

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Poly-γ-glutamic acid (γ-PGA) is a natural polymer composed of glutamate monomers linked with amid bonds between α-amino and γ-carboxylic acid groups. This compound can be synthesized by Bacillus species and isolated from certain fermented soybean products [1]. Several studies have suggested that γ-PGA consumption potentially contributes to increased intestinal calcium absorption [2], supporting the immune system [34], improving glucose metabolism [5], and enhancing sleep quality [6]. Although the biological mechanisms underlying these beneficial effects remain uncertain, γ-PGA has been utilized in industrial applications, for example, as an ingredient in functional foods, a food-thickening agent, a humectant in cosmetic products, and a biomaterial for drug delivery, because of its molecular properties [1]. In a quest to elucidate its potential biological mechanisms, recent studies have proposed that γ-PGA may play a prebiotic role in the gut [78]. A clinical trial revealed the diversity of the gut’s microbial community after the supplementation with a prebiotic mixture of γ-PGA and galactooligosaccharide; nevertheless, it did not demonstrate the individual effects of each prebiotic on the altered microbial composition [8].
Our prior intervention study indicated that sleep quality may be improved by the supplementation with a γ-PGA-vitamin B6 mixture [6]. We had speculated that γ-PGA supplementation potentially enhances the intestinal absorption of vitamin B6, which is essential for the synthesis of serotonin and melatonin to induce sleep; nevertheless, we did not observe any significant post-supplementation changes in serum serotonin levels.
Therefore, to determine the association between the prebiotic properties of γ-PGA and the microbiota-gut-brain axis related to sleep regulation, the current study aimed to ascertain whether γ-PGA-vitamin B6 supplementation can modify a host’s microbiota profile. In particular, this study analyzed microbiota profiles constructed using urinary assays of microbe-derived extracellular vesicles (EVs). These EVs are nanometer-sized particles secreted by gut microbes; thus, they are expected to provide information regarding human gut microbiota [9]. Furthermore, they have gained attention as an emerging biomarker reflecting a host’s microbiota-associated health, including brain health [10111213]. Accordingly, we utilized urinary EVs to determine whether the intervention alters human microbiota.
Go to : 
SUBJECTS AND METHODS
Study design and participants
This intervention study was designed as a randomized, double-blinded, placebo-controlled trial. As part of the screening process, participants, who were recruited via posters and flyers, underwent an initial interview via telephone according to the inclusion and exclusion criteria, which were as follows. The study included male and female Korean adults who were aged 20–65 yrs, free of chronic diseases (e.g., cardiovascular disease, diabetes mellitus, cancer, hepatic disease, kidney disease, etc.) and other illnesses (e.g., insomnia, food allergy, anemia, and any infection in the preceding 3 months), not pregnant, and not taking any medication or dietary supplements (including probiotics) regularly.
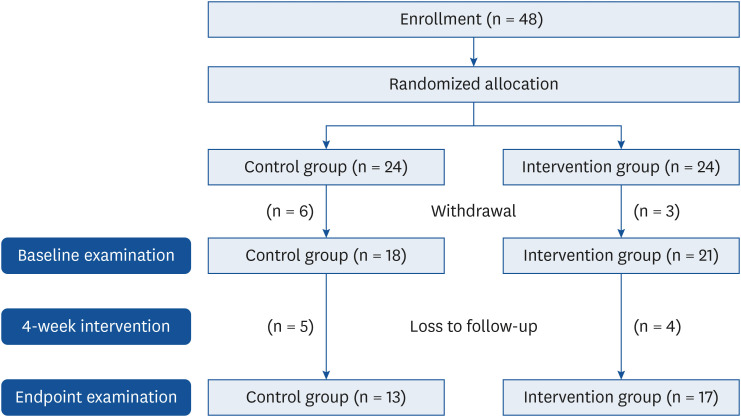
Fig. 1 shows the number of participants who were enrolled, withdrew, and were followed up. The study procedures were scheduled as follows; recruitment and enrollment of participants (from November to December 2018), intervention (from January to April 2019), and assays of biomarkers and microbiome sequencing (from May to December 2019). A total of 48 participants were enrolled after the screening process and randomly and evenly assigned to control and intervention groups. Amongst them, 39 participants (18 and 21 participants in the control and intervention groups, respectively) signed an informed consent form, which was approved by the Kookmin University Institutional Review Board (approval number: KMU-201808-HR-184), and they were subsequently engaged in a 4-week intervention study. Finally, 17 (follow-up rate: 81%) and 13 (follow-up rate: 72%) participants from the intervention and control groups, respectively, completed the intervention and provided urine samples at baseline and endpoint. Reasons of dropouts included time and scheduling conflicts and inconvenience related to study site visits. According to an earlier study [7], a sample size of 20 participants (10 in each group) provides 90% power at a two–sided significance level of 5% for the non–parametric test, as calculated using the G*Power program (version 3.1.9.2; Heinrich-Heine-Universität, Düsseldorf Universität, Germany). Therefore, the sample size of our intervention study was assumed to yield statistically significant results.
