

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Cladophora glomerata (CG) is a freshwater macroalga belonging to the phylum Chlorophyta. CG is grown widely in the Nan River, North Thailand, under the common name “Kai” and is an important human nutrient source. The species contains high quantities of carbohydrates, fat, proteins, vitamins, and minerals [1]. In addition, a CG extract (CGE) has shown anti-inflammatory, anti-hypertensive, and antioxidant activities in vivo and in vitro [23]. A previous study reported that CGE has antidiabetic effects and protects the renal organic anion transport by modulating protein kinases C and ζ [3].
Type 2 diabetes mellitus (T2DM) is a metabolic disorder characterized by hyperglycemia and an insufficiency of endogenous insulin secretion or action [456]. Prolonged hyperglycemia may result in the development of several diabetic complications, including retinopathy, peripheral neuropathy, and nephropathy [7]. On the other hand, elevated blood glucose levels are involved in the overproduction of reactive oxygen species (ROS) and antioxidant depletion in several tissues [8]. Recent studies have indicated that hyperglycemia induces ROS production, leading to oxidative and nitrosative stresses and contributing to diabetic nephropathy (DN) [910]. Nitric oxide (NO) production and function also increase in DN patients and rats [1112].
Several mechanisms are involved in the pathogenesis of DN, including the activation of transcription factors, pro-inflammatory cytokines, chemokines, and adhesion molecules [13]. Among these, nuclear factor kappa B (NF-κB) is the most important therapeutic target in DN [13]. Recent studies have found that deactivating NF-κB by curcumin and thiazolidinediones could improve DN in streptozotocin (STZ)-induced diabetic rats [131415]. In addition, NF-κB can be activated by inflammatory cytokines, tumor necrosis factor-alpha (TNF-α), and interleukin-1β (IL-1β) [71617]. Renal NF-κB are also upregulated and associated with TNF-α, IL-1β, monocyte chemoattractant protein-1, and IL-6 production [18]. The kidney is important in eliminating endogenous and exogenous compounds by various membrane transport proteins [19]. Therefore, organic anion transport mediated by transporters is the critical step in the cellular uptake of organic anions across the basolateral membrane of proximal tubules, resulting in organic anion excretion into the tubular lumen and urine [20].
Among the organic anion transporters (Oats), Oats 1 (SLC22A6) and 3 (SLC22A8) have the highest expression levels and recognize a broad spectrum of substrates and transport with a high-affinity for typical substrate, para-aminohippurate (PAH), and various anionic drugs, such as non-steroidal inflammatory drugs, antivirals and antibiotics in exchange for dicarboxylates inside the cells [2122]. In addition, the changes in organic anion clearances are also involved in the severity and progression of DN. For example, decreased Oat1 and Oat3 gene expression in DN patients resulted in reduced urinary organic acid [11]. Renal PAH transport in T1DM in mice was decreased, corresponding to the downregulation of Oat3 expression, but not Oat1 [23]. Recent studies reported that aqueous extractions of Spirogyra Neglecta and CGE, the major members of fresh macroalgae, exhibited antidiabetic effects and protected the renal organic anion transport function [324]. Nevertheless, it is unknown if the antioxidative effects of CGE have any direct renoprotective effects against T2DM.
Therefore, this study examined the effects of CGE on renal oxidative/nitrosative stresses in T2DM rats and identified the possible mechanisms involved in preventing DN.
MATERIALS AND METHODS
Chemicals
Polyclonal rabbit anti-NF-κB p65 and goat-anti-mouse or rabbit IgG horseradish peroxidase-conjugated secondary antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Monoclonal TNF-α and polyclonal IL-1β were procured from R&D systems (Minneapolis, MN, USA). Monoclonal anti-lamin B1 was acquired from Cell Signaling (Danvers, MA, USA), and monoclonal anti-β actin was obtained from Abcam (Waltham, MA, USA). Tritiated para-aminohippurate ([3H]-PAH; specific activity 1 Ci/mmol) was supplied by PerkinElmer Life Sciences (Branford, CT, USA). STZ and CelLytic™ MT cell lysis reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA), and vitamin C (VC) was obtained from Merck (Darmstadt, Germany). All other chemicals of high purity grade were obtained from commercial sources.
