

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The skin is the largest organ of the body and serves as a crucial interface between the internal and external environments, protecting against pathogens, UV radiation, and harmful chemicals. Its intricate features are partly responsible for various physiological functions. The skin comprises the epidermis, dermis, and subcutaneous tissue. The epidermis, housing keratinocytes, safeguards the skin by minimizing heat and moisture loss. In addition, it features Merkel cells, Langerhans’ cells, and melanocytes. The dermis exists beneath the epidermis and consists of fibroblasts and an extracellular matrix (ECM) with glycoproteins, proteoglycans, elastin, collagen, and hyaluronic acid [12].
Skin aging manifests in two forms: intrinsic and extrinsic aging, characterized by dyspigmentation, elasticity decline, wrinkles, and fragility. Intrinsic aging involves a thinned epidermis, cell loss, and skin wrinkling, whereas extrinsic aging, known as photoaging, results from accumulated damage caused by UV exposure [34]. Exposure to UV radiation in the epidermis induces reactive oxygen species (ROS) and reactive nitrogen species (RNS) that break down the antioxidant defense system, leading to oxidative stress. UVB-irradiated oxidative stress contributes to increased water loss, resulting in skin dryness. In addition, it stimulates the expression of matrix metalloproteinases (MMPs), leading to elastin degradation, and activates the enzyme tyrosinase, which is associated with melanin production. Moreover, it triggers protein degradation and pro-inflammatory cytokine production, culminating in the formation of skin wrinkles [567].
Indian gooseberry (Emblica officinalis), also called amla, has been investigated to understand its potential health benefits and functional properties, such as gastroprotective, antioxidant, anti-obesity, anti-cancer, anti-diabetic, and immunomodulatory effects. In addition, it has also been studied for its anti-bacterial, anti-fungal, and other medicinal properties [891011]. Barley sprouts (Hordeum vulgare L.) have been reported to enhance the phenolic bioactive-linked anti-glycemic, anti-obesity, anti-inflammatory, and anti-cardiovascular effects [121314]. The consumption of Indian gooseberry or barley sprouts has been popularized in the form of pharmaceuticals, cosmetic products, and health foods, but insufficient research on the combined health benefits and mechanism of mixtures of Indian gooseberry and barley sprout (IB) has been conducted. The authors previously investigated the anti-obesity effects of an IB complex using 3T3-L1 cells and high-fat diet-fed animal models [1516]. The current experiments investigated the mechanisms underlying the effects of IB complex on UVB-induced skin oxidative stress by analyzing the hyaluronic acid and ceramide synthesis factors, c-Jun N-terminal kinase (JNK)/c-Fos/c-Jun/MMPs and transforming growth factor-β receptor I (TGF-βRI)/small mothers against decapentaplegic homolog (Smad) pathways, and the cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA)/cAMP-responsive binding protein (CREB)/melanocyte inducing transcription factor (MITF)/tyrosinase-related protein (TRP)/tyrosinase activation pathways.
MATERIALS AND METHODS
IB complex preparation
Indian gooseberry extract powder (I) and barley sprout juice powder (B) were provided by HL Science Co., Ltd. (Uiwang, Korea), and the IB complex was composed of I:B = 2:1 (w/w). The extraction methods of I and B are detailed elsewhere [16]. The powders were stored at −20°C until the experiments were conducted.
Cell culture and treatments
The HaCaT (human keratinocytes) cells were provided by Professor Hwang of the College of Life Sciences, Kyung Hee University. Hs27 (human broblasts), and the B16F10 (melanoma) cells were purchased from the American Type Culture Collection (ATCC). Each cell line was cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin, 1% L-glutamine, and 1% sodium pyruvate. Each cell line seeded for each experiment was washed. The HaCaT and Hs27 cells were exposed to 50 mJ/cm2 of UV rays using ve Sankyo Denki G5T5 lamps (Sankyo Denki Co., Yokohama, Japan). The B16F10 cells were differentiated by treating them with 3-isobutyl-1-methylxanthine (IBMX) (250 μM) for 72 h to induce melanin synthesis. All cells stimulated with UVB or IBMX were treated with ascorbic acid (100 μg/mL) or arbutin (100 μg/mL) and the IB complex (50, 100, and 200 μg/mL; the cell viability of HaCaT, Hs27, and B16F10 cells was not affected by the IB complex at those concentrations, data not shown).
Measurement of hyaluronic acid, sphingomyelin, collagenase inhibitory activity, elastase inhibitory activity, pro-inflammatory cytokines, and antioxidant enzyme activity levels
HaCaT cells were lysed, and hyaluronic acid, sphingomyelin, pro-inflammatory cytokines (tumor necrosis factor [TNF]-α, interleukin [IL]-1β, and IL-6), and the antioxidant enzyme activity (superoxide dismutase [SOD], catalase [CAT], and glutathione peroxidase [GPx]) levels were then determined using a Hyaluronic acid ELISA Kit (BioVision Inc., Milpitas, CA, USA), Sphingomyelin Assay Kit (Abcam, Cambridge, UK), TNF-α, IL-1β, and IL-6 DuoSet ELISA kits (all from R&D System, Minneapolis, MN, USA), and SOD, CAT, and GPx activity kits (all from BioVision Inc.), respectively, according to the manufacturer protocols.
Measurement of intracellular melanin and glutathione (GSH) contents
The B16F10 cells were lysed, and the GSH levels were determined using a Glutathione Assay Kit (BioVision Inc.) according to the manufacturer’s protocols.