Placebo and γ-PGA-vitamin B6 supplementation
γ-PGA (powder form; Vedan Enterprise Corporation, Taichung City, Taiwan) and vitamin B6 (tablet form; Junsei Chemical Co., Tokyo, Japan), which was ground and mixed with the γ-PGA powder, were purchased for the study. The daily dose of vitamin B6 was 100 mg, which corresponds to the tolerable upper intake for female and male adults according to the Dietary Reference Intakes for Koreans [14]. The daily dose of γ-PGA was 600 mg, which is below the level approved by the Ministry of Food and Drug Safety [15], although no studies have reported any toxic side effect of γ-PGA, rendering its use potentially safe. The daily consumption of cornstarch powder, which was selected as a placebo because of its similar color and texture and purchased at a local supermarket, was 700 mg, the same quantity as that of the supplementation dose.
Every participant received a calendar with 30 pockets, each of which contained an envelope with the daily amount of γ-PGA-vitamin B6 supplement or cornstarch powder, and they were instructed to take one envelope every night 1 h before going to sleep, as instructed in the prior study [6], which focused on the sleep-related outcomes.
Questionnaire survey and anthropometric measurements
A questionnaire-based survey was administered at baseline to acquire sociodemographic and behavioral information including smoking status, alcohol consumption, and physical activity data. Smoking and alcohol consumption statuses were categorized into “abstainers” and “current/former consumers.” Physical activity was classified into five categories based on its intensity, and metabolic equivalents were subsequently calculated.
Anthropometric measurements were conducted at baseline, with participants wearing light indoor clothes. Body weight and height were measured to the nearest 0.1 kg and 0.1 cm, respectively, and body mass index was calculated as the weight (kg) divided by the height (m) squared.
Urine sample collection and assays
In both the baseline and endpoint examinations, clean-catch midstream urine (≥ 2 mL) was collected in the morning from participants who had fasted for ≥ 8 h, and sample tubes were subsequently stored in a deep freezer (−80°C). The frozen samples were transferred to a commercial laboratory for bacterial EV assays, which were conducted according to the laboratory’s standard protocol. The methods of isolating microbe-derived EVs, extracting DNA, sequencing bacterial 16S ribosomal DNA, and assigning taxonomy were similar to those reported in an earlier study [16].
Statistical analysis
Before statistical analysis, microbiome data were rarefied to an even depth of 1,069 reads per sample to eliminate the effect of variance in sequencing depth. Alpha-diversity (α-diversity) indices, including observed operational taxonomic units (OTUs) and the Chao1, Shannon, Simpson, and phylogenetic diversity (PD) indices, were calculated and compared between the two groups. These indices provide information on the richness and evenness of a microbial community. Observed OTUs as well as the Chao1, Shannon, and PD indices are used to measure richness, while the Shannon and Simpson indices are used to evaluate evenness. To compare the pairwise dissimilarity of the microbial communities observed in the baseline and endpoint data, beta-diversity (β-diversity) indices, such as the Bray-Curtis and Jaccard indices, were employed. In particular, the Bray-Curtis index indicates microbial abundance, while the Jaccard index provides the presence-absence information of the taxa.
Descriptive statistics were generated and presented as the mean ± standard deviation or proportions. Data comparison between the control and intervention groups was performed using non-parametric analysis methods, such as the Wilcoxon rank-sum test, Wilcoxon signed-rank test, and Fisher's exact test. Permutational multivariate analysis of variance (PERMANOVA) was conducted for β-diversity indices. This statistical method is based on the distance matrix and used to evaluate variability in microbial community composition between two conditions. Principal coordinate analysis (PCoA) of the distance matrix was performed to construct plots illustrating sample distribution; a close distance between symbols indicates the similarity of microbial communities.