CGE preparation, purification, and qualification
CGE was previously identified [3]. A voucher specimen (number AARL G048) was deposited at the herbarium of the Applied Algal Research Laboratory, Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai, Thailand. The CGE was prepared, purified, and quantitated, as shown in a previous study [3]. In addition, the total phenolic content of CGE was quantified to reach a minimum of 8.36 ± 0.13 mg gallic acid equivalent/g of extract before use in this study.
Animals and induction of diabetes in rats
Male Wistar rats weighing 120–150 g were obtained from the National Laboratory Animal Center, Mahidol University, Salaya, Thailand. The Laboratory of Animal Care and Use Committee at the Faculty of Medicine, Chiang Mai University, Chiang Mai, Thailand, approved the animal facilities and protocols (protocol number 12/2553). All rats were housed in a room maintained at 25 ± 1°C at a 12:12 h dark-light cycle. A previous study reported the anti-hyperglycemic effect of CGE [2]. Therefore, the dose of 1,000 g/kg body weight (BW) of CGE was also used in this study. The animals were divided randomly into 6 groups: normal diet (ND), normal diet supplemented with CGE (ND + CGE), T2DM (DM), T2DM supplemented with CGE at 1,000 mg/kg BW (DM + CGE), T2DM treated with positive control antioxidant, and VC at the dose of 200 mg/kg BW (DM + VC), as described elsewhere [24]. During the initial 2 weeks, the high-fat diet (58% calories as fat) rats were injected intra-peritoneally with a low single dose of 40 mg/kg BW of STZ, as reported elsewhere [24]. Ten days after the injection, the rats in which the fasting blood glucose levels exceeded 250 mg/dL were considered T2DM. All non-diabetic rats were excluded from this study. Each rat received either CGE or VC, daily administered either via a vehicle (distilled water), CGE or VC by oral gavage for 12 weeks until sacrifice.
Determination of plasma glucose, triglyceride, and insulin levels
Hyperglycemia, hyperlipidemia, and insulin resistance were measured to determine the characteristics of T2DM. The total plasma glucose and triglyceride levels were analyzed by commercial enzymatic colorimetric assays (Biotech Reagent, Bangkok, Thailand). In contrast, the plasma insulin concentration was obtained using a Sandwich ELISA assay kit from LINCO Research (Millipore, MA, USA). The homeostasis model assessment of insulin resistance (HOMA-IR) was calculated using the following formula: fasting plasma insulin (µU/mL) × fasting plasma glucose (mmol/L)/(22.5).
Determination of total, reduced, and oxidized plasma glutathione (GSH) levels
The effects of CGE on plasma total, reduced, and oxidized GSH were examined by performing colorimetric assays according to the manufacturer’s protocol (Cayman Chemical, Ann Arbor, MI, USA). The plasma samples were deproteinated by adding 5% metaphosphoric acid (MPA) and centrifuging at 3,500 g for 10 min. The supernatant was added to the assay buffer, and the MPA extract samples were collected for GSH evaluation.
Determination of the total malondialdehyde (MDA) level in renal cortical tissue
The renal oxidative stress condition was examined by measuring the total MDA level according to the manufacturer’s protocol (Cayman Chemical). Briefly, renal cortical tissues were cut and suspended in CelLytic™ MT cell lysis reagent containing protease inhibitors (Roche Applied Science, Indianapolis, IN, USA) according to the manufacturer’s protocol (Sigma-Aldrich). The tissues were then homogenized and centrifuged at 1,600 × g for 10 min at 4°C. The supernatant was then collected for MDA measurement. Each sample was expressed as the total MDA level to total protein concentration (ηmol/mg protein).
Quantitative real-time polymerase chain reaction (qPCR) analysis
The total RNA was extracted from renal cortical tissues using the total RNA extraction kit (Amresco, Solon, OH, USA). First-strand cDNA was obtained using the iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA), and qPCR was performed using Bio-Rad iQ SYBR green supermix on Bio-Rad iQ5 (Bio-Rad). The specific primer sets for the antioxidant and β-actin genes were purchased from Integrated DNA Technologies (Coralville, IA, USA), as listed in Table 1. The expression of antioxidant genes was normalized by β-actin and reported as the relative fold changes. The qPCR amplification was performed in duplicate for each cDNA.