Measurement of tyrosinase activity, nitric oxide, and cAMP Levels
The B16F10 cells were lysed, and the tyrosinase activity and the nitric oxide (NO) and cAMP levels were then determined using a Tyrosinase Activity Assay Kit (Abcam), Nitric Oxide Assay Kit (Abcam), and cAMP ELISA Kit (Enzo Life Sciences, Plymouth Meeting, PA, USA), respectively, according to the manufacturer protocols.
Isolation of total RNA and reverse transcription-PCR
The total RNA from cells was isolated using a RNeasy Mini kit (Qiagen, Valencia, CA, USA). Complementary DNA synthesis was performed using an iScript™ cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA, USA). PCR amplication consisted of 40 cycles of 95°C for 15 s, 58°C for 15 s, and 72°C for 30 s with a SYBR Green PCR Master Mix (Bio-Rad Laboratories) and primer pairs (Table 1). The data were analyzed using CFX Maestro™ Analysis Software (Bio-Rad Laboratories).
Table 1
Primer sets used for real-time polymerase chain reaction
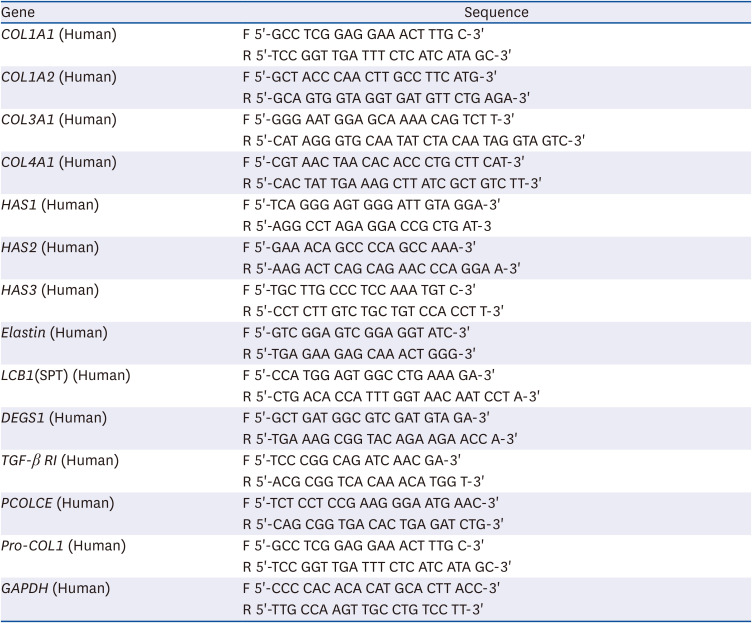
HAS, hyaluronic acid synthase; LCB1 (SPT), long chain base subunit 1 (serine palmitoyltransferase); DEGS1, dihydroceramide desaturase; TGF-β RI; transforming growth factor beta receptor 1, PCOLCE; procollagen C-endopeptidase enhancer, Pro-COL1; procollagen type I; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Protein extraction and Western blot analysis
The cells were lysed using a CelLytic™ MT cell lysis reagent (Sigma, St. Louis, MO, USA). The protein samples (100 µg) were separated using a 10% Mini-PROTEAN® TGX™ Precast Protein Gel (Bio-Rad Laboratories) and electrotransferred onto polyvinylidene diuoride (PVDF) membranes (Bio-Rad Laboratories). The membranes were blocked with 5% skimmed milk in Tris-buffered saline with 0.1% Tween 20 (TBST) for 1 h, and incubated for 12 h at 4°C with antibodies against CerS4 (LASS4), p-IκB, IκB, p65, p-p65, COX2, JNK, p-JNK, c-Fos, p-c-Fos, c-Jun, p-c-Jun, MMP-1, MMP-2, MMP-3, MMP-9, Smad3, p-Smad3, PKA, p-PKA, CREB, p-CREB, MITF, TRP-1, TRP-2, and β-actin (1:1,000). The antibodies were purchased from Cell Signaling (Beverly, MA, USA), Abcam, and LSbio (Seattle, WA, USA). After incubation with the primary antibody, the membranes were incubated with a secondary antibody (anti-rabbit IgG HRP-linked antibody, 1:5,000; Cell Signaling) for 1 h at room temperature. The protein bands were detected with EzWestLumi Plus (ATTO, Tokyo, Japan), developed using Ez-Capture II (ATTO), and quantied using CS Analyzer 3.0 (ATTO).
Statistical analysis
All results are reported as the mean ± SD. The data were assessed statistically using Duncan’s multiple range tests after one-way analysis of variance (ANOVA) using SPSS PASW Statistic v.23.0 (SPSS Inc., Chicago, IL, USA). The differences were considered statistically significant at P < 0.05.
