

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Obesity is caused by various factors, and mostly arises from excessive energy accumulation and imbalance owing to an excessive calorie intake and insufficient energy consumption. Excess energy is stored in adipose tissue as triglycerides (TGs) [12]. There are two types of adipose tissue: white adipose tissue (WAT) and brown adipose tissue (BAT), with the former accounting for the major proportion. WAT stores energy and secretes hormones such as adiponectin and leptin, whereas BAT is responsible for thermogenesis [34]. The energy-storing function of WAT requires its expansion in the presence of excess residual energy, leading to adipocyte enlargement (hypertrophy) and increased adipocyte numbers (hyperplasia or adipogenesis) [5]. These conditions induce endoplasmic reticulum stress and mitochondrial dysfunction in the adipocytes [6]. Further, loss of normal adipocyte function leads to adipocyte secretion failure, which underlies insulin resistance and inflammatory responses to ultimately increase the risk of obesity-related complications [7].
Saxenda®, an analog of glucagon-like peptide 1, can be used as an adjunct to control body weight (BW) during diet control and continuous exercise; however, its reported side effects include nausea, hypoglycemia, diarrhea, constipation, vomiting, headache, and dyspepsia [89]. Lorcaserin regulates serotonin receptors that elicit satiety by exciting the regulatory neurons of anorexic reactions; however, it causes side effects such as headache, dizziness, fatigue, nausea, thirst, and constipation [10]. Thus, active ongoing investigations are focused on materials or foods with few side effects or those that can exert independent anti-obesity effects. Natural products demonstrate lower disease treatment effects than those of pharmaceuticals; however, they contain active ingredients such as polyphenols which can produce a wide range of pharmacological effects while posing a low risk of side effects.
In addition to dietary control, exercise, and drug treatments, functional health foods have been scientifically verified to prevent and alleviate obesity in vitro and in vivo and have fewer side effects than those of drugs. These are called health functional foods in South Korea, dietary supplements in the U.S., health functional foods in Japan, and food supplements in Europe, and are defined and classified differently in each country. The local health functional food market in South Korea reached KRW 3.33 trillion in 2020, of which individually authorized products, comprising 19.7% of the market, accounted for KRW 653.4 billion [11]. Individually authorized products for body fat reduction are worth KRW 156.7 billion and include Garcinia cambogia extract, Pu-ehr tea extract, and Cissus extract. The effects of these individual authorized ingredients for body fat reduction on adipocyte and adipose differentiation, lipid storage, and fat synthesis have been verified in vitro. Their effects on body fat mass, weight gain, and adipose tissue mass have been confirmed in vivo using high-fat diet (HFD)-induced obese animal models. These compounds have been acknowledged and individually authorized as ingredients that help reduce body fat.
Rosa multiflora, a plant belonging to the Rosaceae family, is commonly found in mountains, valleys, and the edges of fields. In oriental medicine, this plant is used to treat arthritis, edema, and blood stasis; it has also been used in home remedies to regulate digestion, suppress hemorrhage, reduce fever, and to treat beriberi and dropsy. R. multiflora contains flavonoids, such as quercetin, kaempferol glycosides, and tannins like methyl gallate and lycopene [1213]. This plant has been investigated for its antioxidant [1415] and anti-inflammatory effects [16], serum lipid improvement [17], anti-microbial activity [1819], and cosmetic uses; however, only a few studies have reported its anti-obesity effects. In this study, we aimed to determine the effects of R. multiflora root extract (KWFD-H01) on adipogenesis, lipogenesis, and fat metabolism in 3T3-L1 adipocytes and Sprague-Dawley (SD) rats.
MATERIALS AND METHODS
R. multiflora root extract
Wild R. multiflora roots collected from South Korea were extracted at 60°C for 5 h in 70% ethanol (10.7-fold dry w/v) and concentrated using a 1 µm filter and 20 brix. KWFD-H01 concentrate was spray-dried at inlet and outlet temperatures of 170°C and 100 ± 5°C, respectively.