Table 1
Primer sequences and expected amplicon sizes for the gene amplification

Subcellular fractions and western blot analysis
Subcellular fractions were extracted from the renal cortical tissues using differential centrifugation to determine the target protein expressions in each cellular compartment, as described in a previous study [24]. Briefly, renal cortical tissues were cut and suspended in CelLytic™ MT cell lysis reagent (Sigma-Aldrich) containing 1% complete protease inhibitor cocktail (Roche Applied Science). The homogenate samples were centrifuged at 5,000 g for 10 min at 4°C, and the supernatant was specified as whole cell lysate. The pellet was re-suspended in the same solution and centrifuged at 10,000 g for 10 min at 4°C. The supernatant from this step was specified as the nuclei-rich fraction. The whole-cell lysate fraction was then centrifuged at 100,000 g for 2 h at 4°C, and the supernatant from the spin was specified as the cytosolic fraction. The commercial Bradford assay (Bio-Rad) was used to measure the total protein concentration of each sample. All samples were stored at −80°C prior to use in further experiments.
For western blotting, the protein samples (50 µg/lane for nuclei samples and 100 µg/lane for whole cell lysate and cytosolic samples) were resolved in 4X Laemmli solution and separated on 10% sodium dodecyl sulfate-polyacrylamide gel. The proteins were then transferred onto a polyvinylidene difluoride (PVDF) membrane (GE Healthcare, Milwaukee, WI, USA) using the Bio-Rad system. The non-specific bindings on the membrane were then eliminated by blocking with 5% non-fat dry milk in 0.05% Tween 20 in Tris-buffered saline (TBS-T) for 1 h at 4°C. The membrane was incubated overnight with the desired specific primary antibodies against NF-κB p65, TNF-α, and IL-1β. An anti-lamin B1 antibody was applied to confirm the enrichment of the nuclei fraction, whereas an anti-β actin antibody was also used as a loading control for all samples. The PVDF membranes were washed with TBS-T and incubated with goat-anti-mouse or rabbit IgG horseradish peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology) for 1 h at 4°C. The target proteins were then detected using an enhanced chemiluminescent kit (GE Healthcare, Buckinghamshire, UK) and quantitatively analyzed by the Image J program from the Research Services Branch of the National Institute of Mental Health (Bethesda, MD, USA).
Renal slice preparation and transport study
The antioxidative effect of CGE on renal transport activity was examined by measuring the PAH uptake in renal cortical slices, as previously described [25]. Briefly, rat kidneys were removed and placed in an oxygenated saline buffer; the renal cortical slices (≤ 0.5 mm; 5–15 mg wet weight) were then cut with a Stadie–Riggs microtome and maintained in ice-cold oxygenated modified Cross and Taggart buffer containing the following (mM): 95 NaCl, 80 mannitol, 5 KCl, 0.74 CaCl2, and 9.5 Na2HPO4, pH 7.4. The renal slices were pre-incubated in modified Cross and Taggart buffer in the absence or presence of 5 mM sodium nitroprusside (SNP) for 30 min and then incubated in a buffer containing 5 μM [3H]-PAH for 30 min at reverse transcription. The uptake was stopped by adding ice-cold buffer. The slices were then washed, blotted, weighed, dissolved in 1 N NaOH, and neutralized with 1 N HCl. A scintillation fluid was added, and radioactivity was measured using a liquid scintillation analyzer (PerkinElmer Life Sciences, Hopkinton, MA, USA). The uptake of [3H]-PAH was calculated as the tissue to medium ratio, i.e., (DPM/g tissue)/(DPM mL medium).
Quantification of total NO level
The production of NO induced by SNP was determined by measuring the nitrite and nitrate levels using chemiluminescence assays. The slices were pre-incubated in 0.5 mL of buffer in the absence or presence of 5 mM SNP for 30 min, and the tissues were homogenized and centrifuged as described above. The nitrite and nitrate levels were measured using a slight modification of the methodologies reported elsewhere [2627]. Briefly, nitrite, iron-nitrosyl, and S-nitrosothiols were reduced to NO gas using a triiodide solution. In contrast, all NO metabolites, including nitrate, were reduced to NO gas using a vanadium III chloride solution at 95°C [2627]. The emitted photons were detected and quantified by a photomultiplier tube of a chemiluminescence-based NO analyzer (Eco Medics AG, Duernten, Switzerland). Sodium nitrite and sodium nitrate were used to produce standard curves for nitrite and nitrate measurement, respectively. The NO levels were calculated from the area under the curve using Origin7 (Origin Lab, Northampton, MA, USA). The value from the vanadium III chloride-based assay was subtracted from the nitrite level from the triiodide-based assay to yield the nitrate level. The nitrite and nitrate levels are reported as the total NO production.