RESULTS
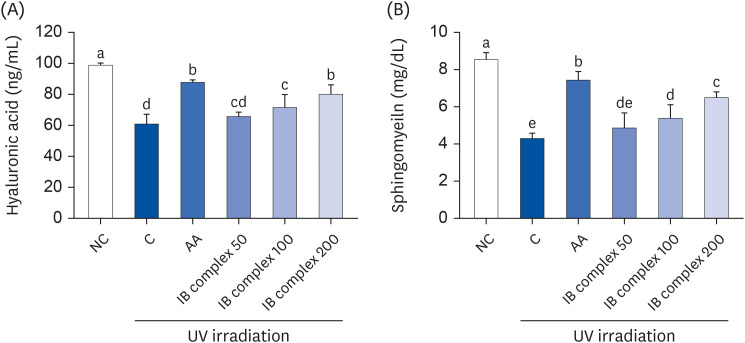
Effects of IB complex on hyaluronic acid, sphingomyelin in UVB-irradiated HaCaT cells, and collagenase inhibitory activity, and elastase inhibitory activity
The effects of the IB complex on hyaluronic acid and sphingomyelin in UVB-irradiated HaCaT cells were analyzed using ELISA methods. The levels of hyaluronic acid were significantly lower in the control group than in the normal control (NC) group, but substantial improvements were observed after the L-ascorbic acid (AA), IB complex 100, and IB complex 200 treatments, showing 44.6%, 17.9%, and 31.7% enhancements, respectively (P < 0.05) (Fig. 1A). The sphingomyelin levels were significantly lower in the control group than the NC group (P < 0.05). The UVB-irradiated HaCaT cells treated with AA, IB complex 100, and IB complex 200 showed significantly higher sphingomyelin levels than the control group (72.7%, 25.0%, and 50.5%, respectively; P < 0.05) (Fig. 1B).
Fig. 1
Effects of the IB complex on hyaluronic acid (A) and Sphingomyelin (B) in UVB-irradiated HaCaT cells. The cells were treated with UVB (50 mJ/cm2) except NC and incubated for 24 h with 100 μg/mL of AA and various concentrations (50, 100, and 200 μg/mL) of the IB complex. The values are presented as means ± SD.
IB, Indian gooseberry and barley sprout; NC, normal control; C, control; AA, L-ascorbic acid.
Different letters show a significant difference at P < 0.05, as determined by Duncan’s multiple range test.
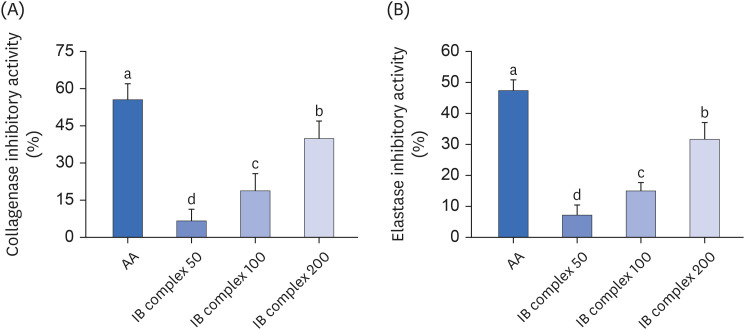
The effects of the IB complex on the collagenase and elastase inhibitory activities were analyzed using the methods reported by Wünsch and Heidrich [17] and Cannell et al. [18]. The collagenase and elastase inhibitory activities were markedly higher in the AA group than in the IB group (55.5% and 47.4%). Furthermore, the IB complexes induced dose-dependent increases in the collagenase and elastase inhibitory activities. The collagenase inhibitory activities in the IB complex 50, IB complex 100, and IB complex 200 were 6.6%, 18.8%, and 39.8%, respectively. The elastase inhibitory activities in the IB complex 50, IB complex 100, and IB complex 200 were 7.1%, 15.0%, and 31.6%, respectively (P < 0.05) (Fig. 2A and B).
Fig. 2
Effects of the IB complex on the collagenase inhibitory activity (A) and elastase inhibitory activity (B). A 100 μg/mL of AA and various concentrations (50, 100, and 200 μg/mL) of the IB complex. The values are presented as the means ± SD.
IB, Indian gooseberry and barley sprout; AA, L-ascorbic acid.
Different letters show a significant difference at P < 0.05, as determined by Duncan’s multiple range test.
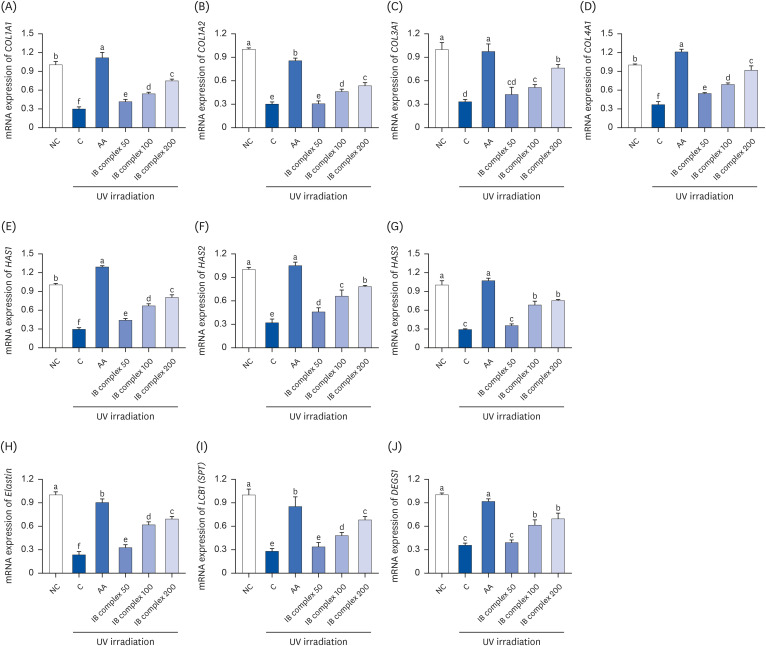
Effects of IB complex on mRNA expression of the factors related to collagen synthesis and skin moisturizing in UVB-irradiated HaCaT cells
The UVB-irradiated HaCaT cells were analyzed using real-time PCR to investigate the collagen synthesis and skin moisturizing-related factors affected by the IB complex. In the control group, the mRNA expressions of COL1A1, COL1A2, COL3A1, COL4A1, hyaluronic acid synthase (HAS) 1, HAS2, HAS3, elastin, long chain base subunit 1 (LCB1) (serine palmitoyltransferase; SPT), and delta 4-desaturase sphingolipid 1 (DEGS1) were significantly lower than the NC group. In particular, the AA group showed significant increases in these expressions (P < 0.05) (Fig. 3). The mRNA expression levels of COL1A1, COL1A2, COL3A1, and COL4A1 were higher in the IB complex 100 and IB complex 200 groups than in the control group (81.0% and 150.7%, 52.5% and 76.8%, 54.3% and 128.4%, and 87.3% and 148.2%, respectively; P < 0.05) (Fig. 3A-D). In addition, the mRNA expression levels of