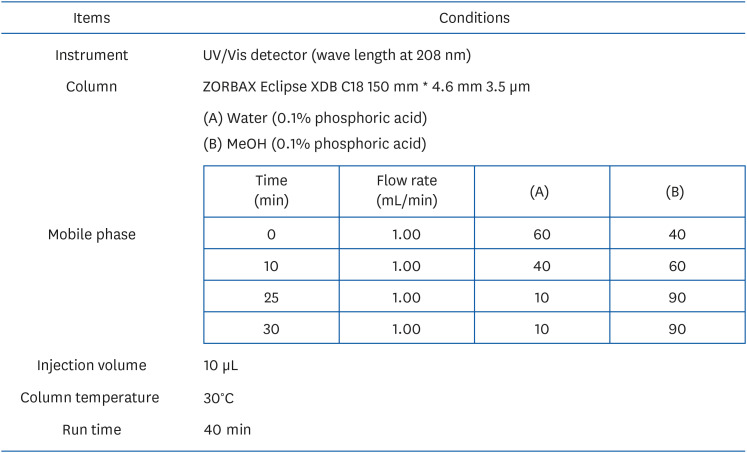
Euscaphic acid high-performance liquid chromatography (HPLC) detection
We analyzed the contents of euscaphic acid in KWFD-H01 under the conditions described in Fig. 1. Standard (1 mg/mL) was dissolved in methanol. KWFD-H01 was dissolved in mobile phase solution (Fig. 1) and concentrated to 10 mg/mL. The HPLC analysis method was validated in terms of specificity, linearity, accuracy, precision. The specificity was determined by analysis of the chromatograms of the standard and sample solutions. The linearity between peak area and concentration was analyzed using three calibration curves obtained with standard solutions of rosamultin at five different concentrations. The accuracy was evaluated by means of recovery tests conducted by adding known amounts to the sample at three different levels. The precision was carried out using five independent tests, including LOD and LOQ. The LOD of the method was calculated for rosamultin as 3.3 times the SD, and LOQ as 10 times the SD.
Cell culture and differentiation
We cultured 3T3-L1 preadipocytes (American Type Culture Collection; ATCC, Manassas, VA, USA) in Dulbecco Modified Eagle Medium containing 10% bovine calf serum (BCS) and 1% penicillin-streptomycin (all from Welgene, Daegu, Korea) and further cultivated at 37°C under a 5% CO2 atmosphere.
We seeded 3T3-L1 preadipocytes (5 × 105/well) into 6-well plates and incubated them for 2 days followed by 2 days in Dulbecco’s modified eagle’s media containing 0.5 mM 3-isobutyl-1-methylxanthine, 0.5 μM dexamethasone, 10 μg/mL insulin (MDI) and 10% fetal bovine serum (FBS). When the cells started to differentiate, the medium was changed and the cells were incubated for 2 days in DMEM containing 10 μg/mL insulin and 10% FBS. Differentiation into adipocytes that formed lipids via cellular fat storage was completed after culture for 4 days in DMEM with 10% FBS.
Cell viability assays
The toxicity effect of KWFD-H01 on non-induced and induced to differentiation 3T3-L1 was assessed using 3-[4,5-dimethylthiazole-2-yl]-2,5- di-phenyl-tetrazolium bromide (MTT) reduction. Undifferentiated 3T3-L1 cells (1 × 104 cell/well) were seeded in 96-well plates and prepared non-induced and induced to differentiate cells. The reagent diluted in culture medium without BCS was added to the wells and the cells were further incubated for 24 h. The medium was replaced with MTT (0.5 mg/mL; 100 μL/well) in the culture medium and incubated at 37°C for 4 h. The culture medium was discarded and dimethyl sulfoxide (100 μL/well) was added to the formazan product that was dissolved by shaking for 30 min. The absorbance of the samples was measured at 570 nm using a UV/vis Opiwen 2120UV plus spectrophotometer (Mecasys Co. Ltd., Daejeon, Korea). Cytotoxicity was then determined by comparison with absorbance of controls.