Statistical analysis
The data are reported as mean ± SE of the mean. Statistical differences were assessed using a one-way analysis of variance, followed by a Tukey-Kramer test. The SNP-induced NO production was analyzed using an unpaired, 2-tailed Student’s t-test. Statistical analyses were conducted using the Statistical Package for the Social Sciences (SPSS) version 11.5 (SPSS Inc., Chicago, IL, USA). A P-value of 0.05 was considered significant.
RESULTS
Effects of CGE on general characteristics of T2DM rats
The BW, kidney weight per BW ratio, food consumption, and water intake were similar among the experimental groups (Table 2). The fasting plasma glucose, triglyceride, and HOMA-IR values were significantly higher in the T2DM rats than in the control. In contrast, these parameters were markedly lower in the CGE-treated rats than in the T2DM rats. In addition, the DM + VC group showed significantly reduced triglyceride levels without changes in the glucose levels compared to T2DM rats. Despite no significant difference in the plasma insulin among experimental groups, the HOMA-IR was significantly lower in the CGE-treated rats than in those with T2DM. Moreover, the plasma parameters were not altered in ND + CGE compared to the control. These results suggest that CGE improved the diabetic conditions without affecting the biochemical parameters under normal conditions.
Table 2
Effects of CGE on the general characteristics of T2DM experimental rats
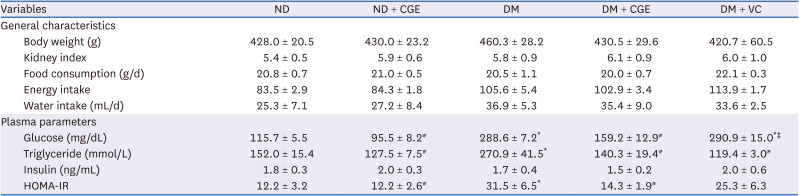
Data are expressed as the mean ± SE from 6 to 8 animals per group.
CGE, Cladophora glomerata extract; T2DM or DM, type 2 diabetes mellitus; ND, normal diet; VC, vitamin C; HOMA-IR, homeostatic model assessment of insulin resistance.
*P < 0.05 indicates the significant differences from ND; #P < 0.05 indicates the significant differences from DM group; ǂP < 0.05 indicates the significant differences from DM + CGE rats.
Effect of CGE on plasma antioxidant levels and renal cortical oxidative stress
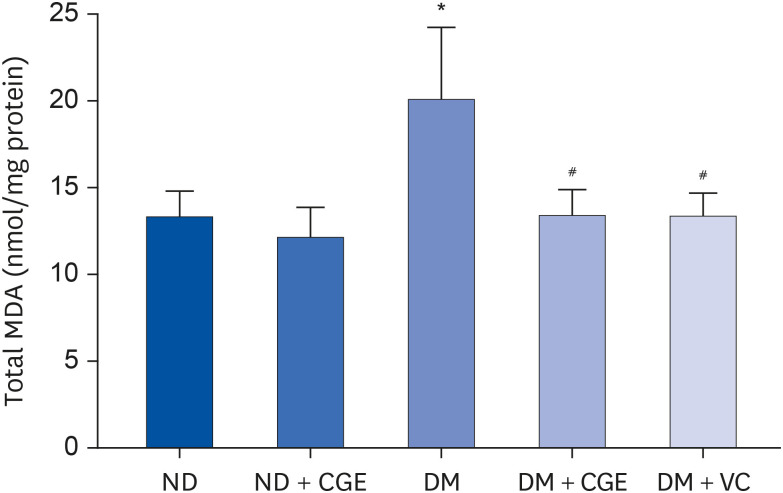
The total, oxidized, and reduced GSH levels were similar in all experimental groups (Fig. 1). In addition, the diabetic rats had significantly higher renal MDA concentrations than the control (Fig. 2), whereas the MDA level was lower in the CGE and VC-treated rats. Furthermore, CGE did not alter the MDA level in normal rats. These results suggest that the CGE could reduce renal oxidative stress in T2DM.
Fig. 1
Effects of CGE on the plasma total, reduced, and oxidized GSH levels, determined in plasma samples from each experimental group using a colorimetric assay kit. The results are expressed as the mean ± SE of the mean (n = 6).
CGE, Cladophora glomerata extract; T2DM or DM, type 2 diabetes mellitus; ND, normal diet; VC, vitamin C; GSH, glutathione.