HAS1, HAS2, and HAS3 were significantly higher in the IB complex 100 and IB complex 200 than in the control group (128.5% and 174.8%, 105.7% and 144.0%, and 135.4% and 159.6%, respectively; P < 0.05) (Fig. 3E-G). The mRNA expression of elastin was improved markedly in the IB complex-treated groups than in the control group (39.7–194.1%, P < 0.05) (Fig. 3H). Moreover, LCB1 (SPT) and DEGS1 expression levels were significantly higher in the IB complex 100 and IB complex 200 groups (72.2% and 144.5%, 71.8% and 94.5%, respectively) than in the control group (P < 0.05) (Fig. 3I and J). Therefore, the IB complex directly activated collagen synthesis and moisturizing factors in keratinocytes.
Fig. 3
Effects of the IB complex on COL1A1 (A), COL1A2 (B), COL3A1 (C), COL4A1 (D), HAS1 (E), HAS2 (F), HAS3 (G), Elastin (H), LCB1 (I), and DEGS1 (J) in UVB-irradiated HaCaT cells. The cells were treated with UVB (50 mJ/cm2) except NC and incubated for 24 h with 100 μg/mL of AA and various concentrations (50, 100, and 200 μg/mL) of the IB complex. The values are presented as means ± SD.
IB, Indian gooseberry and barley sprout; HAS, hyaluronic acid synthase; LCB1 (SPT), long chain base subunit 1 (serine palmitoyltransferase); DEGS1, desaturase sphingolipid 1; NC, normal control; C, control; AA, L-ascorbic acid.
The different letters show a significant difference at P < 0.05, as determined by Duncan’s multiple range test.
Effects of IB complex on inflammatory cytokines (TNF-α, IL-1β, IL-6), antioxidant activities (SOD, CAT, GPx), and protein expression of skin moisturizing-related factors in UVB-irradiated HaCaT cells
The effects of the IB complex on the inflammatory cytokines levels and antioxidant activities in UVB-irradiated HaCaT cells were examined using an ELISA kit. The levels of TNF-α, IL-1β, and IL-6 showed a significant increase in the control group compared to the NC group, while the AA group showed a significant decrease compared to the control group (P < 0.05) (Fig. 4A-C). The IB complex 100 showed significantly lower TNF-α and IL-6 levels than the control (20.8% and 23.4%, respectively; P < 0.05) (Fig. 4A-C). Moreover, the IB complex 200 group showed a significant decrease in the TNF-α, IL-1β, and IL-6 levels compared to the control group (35.9%, 31.8%, and 36.0%, respectively; P < 0.05) (Fig. 4A-C). The activities of the antioxidant enzymes (SOD, CAT, and GPx) were significantly lower in the control group than in the NC group. In contrast, the AA group showed significant improvement compared to the control group (P < 0.05) (Fig. 4D-F). Furthermore, the IB complex 100 showed significantly higher SOD and GPx activities than the control (78.4% and 88.1%, respectively; P < 0.05) (Fig. 4D-F), and the IB complex 200 showed remarkably higher SOD, CAT, and GPx activities than the control (118.9%, 58.4%, and 200.8%, respectively; P < 0.05) (Fig. 4D-F).
Fig. 4
Effects of the IB complex on TNF-α (A), IL-1β (B), IL-6 (C), SOD activity (D), CAT activity (E), GPx activity (F), western blot band (G), CerS4 (H), p-IκBα (I), p-p65 (J), MMP-1 (K), MMP-2 (L), MMP-9 (M), and COX-2 (N) in UVB-irradiated HaCaT cells. The cells were treated with UVB (50 mJ/cm2) except NC and incubated for 24 h with 100 μg/mL of AA and various concentrations (50, 100, and 200 μg/mL) of IB complex. The values are presented as means ± SD.
IB, Indian gooseberry and barley sprout; TNF, tumor necrosis factor; IL, interleukin; SOD, superoxide dismutase; CAT, catalase; GPx, glutathione peroxidase; MMP, matrix metalloproteinase; COX-2, cyclooxygenase-2; NC, normal control; C, control; AA, L-ascorbic acid.
Different letters show a significant difference at P < 0.05, as determined by Duncan’s multiple range test.