Statistical analyses were performed using SAS software (SAS 9.4; SAS Institute, Cary, NC, USA) and R statistical software (v3.5.3; RStudio Inc., Boston, MA, USA). In all tests, a two-sided P-value less than 0.05 (P < 0.05) was considered statistically significant.
Go to :
RESULTS
Participant baseline characteristics
The baseline characteristics of the 39 participants who were included in the intention-to-treat analysis as well as those of the 30 participants who completed the intervention study and were included in the per-protocol analysis are summarized in Table 1. On comparing demographic variables and lifestyle factors between the two groups, no significant difference was observed in both analyses.
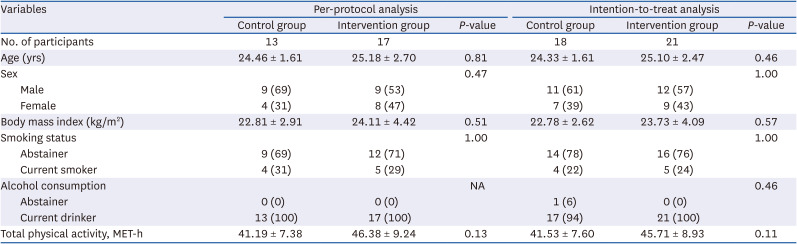
Table 1
Comparison of baseline characteristics between the control and intervention groups according to analytical methods
Values are presented as mean ± SD or number (%). The Wilcoxon rank-sum test or Fisher’s exact test was used.
NA, not applicable; MET-h, metabolic equivalent.
![]()
Comparison of the α- and β-diversity indices of the urine samples’ microbial communities
Table 2 displays the baseline and endpoint α-diversity indices. No significant difference between the baseline and endpoint data was observed in both the control and intervention groups; nevertheless, in the intervention group, all indices tended to increase from baseline, while in the control group, the observed OTUs and Simpson indices remained relatively constant, and the Chao1 index tended to decrease from baseline.
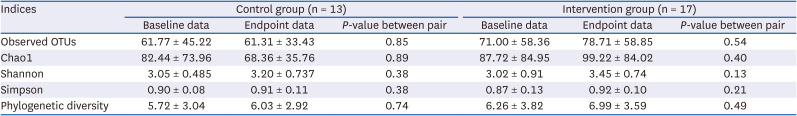
Table 2
Comparison of the α-diversity indices between baseline and endpoint in the control and intervention groups
Values are presented as mean ± SD or number (%). The Wilcoxon rank–sum test or Fisher’s exact test was used.
OTU, operational taxonomic unit.
![]()
Table 3 presents the results of PERMANOVA, which evaluated the placebo and supplementation effects in the control and intervention groups, respectively, by comparing microbial community composition between the baseline and endpoint data. The supplementation effect was exclusively significant in the β-diversity analysis based on the Bray-Curtis and Jaccard indices (P < 0.05). These results indicate the dissimilarity in microbial community composition between the baseline and endpoint data of the intervention group.

Table 3
β-diversity analysis comparing microbial community composition between baseline and endpoint data
![]()
Fig. 2 shows the PCoA plots, which illustrate the sample distribution of the microbial community between the baseline and endpoint data for each group. Baseline data symbols were more likely to be dispersed from those of the endpoint data in the intervention group.