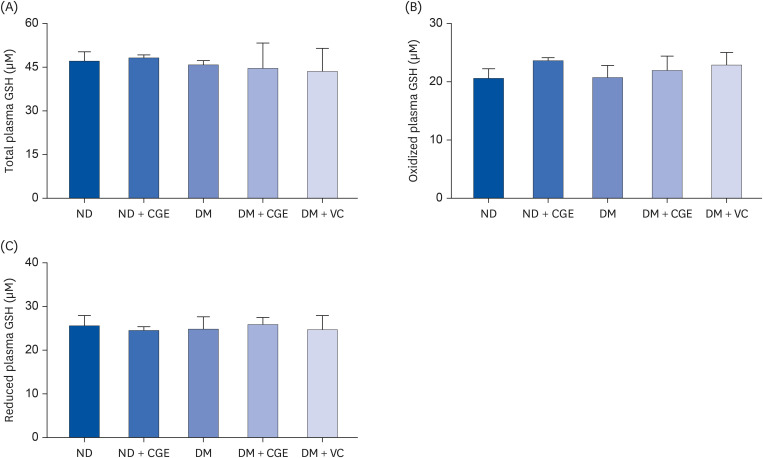
Fig. 2
Effects of CGE on renal cortical MDA level, determined in renal cortical tissue homogenates from each experimental group using a commercial TBARS assay kit. The results are expressed as mean ± SE of the mean (n = 6).
CGE, Cladophora glomerata extract; T2DM or DM, type 2 diabetes mellitus; ND, normal diet; VC, vitamin C; MDA, malondialdehyde.
*P < 0.05 indicates a significant difference from ND; #P < 0.05 indicates a significant difference from DM rats.
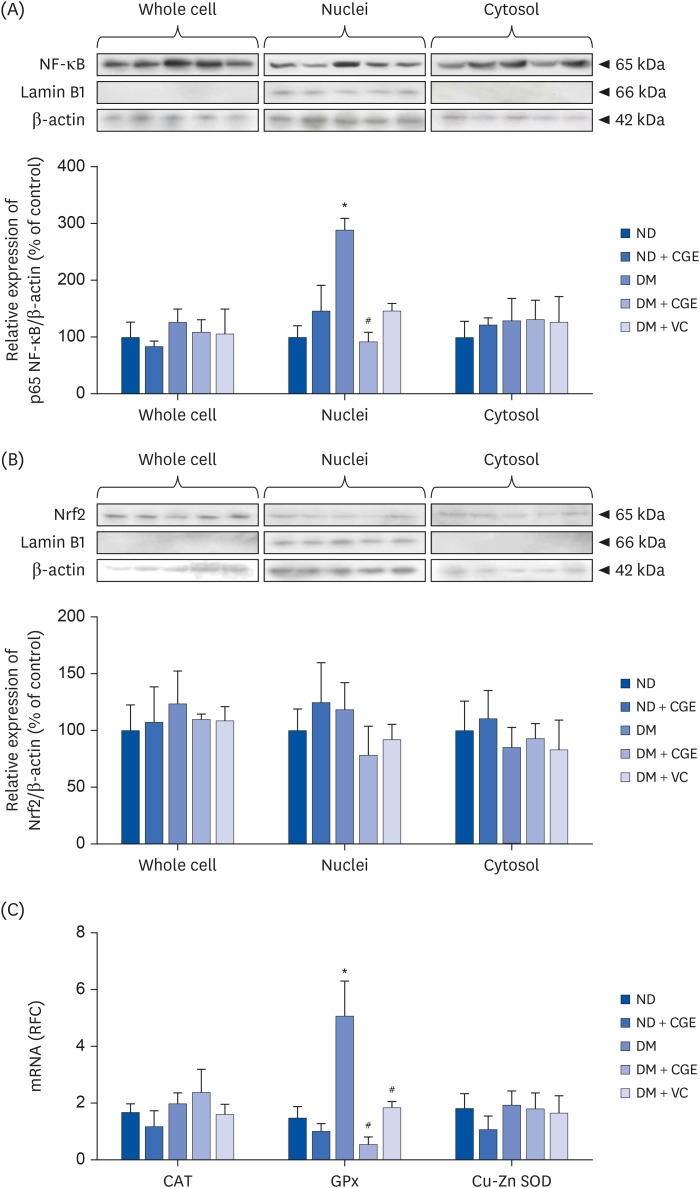
Effect of CGE on renal antioxidant
This study further investigated the effects of CGE on oxidative stress markers, including NF-κB, nuclear factor (erythroid-derived 2)-like 2 (Nrf2), catalase (CAT), glutathione peroxidase (GPx) and superoxide dismutase (SOD). Based on a previous study, hyperglycemia, free fatty acid, and oxidative stress were the major stimulators of NF-κB [28]. The levels of NF-κB active subunit (p65 nuclear factor κB [p65NF-κB]) expression in each subcellular fraction extracted from the renal cortical tissues were quantified using western blotting analysis. NF-κB activation (p65NF-κB) was significantly higher in the nuclei fraction of T2DM rat kidneys than in the control (Fig. 3A), whereas CGE and VC markedly reduced its activation. By contrast, there were no significant differences in Nrf2 protein expression in all cell compartments (Fig. 3B). In addition, the T2DM rats showed significantly higher renal GPx gene expression than the control. In contrast, CGE and VC could normalize GPx expression relative to that of T2DM without affecting CAT and SOD expression, suggesting that CGE affected the induction of antioxidant enzyme transcription (Fig. 3C). This result indicates that CGE improved oxidative stress in T2DM by preventing the activation and translocation of NF-κB.
