhibiting the level of COX-2 [76]. Moreover, N-feruloylserotonin and N-(p-coumaroyl) serotonin isolated from CTS may exert anti-arthritic effects by blocking the nuclear factor-ĸB signaling pathway [77]. Although the memory improvement effects of CTS–TC combination against Sco may be attributed to these active components, the exact mechanism remains to be explored.
However, in the present study, animal models were used at the age of 6 weeks, which is a period of neural development. Despite the refinement of synaptic connections and myelination during this period, mice are not fully mature, and their neurons proliferate and migrate to their general locations [78]. Another study reported the onset of myelination, including in the hippocampus and corpus callosum, mostly at 2 weeks of age, and mouse neurons progressively upregulated maturation markers over 20 days [7980]. A previous study related to Parkinson’s disease demonstrated that adolescent mice (1-mon-old) also induced dramatic spine elimination and formation by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyriding (a neurotoxin that leads to the selective loss of dopaminergic neurons), and this result was significantly increased in the motor cortex of adult mice, indicating that both young and adult mice can effectively mimic the disease [81]. Therefore, rodents at 6 weeks of age can be chosen for neurodegenerative disease models, with the reason that they can significantly represent a critical developmental period and allow researchers to observe disease progression and develop therapeutic interventions in an early stage of the disease. The present study explored neurodegenerative disease as an early disease mechanism; however, further studies are required to investigate later-stage pathology in adult animal models.
The present study showed that the CTS–TC combination improved Sco-induced memory impairment, as demonstrated by behavioral tests, including the T-maze and novel object recognition tests. Moreover, the CTS–TC combination synergistically downregulated the expression of inflammation-related proteins (iNOS, COX-2, and GFAP) and inhibited AChE protein expression in Sco-treated mice. These protective effects may be supported by the presence of active compounds (chicoric acid, N-feruloylserotonin, N-(p-coumaroyl) serotonin, chlorogenic acid, and serotonin) in the CTS–TC combination, which exert potential activities in modulating cholinergic function and inflammation. Thus, The CTS–TC combination may be considered a potential agent for memory improvement. Considering these limitations, mechanistic studies of CTS–TC combinations in different signaling pathways need to be conducted.
Go to :
 XML Download
XML Download