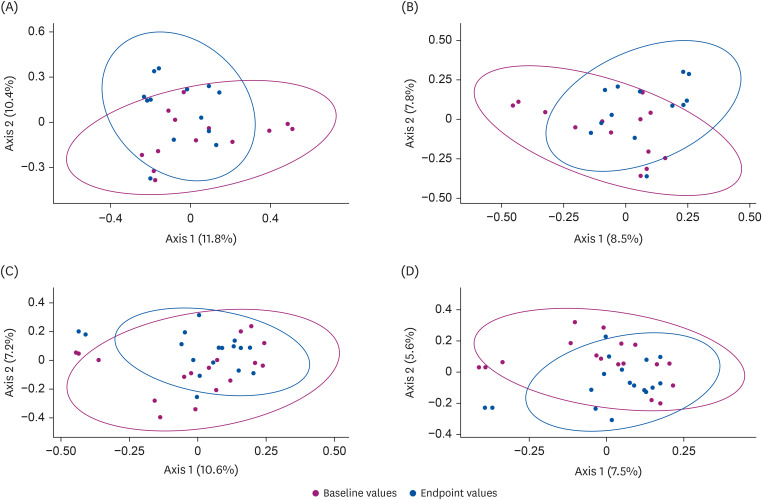 | Fig. 2Principal coordinate analysis of the β-diversity indices in the control and intervention groups. The scatter plots represent principal coordinate 1 (Axis 1) vs. principal coordinate 2 (Axis 2), with variations explained by the components indicated. Samples are indicated by points and colored according to baseline (red) and endpoint (blue) data. The results of the β-diversity indices, namely, the Bray-Curtis (A) and Jaccard (B) indices for the control group as well as those of the Bray-Curtis (C) and Jaccard (D) indices for the intervention group are presented.
|
Comparison of phylum-level microbial community composition in the urine samples
Table 4 compares the mean changes in phylum-level microbial community composition from baseline between the control and intervention groups. The relative abundance values of four dominant phyla, namely, Actinobacteria, Bacteroidetes, Firmicutes, and Proteobacteria, and certain minor phyla, including Cyanobacteria, Deinococcus-Thermus, Saccharibacteria, Verrucomicrobia, and residual miscellaneous phyla, are listed therein. The mean composition changes were not significant within each group, except the Proteobacteria composition of the control group, as well as between the groups. However, the mean changes in the Firmicutes to Bacteroidetes ratio (FB ratio) were significantly different between the control and intervention groups (P < 0.05); the FB ratios decreased from baseline in the intervention group but increased in the control group.
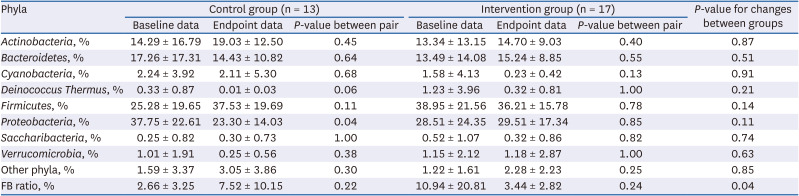
Table 4
Comparison of phylum-level microbial composition between baseline and endpoint data according to the control and intervention groups
![]()
Fig. 3 depicts the relative abundance of the four dominant phyla and a combined phylum group, which included minor phyla, for each participant. In the control group, the relative abundance of Firmicutes tended to increase, whereas that of Proteobacteria and Bacteroidetes decreased from baseline. These results are coherent with those shown in Table 4. In the intervention group, though the mean composition of Bacteroidetes slightly increased from baseline (Table 4), changes in the relative abundance of this phylum changes were quite diverse (Fig. 3). As illustrated in Fig. 3, five participants exhibited a lack of the Bacteroidetes phylum in their baseline data; however, all demonstrated the presence of the Bacteroidetes phylum after supplementation.
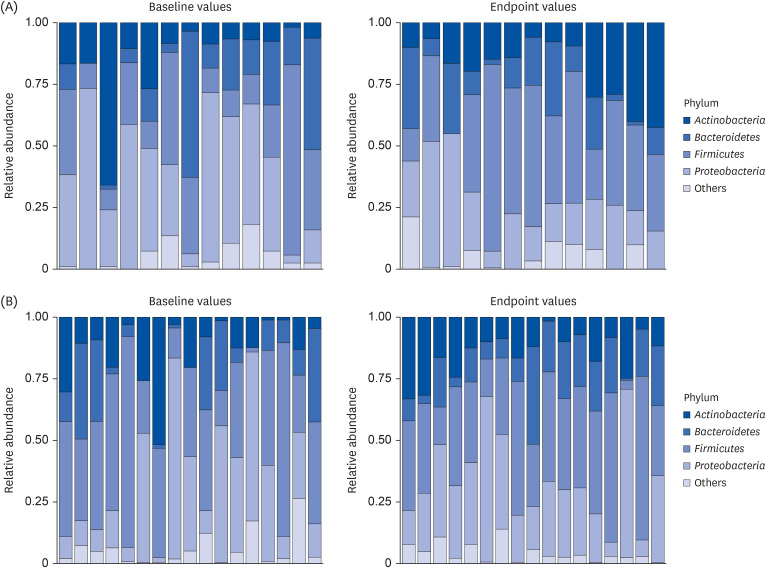
Go to :
DISCUSSION
This intervention study investigated the effects of 4-week γ-PGA-vitamin B6 supplementation on human microbiota profiles derived from urinary EV assays. The results revealed that the β-diversity indices, suggesting the variation in bacterial composition, changed significantly after supplementation in the intervention group without significant changes in the count of species indicating α-diversity. On analyzing phylum-level microbiome data, the FB-ratio changes from baseline significantly differed between the intervention and control groups.