Fig. 3
Effect of CGE on the expression of stress-sensitive markers. (A) p65NF-κB expression in subcellular fractions of rat kidneys. Anti-p65NF-κB antibody was then detected, whereas anti-β-actin and anti-lamin B1 antibodies were used as a loading control and nuclei marker, respectively. The data are expressed as mean ± SE of the mean and repeated for separate sets of animals (n = 3). p65NF-κB protein expression is shown at the top, and the quantification of relative protein expression in each fraction is at the bottom. (B) Nrf2 expression in the subcellular fractions of the rat kidneys. Anti-Nrf2 antibody was subsequently detected, whereas anti-β-actin and anti-lamin B1 antibodies were used as a loading control and nuclei marker, respectively. The data are expressed as the mean ± SE of the mean and repeated for separate sets of animals (n = 3). Nrf2 protein expression is shown at the top and quantification of the relative protein expression in each fraction at the bottom. (C) mRNA expression levels of CAT, GPx, and Cu-Zn SOD from experimental rat kidneys. The values represent mean ± SE of the mean (n = 6).
CGE, Cladophora glomerata extract; T2DM or DM, type 2 diabetes mellitus; ND, normal diet; VC, vitamin C; p65NF-κB, p65 nuclear factor κB; Nrf2, nuclear factor (erythroid-derived 2)-like 2; CAT, catalase; GPx, glutathione peroxidase; Cu-Zn SOD, copper-zinc superoxide dismutase.
*P < 0.05 indicates the significant differences from ND; #P < 0.05 indicates significant differences from DM rats.
Direct effect of CGE on renal inflammatory response
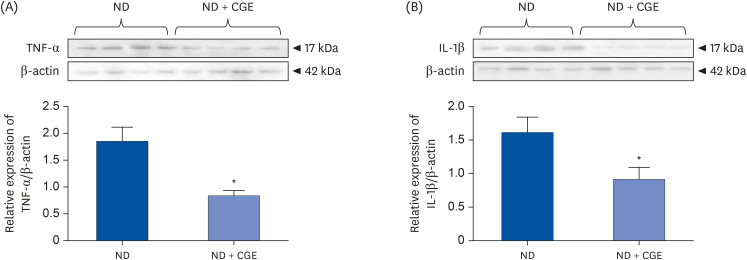
TNF-α and IL-1β protein expression in renal cortical tissues was further determined to confirm whether CGE directly affects NF-κB activation, the downstream of NF-κB. As observed in p65NF-κB, CGE significantly decreased the TNF-α and IL-1β protein expression levels in renal cortical tissues relative to normal kidneys (Fig. 4). Hence, CGE directly improved oxidative stress by reducing pro-inflammatory cytokine production.
Fig. 4
Effect of CGE on the protein expressions of pro-inflammatory cytokines. (A) TNF-α and (B) IL-1β expression in whole-cell lysate from the renal cortical tissues in ND and ND + CGE groups. Anti-TNF-α and anti-IL-1β antibodies were then detected, whereas anti-β-actin antibody was used as a loading control. The values represent the mean ± SE of the mean from separate sets of animals (n = 4).