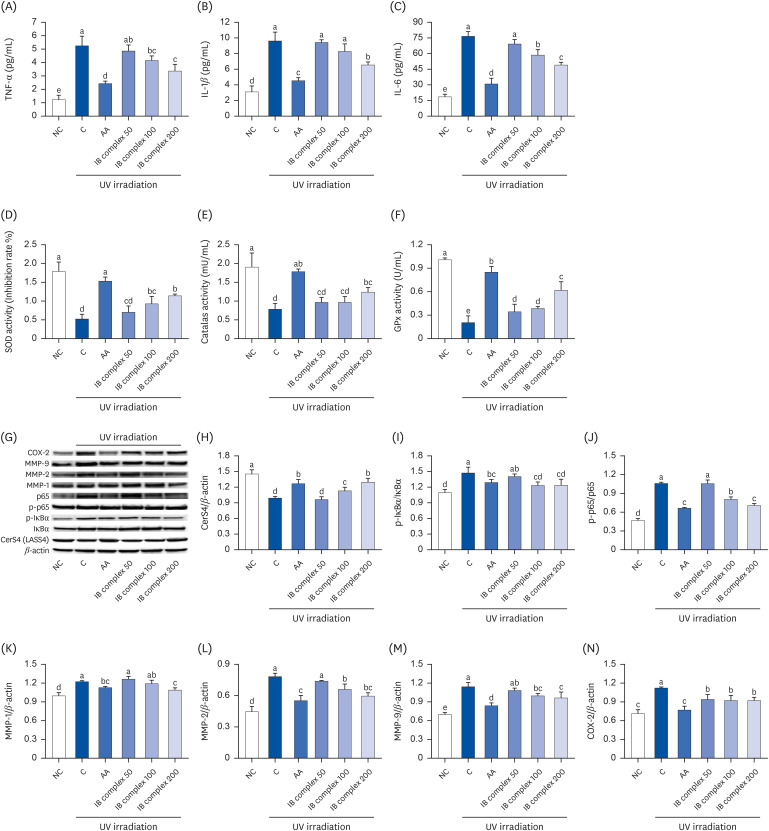
The effects of the IB complex on the protein expression of skin moisturizing-related factors in UVB-irradiated HaCaT cells were examined using Western blot analysis. The protein expression of CerS4 was significantly lower in the control group than in the NC group; however, the protein expressions of p-IκB/IκB, p-p65/p65, MMP-1, MMP-2, MMP-9, and cyclooxygenase-2 (COX-2) were significantly higher in the control group than in the NC group (P < 0.05) (Fig. 4G-N). By contrast, the AA group exhibited a significant increase in the protein expression of Cers4 and a decrease in the protein expression of p-IκB/IκB, p-p65/p65, MMP-1, MMP-2, MMP-9, and COX-2 compared to the control group (P < 0.05) (Fig. 4G-N). The protein expression of CerS4 was significantly higher in the IB complex 100 and IB complex 200 than in the control group (14.0% and 30.9%, respectively; P < 0.05) (Fig. 4G). The cells treated with IB complex 100 and IB complex 200 exhibited a significant decrease in the protein expression of p-IκB/IκB, p-p65/p65, MMP-2, MMP-9, and COX-2 compared to the control group (16.1% and 15.8%, 24.2% and 33.2%, 15.5% and 23.7%, 12.7% and 15.4%, and 18.0% and 17.8%, respectively; P < 0.05) (Fig. 4I, J, L-N). MMP-1 protein expression was significantly lower in the IB complex 200 than in the control group (11.0%) (P < 0.05) (Fig. 4K). These findings suggest that the IB complex effectively adjusted the inflammatory cytokines, antioxidant enzyme activities, and protein expression of skin moisturizing-related factors in response to UVB irradiation.
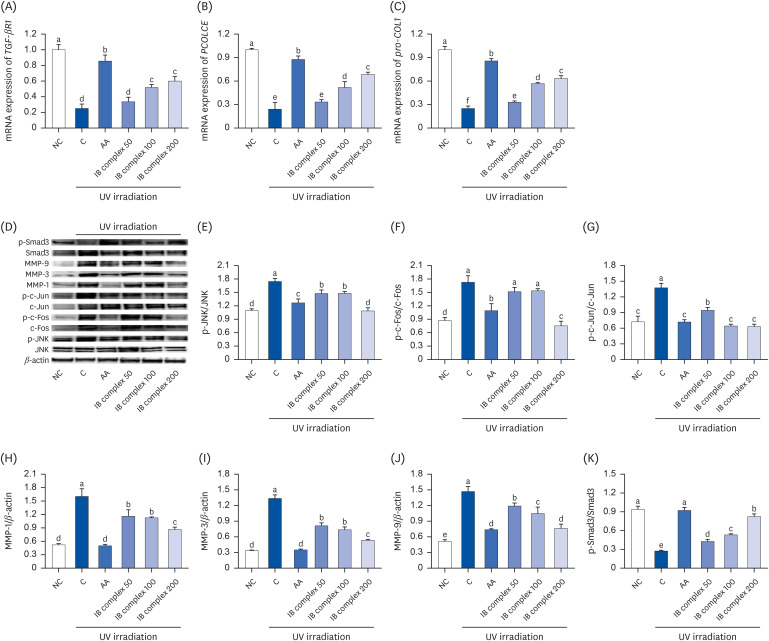
Effects of the IB complex on the skin wrinkle-related factors in UVB-irradiated Hs27 cells
The skin wrinkle-related factors affected by the IB complex in the UVB-irradiated Hs27 cells were analyzed using real-time PCR and Western blot analysis. The mRNA expression of TGF-β RI, procollagen C-endopeptidase enhancer (PCOLCE), and pro-COL1 was significantly lower in the control group than the NC group but higher in the AA group (P < 0.05) (Fig. 5A-C). The UVB-irradiated Hs27 cells treated with IB complex 100 and IB complex 200 showed a significant increase in the mRNA expression of TGF-β RI and PCOLCE compared to the control group (107.8% and 140.9%, 114.9% and 184.7%, respectively; P < 0.05) (Fig. 5A and B). In particular, the mRNA expression of pro-COL1 showed a significant increase in the IB complex-treated group than the control group (32.0–153.2%, P < 0.05) (Fig. 5C). Moreover, the protein expression of p-JNK/JNK, p-c-Fos/c-Fos, p-c-Jun/c-Jun, MMP-1, MMP-3, and MMP-9 showed a significant increase in the control group than the NC group (P < 0.05). The UVB-irradiated Hs27 cells treated with AA showed a significant decrease in the protein expression of p-JNK/JNK, p-c-Fos/c-Fos, p-c-Jun/c-Jun, MMP-1, MMP-3, and MMP-9 (P < 0.05) On the other hand, Smad3 phosphorylation showed a significant decrease in the control groups compared to the NC group (P < 0.05). The UVB-irradiated Hs27 cells treated with AA showed a significant increase in Smad3 phosphorylation (P < 0.05) (Fig. 5D-K). The protein expression levels of p-JNK/JNK, p-c-Jun/c-Jun, MMP-1, MMP-3, and MMP-9 were substantially lower in the IB complex treated groups than in the control group (15.1–37.6%, 31.5–54.3%, 27.4–46.0%, 39.2–60.0%, and 18.8–47.9%, respectively; P < 0.05) (Fig. 5D, E, G-J). The protein expression of p-c-Fos/c-Fos was significantly lower in the IB complex 200 group than in the control groups (56.2%) (P < 0.05) (Fig. 5F). In addition, the UVB-irradiated Hs27 cells treated with IB complex showed a significant increase in Smad3 phosphorylation than the control group (55.3–200.9%) (P < 0.05) (Fig. 5K). Therefore, the IB complex directly controls the wrinkle-related factors in response to UVB irradiation.