Dysbiosis refers as the disruption of gut microbiome homeostasis, partly characterized by loss of microbial diversity; in contrast, microbial abundance strengthens the gut environment against the external milieu, thus maintaining a healthy gut [17]. Accumulating data reveal a link between dysbiosis and several diseases, including inflammatory bowel disease, obesity, and type 2 diabetes [181920]. Furthermore, based on the gut-brain axis hypothesis, dysbiosis may be associated with psychiatric disorders and sleep disturbance [2122]. The FB ratio, which is generally considered a marker of gut dysbiosis, has been reported to be positively associated with aging, cardiovascular disease, and obesity [2324]; however, it has demonstrated inconsistent association with sleep quantity and quality in human studies [25]. In an animal study, mice subjected to sleep fragmentation exhibited increased Firmicutes and decreased Bacteroidetes abundance [26].
Our prior intervention study indicated that supplementation with a γ-PGA-vitamin B6 mixture improved sleep quality [6]. To explore the potential biological mechanism supporting these previous findings, the current study analyzed the effects of the same supplementation on participants’ microbiota profiles, expecting the prebiotic role of γ-PGA in the gut. Consistent with this study’s findings, the microbial profiles of γ-PGA-administrated mice in an experimental study differed from those of the controls, especially regarding the relative abundance of Lactobacillales and Bacteroidales, which belong to the Firmicutes and Bacteroidetes phyla, respectively [7]. These two phyla are reportedly associated with the production of γ-aminobutyric acid (GABA), which is a brain neurotransmitter that plays a central role in physiological sleep regulation by inhibiting histamine-induced over-excitation in the brain and reducing wakefulness [2728]. The gut microbiota may contribute to γ-PGA degradation, producing glutamic acid, from which GABA is synthesized through the activation of glutamic acid decarboxylase (GAD) [29]. Both GABA and GAD are reportedly produced by gut microbiota [30]. Besides the GABA synthesis mechanism, γ-PGA-containing supplement consumption may modify a host’s microbiota, causing the production of proteins that directly affect sleep regulation. In an animal study, the administration of EVs from Prevotella histicola increased non-rapid eye movement sleep, which includes a deep sleep stage [31].
Microbe-derived EVs, which are spherical particles with a diameter < 300 nm, are released from the cell surface into the extracellular milieu. They contain bioactive molecules, such as DNA, RNA, proteins, and metabolites, and possibly interact with host cells [9]. Accumulating data have revealed that microbe-derived EVs are found in human urine and blood [32] and can be utilized as biomarkers that reflect commensal microbes and a host’s health traits [10111213].
This study has certain limitations. First, the lack of significance in the α-diversity index results might have emanated from the small sample size. Second, the average follow-up rate was 77% after the baseline examination, and loss to follow-up might have impacted the planned randomization. However, the intention-to-treat and per-protocol analyses revealed no significant difference in the participant’s baseline characteristics between the groups. Third, we evaluated the effects of a mixture supplement containing γ-PGA and vitamin B6 on microbiota profiles, potentially linked to sleep improvement based on our prior report [6], but we were unable to assess such effects of exclusive γ-PGA supplementation because of limited study budget. The current intervention study had the following strengths: 1) it applied a randomized, double-blinded, placebo-controlled design; 2) pre- and post-intervention microbiota profile analyses were performed; and 3) to the best of our knowledge, it is the first trial to investigate the association between γ-PGA-containing supplementation and human microbiota profiles. Further studies involving a large sample or population characterized by various ethnicities are warranted.
Go to :
 XML Download
XML Download