CGE, Cladophora glomerata extract; ND, normal diet; TNF-α, tumor necrosis factor-alpha; IL-1β, interleukin-1β.
*P < 0.05 indicates significant differences from ND rats.
Effect of CGE on SNP-induced renal NO production
The data showed that the pre-incubation with SNP inhibited PAH transport mediated by Oat1 and Oat3 in the renal slices from normal rats (Fig. 5A). Interestingly, the CGE-treated normal rats did not differ in PAH uptake after SNP pre-incubation. This suggests that CGE directly prevented nitrosative stress-induced impairment of the organic anion transport function.
Fig. 5
Effect of CGE on sodium nitroprusside-induced NO production in renal cortical tissue. (A) PAH transport in the renal cortical slices from ND and ND + CGE rats. Rat renal cortical slices were incubated for 30 min in the buffer containing 10 μM of tritiated PAH in the presence or absence of SNP. Data are expressed as mean ± SE of the mean (n = 5). (B) Sodium nitroprusside induced NO production in renal cortical tissues from ND and ND + CGE rats. Rat renal cortical slices were incubated for 30 min in the buffer with or without SNP (n = 5). The data are expressed as mean ± SE.
CGE, Cladophora glomerata extract; ND, normal diet; NO, nitric oxide; PAH, para-aminohippurate; SNP, sodium nitroprusside; T/M, tissue to medium.
*P < 0.05 indicates significant differences from slices incubated with buffer alone.
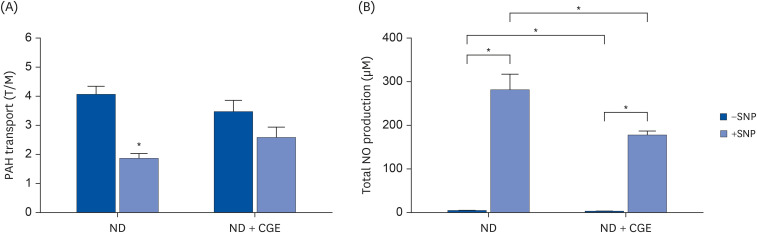
The effects of SNP in renal tissues were confirmed by measuring the total NO levels. The results showed that CGE reduced renal NO production under normal and SNP induction conditions (P < 0.05; Fig. 5B). Furthermore, an increase in NO production induced by SNP was opposed after CGE supplementation (P < 0.05). These data confirm that CGE prevented nitrosative stress, restoring the organic anion transport function.
DISCUSSION
The CGE is widely produced along the Nan River North Thailand. Previous studies have shown its beneficial effects, including antigastric ulcer, analgesic, hypotensive, anti-inflammatory, and antioxidant activities [2]. In addition, a recent study reported that CGE has antidiabetic and renoprotective effects in T2DM rats by restoring the renal organic anion transport function, but the underlying mechanisms are unclear [3]. This question was addressed by evaluating the antioxidant effects of CGE on renal oxidative stress and inflammation in the T2DM rat model. The present work shows that the antioxidant effect of CGE reduced renal cortical oxidative/nitrosative stress by inducing antioxidative gene expression, GPx, inhibiting NKB translocation, and decreasing pro-inflammatory cytokine production. These factors could prevent the impairment of renal organic anion clearance, leading to a significant decrease in the risk of DN. According to a previous study, CGE at 1,000 mg/kg BW in this study can be converted to a human dose at 9,720 mg/60 kg BW/day [28].
Hyperglycemia and hyperlipidemia are prominent causative factors in ROS production [2930]; ROS can directly damage DNA, proteins, and lipids in the cells. Previously, membrane lipids have been shown as one of the targets of ROS [31]. Lipid peroxidation produces several reactive aldehydes, such as MDA, acrolein, and 4-hydroxynonenal [32]. Among these, MDA is a primary biomarker of membrane lipid damage and oxidative stress [33]. In addition, oxidative stress contributes to the progression and severity of DN for 20–30% of types 1 and 2 diabetes patients [3435]. Thus, preventing ROS generation could delay the progression of DN. CGE can decrease renal cortical MDA levels, which could then attenuate renal oxidative stress induced by diabetic conditions. Therefore, the antioxidant effects of CGE are a primary defense mechanism for DN prevention.