Fig. 5
Effects of the IB complex on TGF-βR1 (A), PCOLCE (B), Pro-COL1 (C), western blot band (D), p-JNK (E), p-c-FOS (F), p-c-Jun (G), MMP-1 (H), MMP-3 (I), MMP-9 (J), and p-Smad3 (K), in UVB-irradiated Hs27 cells. The cells were treated with UVB (50 mJ/cm2) except NC and incubated for 24 h with 100 μg/mL of AA and various concentrations (50, 100, and 200 μg/mL) of the IB complex. The values are presented as means ± SD.
IB, Indian gooseberry and barley sprout; TGF-β RI, transforming growth factor-β receptor I; PCOLCE, procollagen C-endopeptidase enhancer; Pro-COL1; procollagen type I; MMP, matrix metalloproteinase; Smad, small mothers against decapentaplegic homolog; NC, normal control; C, control; AA, L-ascorbic acid.
Different letters show a significant difference at P < 0.05, as determined by a Duncan’s multiple range test.
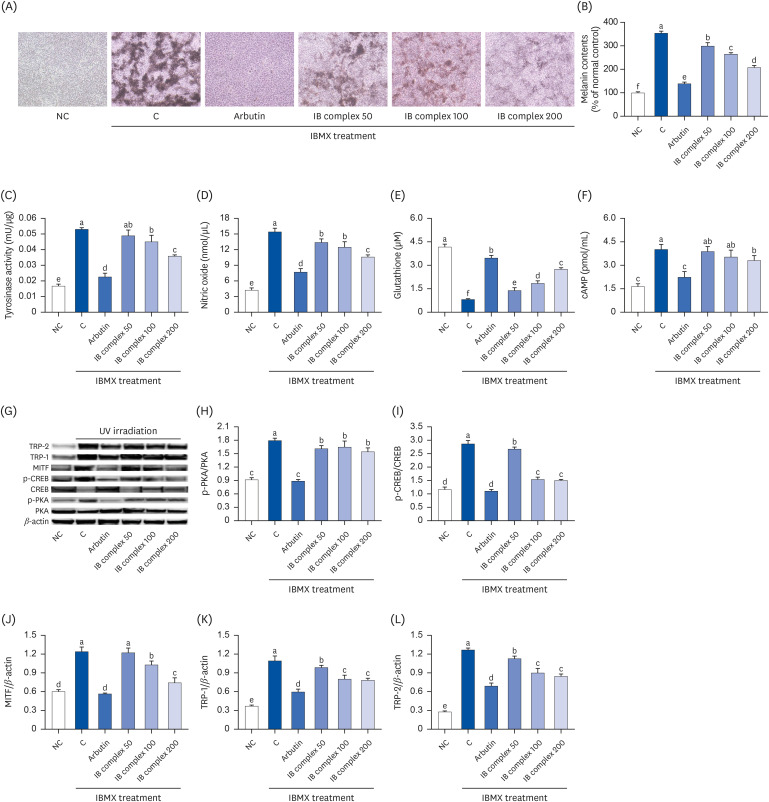
Effects of the IB complex on the melanogenesis-related factors in IBMX-treated B16F10 cells
IBMX-treated B16F10 cells were analyzed using an ELISA kit and Western blot analysis to investigate the effects of the IB complex on the following: melanin content; tyrosinase activity, NO, cAMP, and GSH levels; and melanogenesis-related factors. The melanin content, tyrosinase activity, NO, and cAMP levels were significantly higher in the control group than the other groups, but the arbutin treatment applied to the IBMX-treated B16F10 cells induced a significant decrease in those levels compared to the control group (P < 0.05) (Fig. 6A-D, F). The glutathione levels were significantly lower in the control group than in the NC group, but the arbutin treatment of the IBMX-treated B16F10 cells induced a significant increase in those levels compared to the control group (P < 0.05) (Fig. 6E). The addition of the IB complex to IBMX-treated B16F10 cells with IB complex induced a dose-dependent decrease in the melanin contents compared to the control group (15.6–41.4%; P < 0.05) (Fig. 6A and B). The tyrosinase activities were significantly lower in the IB complex 100 and IB complex 200 than in the control group (15.0% and 32.4%, respectively; P < 0.05) (Fig. 6C). The NO levels were decreased significantly by the addition of the IB complex to the IBMX-treated B16F10 cells compared to the control group (13.0–31.3%; P < 0.05) (Fig. 6D). The addition of the IB complex to the IBMX-treated B16F10 cells showed a dose-dependent increase in the glutathione levels than the control group (72.0–240.3%; P < 0.05) (Fig. 6E), and the cAMP levels were significantly lower in the IB complex 200 group than in the control group (17.5%) (P < 0.05) (Fig. 6F). The protein expressions of p-PKA/PKA, p-CREB/CREB, MITF, TRP-1, and TRP-2 were significantly higher in the control group than in the NC group, but the arbutin treatment of IBMX-treated B16F10 cells induced a significant decrease in p-PKA/PKA, p-CREB/CREB, MITF, TRP-1, and TRP-2 than in the control group (P < 0.05) (Fig. 6G-L). The protein expression of