According to a previous study, ROS can trigger Nrf2 translocation [36], which could induce the expression of the 3 primary antioxidant enzymes SOD, CAT, and GPx, preventing intracellular ROS production [37]. The T2DM rat model in this study showed strong expression of the GPx gene in the kidney. In agreement with the data, renal and cardiac GPx protein expression and function were higher in diabetic rat and mice models against oxidative stress [383940]. According to GSH, it is the most abundant antioxidant molecule that uses GPx as a cofactor to reduce oxidative stress molecules, such as hydrogen peroxide radicals (H2O2) [40]. In addition, upregulated GPx by ROS could induce NF-κB activation in skeletal muscle cells [41]. The NF-κB subunits are bound to the promotor region of GPx in response to oxidative stress in lipopolysaccharide-induced U937 cells [42]. This active p65NF-κB subunit induces the generation of inducible nitric oxide synthase (iNOS), resulting in NO production [43]. This partly contributes to hyperfiltration and microalbuminuria in early DN [44]. Previous studies have shown that epigallocatechin-3-gallate, isoquinoline alkaloid, and berberine prevent the transformation of renal epithelial cells to fibroblasts in DN by activating Nrf2 [4546]. Although Nrf2 protein expression did not change in any cellular compartments in this study, up-regulation of the downstream targeting protein of Nrf2 (GPx) by CGE supplementation did preserve and counterbalance renal oxidative stress in T2DM rat kidneys. Similarly, a previous study showed that curcumin normalized GPx expression in STZ-induced DM rats [38]. Moreover, flavonoids, one of the active compounds in CGE, potentially inhibited phosphoinositide 3-kinase/protein kinase B-induced eNOS function, reducing NO production [47]. Therefore, the strong inhibition by CGE is a primary defense mechanism for DN prevention.
In addition to activating renal iNOS, p65NF-κB can induce renal TNF-α and IL-1β production in diabetic rats [71617]. Both pro-inflammatory cytokines, TNF-α and IL-1β, are involved in diabetic pathogenesis and are crucial in diabetic complications [48]. The extent of inflammatory infiltration into the tissues, including macrophages and T-cells, is associated with the severity of DN [4950]. CGE also showed a direct antioxidative effect by reducing TNF-α and IL-1β production in ND treated with CGE compared to ND rats, suggesting that CGE has a strongly direct effect on proinflammatory cytokines, which are the oxidative stress markers. Several major chemical constituents in CGE exist, including isoquercetin, catechin, tannic acid, hydroquinin, quercetin, rutin, gallic acid, and kaempferol [3]. Isoquercetin blunts H2O2-induced ROS production through H2O2, hydroxyl radicals, and superoxide radicals in retinal ganglion cell 5 cells [51]. In addition, green tea catechin reduced the ROS levels by inhibiting ERK activation in the human breast epithelial cell line MCF10A [52].
As Oat1 and Oat3 play a crucial role in organic anion uptake across the basolateral membrane and are strongly expressed in the kidney [20], the function of these 2 transporters is regulated by several factors, such as hormones, endogenous/exogenous substances, pathological status, and oxidative stress condition [2053]. On the other hand, oxidative stress impairs transporter function, and nitrosative stress is generated by reactive nitrogen species that impact several transporters. For example, SNP-induced nitrosative stress reduced organic cation transporter and Na+/taurocholate co-transporting polypeptide function in isolated rat hepatocytes [54]. Similarly, nitrosative stress induced by SNP-impaired cyclosporin A transport is mediated by P-glycoprotein (P-gp) in mouse brain capillary endothelial (MBEC4) cells [55]. Similarly, SNP injection into a rat brain decreased the daunomycin efflux from the brain, suggesting that the P-gp function was impaired by nitrosative stress under in vivo and in vitro conditions [56]. These results suggest that PAH transport mediated by Oat1 and Oat3 was blunted under nitrosative stress induced by SNP. CGE could restore this defect directly. Overall, CGE has renoprotective effects against NO production, probably by directly scavenging NO molecules, leading to reduced renal organic anion transport.
This study reported the antioxidative, anti-inflammatory, and anti-nitrosative effects of CGE against impairment of renal organic anion transport mediated by Oat1 and Oat3 in experimental rats. CGE could improve renal oxidative/nitrosative stresses, restoring the Oat1 and 3 transport functions. The mechanisms through which CGE improved PAH transport involved the modulation of free radical scavenging enzymes, NF-κB, and proinflammatory cytokines. These findings facilitate the further development of CGE, making it a potential nutraceutical product for kidney diseases.
 XML Download
XML Download