p-PKA/PKA, p-CREB/CREB, TRP-1, and TRP-2 in the IBMX-treated B16F10 cells exhibited a marked decrease in IB complex treatment groups than the control group (8.2–14.0%, 6.7–47.6%, 9.7–28.5%, and 10.8–33.3%, respectively; P < 0.05) (Fig. 6G-I, K, L), and the protein expression of MITF was significantly decreased in the IB complex 100 and IB complex 200 groups than the control group (17.2% and 40.1%, respectively; P < 0.05) (Fig. 6J). These results suggested that the IB complex directly inhibited melanogenesis in IBMX-treated melanocytes.
Fig. 6
Effects of the IB complex on the skin melanogenesis-related factors, melanin contents (A, B), tyrosinase activity (C), NO (D), Glutathione (E), cAMP (F), western blot band (G), p-PKA (H), p-CREB (I), MITF (J), TRP-1 (K), and TRP-2 (L) in IBMX-irradiated B16F10 cells. The cells were treated with 250 μM IBMX except for NC and incubated for 24 h with 100 μg/mL of arbutin and various concentrations (50, 100, and 200 μg/mL) of the IB complex. The values are presented as the means ± SD.
IB, Indian gooseberry and barley sprout; NO, nitric oxide; cAMP, cyclic adenosine monophosphate; PKA, protein kinase A; CREB, cAMP-responsive binding protein; MITF, melanocyte inducing transcription factor; TRP, tyrosinase-related protein; IBMX, 3-isobutyl-1-methylxanthine; NC, normal control; C, control.
Different letters show a significant difference at P < 0.05 as determined by a Duncan’s multiple range test.
DISCUSSION
Skin aging is caused by specific genes related to the skin structure, repair mechanisms, sensitivity to environmental stressors, decreased cell turnover, impaired barrier function, and decreased expression of proteins, such as collagen and elastin, which play essential roles in maintaining skin elasticity and elasticity. Among the environmental factors, UVB and UVA cause degradation of the ECM components, such as hyaluronic acid and proteoglycan, damage to the cell structure due to chronic inflammation and oxidative stress, DNA damage, and promote collagen degradation. UVB irradiation causes oxidative stress in the cellular components, including lipids, proteins, and DNA. Their cumulative impact on the skin-aging process activates MMPs, contributing to collagen and elastin breakdown. This decreases the skin elasticity and causes wrinkle formation. In addition, UVB irradiation increases melanin synthesis [192021]. The present study examined whether the IB complex could inhibit the skin damage caused by UVB-irradiated photoaging and oxidative stress through its effects on skin hydration, wrinkles, and melanogenesis.
HaCaT, Hs27, and B16F10 cells are the commonly used cell lines in skin health-related experiments. HaCaT cells are a human keratinocyte cell line that represents the epidermal cells of the skin. These cells are used widely to study the skin responses to external stimuli such as UV radiation or chemicals. Hs27 cells are a human fibroblast cell line representing the dermal cells of the skin. These cells are used to examine skin cell interactions and skin tissue regeneration. B16F10 cells are a mouse melanoma cell line used frequently in research on melanin production and skin whitening. These cells have a high melanin production capacity, making them a suitable model for evaluating skin whitening effects [222324]. Each cell line represents different skin cell types, allowing researchers to study the skin physiology and pathology.
Maintaining optimal skin hydration is imperative for preserving the epidermal barrier function and overall skin health. This intricate process is orchestrated by the expression and regulation of various genes, each playing a crucial role in the intricate network governing skin hydration. Among these pivotal genes are collagen and elastin, which contribute to the structural integrity of the skin. In addition, HAS1-3 is involved in synthesizing hyaluronic acid, a key component in skin hydration. Furthermore, LCB1 and DEGS1 play essential roles in sphingolipid metabolism, helping to maintain the skin barrier. Sphingolipids are crucial components of the stratum corneum, the outermost layer of the epidermis, and are integral for preventing excessive water loss from the skin [2526]. The degradation of the skin barrier is a multifaceted process influenced by various molecular factors. Among them, oxidative stress, nuclear factor kappa B (NF-κB), and COX-2 play crucial roles. Oxidative stress, characterized by an imbalance between ROS production and antioxidant defenses, contributes significantly to skin barrier dysfunction. The NF-κB, a transcription factor involved in inflammatory responses, has been implicated in regulating the genes associated with immune and inflammatory reactions. Its activation is often observed in the context of skin barrier degradation, linking inflammation to the impairment of skin integrity [272829]. These findings suggest that the IB complex mediates skin hydration by upregulating hyaluronic acid, sphingomyelin, collagen, HAS 1-3, elastin, LCB1, and DEGS1, and suppressing the NF-κB pathways and COX-2 in HaCaT cells.
The formation of skin wrinkles is intricately linked to the degradation of the ECM proteins, with MMP secretion and collagen fragmentation playing pivotal roles in this process. MMPs, a family of enzymes, are key players in ECM remodeling, and their increased activity is associated with the breakdown of structural components, particularly collagen. Collagen, a major protein in the ECM, provides the skin with strength and elasticity. The secretion of MMPs, such as MMP-1 (collagenase), MMP-2 (gelatinase), and MMP-3 (stromelysin), leads to the proteolytic cleavage of collagen fibers. This collagen fragmentation deteriorates the structural integrity of the skin, compromising its ability to resist deformation and maintain a smooth appearance [213031]. The TGF-β/Smad pathway emerges as a pivotal regulatory mechanism governing collagen synthesis, playing a central role in maintaining skin homeostasis. In particular, previous studies have elucidated the intricate interplay between UV irradiation and the modulation of collagen metabolism. UV irradiation has a dual effect on the skin, orchestrating a complex molecular response. One facet of this response involves the up-regulation of MMPs through the mitogen-activated protein kinase (MAPK) pathway. MMPs, such as MMP-1, MMP-2, and MMP-9, are enzymes responsible for degrading various extracellular matrix components, including collagen. The activation of the MAPK pathway by UV irradiation helps increase the expression of MMPs, promoting the breakdown of collagen fibers. On the other hand, the TGF-β/Smad pathway, known for its role in collagen synthesis and fibrogenesis, is down-regulated in response to UV irradiation. This down-regulation of the TGF-β/Smad pathway contributes to a decrease in collagen expression, further exacerbating the imbalance in collagen homeostasis induced by UV exposure [323334]. These results showed that the IB complex could protect against wrinkle formation by enhancing the TGF-β/Smad3 pathway and inhibiting the JNK/c-FOS/c-Jun/MMP pathway activation in Hs27 cells.
The induction of melanogenesis is closely associated with exposure to UV irradiation. Melanogenesis is the process by which melanocytes, specialized pigment-producing cells in the skin, synthesize and distribute melanin, the pigment responsible for skin color. UV irradiation, particularly in the form of UVB and UVA rays, is a potent stimulus for initiating and regulating melanin production. Upon exposure to UV radiation, melanocytes within the skin undergo a series of complex molecular responses. UVB radiation, in particular, directly activates melanocytes by triggering the DNA damage response, leading to the upregulation of various melanogenic enzymes, including tyrosinase. Tyrosinase is a key enzyme in the melanin synthesis pathway, catalyzing the conversion of tyrosine to melanin precursors, ultimately contributing to melanin production. In addition to direct effects on melanocytes, UV irradiation induces the release of signaling molecules, such as α-melanocyte-stimulating hormone (α-MSH), which further stimulates melanogenesis. This hormone binds to the melanocortin receptors on melanocytes, activating the cAMP signaling pathway and promoting the expression of melanogenic genes [353637]. The present study indicated that the IB complex protected melanogenesis by glutathione synthesis, inhibiting tyrosinase activity and the cAMP/CREB/MITF pathway in B16F10 cells.
This study revealed the protective effects of the IB complex against UVB irradiation-induced skin dryness, oxidative stress, wrinkles, and melanogenesis in cellular models. The IB complex uses a multifaceted approach to enhance skin health, promoting increased skin hydration by synthesizing hyaluronic acid and ceramide. Furthermore, it effectively inhibits wrinkle formation by modulating the JNK/c-FOS/c-Jun/MMPs pathway and reduces melanogenesis by down-regulating the PKA/CREB/MITF/TRP-1/TRP-2 pathway in response to UVB irradiation-induced oxidative stress. These findings suggest that supplementation with the IB complex holds significant promise as a preventive measure against skin photoaging. By targeting the key molecular pathways involved in skin hydration, wrinkle formation, and melanogenesis, the IB complex is a valuable candidate for maintaining skin health and combating the adverse effects of UVB irradiation on the skin.
 XML Download
XML Download