Adipocyte differentiation and oil red O staining
For the differentiation into adipocytes, 3T3-L1 preadipocytes (5 × 105 cell/well) were aliquoted to a 6 well plate and cultured up to the highest cell density. After a 2 days culture, the cells were transferred to DMEM containing MDI solution and 10% FBS for another 2 days culture to initiate the differentiation; then, the media was replaced with DMEM containing 10 μg/mL insulin and 10% FBS for another 2 days culture to promote differentiation. Next, the cells were cultured in DMEM containing 10% FBS for 4 days to complete the differentiation process into adipocytes that exhibit lipid droplets due to intracellular lipid accumulation.
To investigate the effect of KWFD-H01 on lipid accumulation during adipocyte differentiation induction, the cells were treated with MDI solution and KWFD-H01 each time the media was replaced at 0.05 mg/mL, 0.1 mg/mL, and 0.2 mg/mL concentrations; then, the cells were cultured until differentiation induction was completed. Afterward, the cells were washed twice with phosphate buffered saline (PBS) and fixed using 10% formalin at 4°C for 1 h, followed by additional washing and treatment with 60% isopropanol solution to stain for adipocytes. The stained cells were washed with PBS and after eluding oil red O using 100% isopropanol, the absorbance was measured at 520 nm. The lipid accumulation was verified through comparison with the control.
Isolation of total RNA and quantitative real-time polymerase chain reaction (PCR)
There is confirmation that KWFD-H01 has an effect on adipogenesis and lipogenesis related gene expression in 3T3-L1 cells. Differentiated 3T3-L1 adipocytes were incubated for 24 with KWFD-H01 concentrates (0.05, 0.1, and 0.2 mg/mL) h. Total RNA was isolated from the cells using PURY RNA Plus kits (P2030; GenDEPOT LLC., Katy, TX, USA) as described by the manufacturer. The concentration and purity of the isolated RNA was measured at A260/A280, using a NanoDrop spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). Complementary DNA was synthesized using 1 μg RNA and Reverse Transcription Master Premix (Elpis-Biotech, Daejeon, Korea). Real-time PCR of the synthesized cDNA proceeded using a LightCycler 480 Instrument II (Roche, Mannheim, Germany) and the primers used shown in Table 1. The PCR mix comprised a final volume of 10 μL containing 0.5 μL of each primer, 5 μL SYBR Mixture (Roche), 1 μL cDNA, and 3 μL RNase-free water and the cycling conditions were: 95°C for 5 min followed by 95°C for 15 s, 58°C for 15 s, and 72°C for 30 s. Relative gene expression levels were normalized to that of Gapdh, and fold changes in gene expression were calculated in comparison with the reference gene Gapdh.
Table 1
Real-time polymerase chain reaction primer sequence
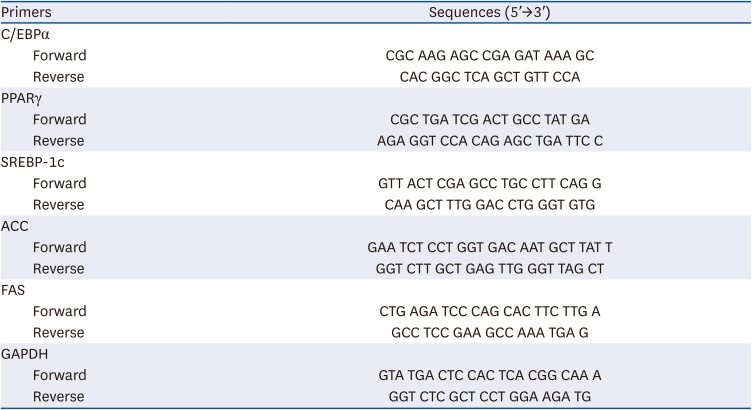
C/EBPα, cytidine-cytidine-adenosine-adenosine-thymidine (CCAAT)/enhancer binding proteins; PPARγ, peroxisome proliferator-activated receptor γ; SREBP-1c, sterol regulatory element-binding transcription factor 1; ACC, acetyl-CoA carboxylase; FAS, fatty acid synthase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Western blot analysis on 3T3-L1 adipocytes
There is confirmation that KWFD-H01 has an effect on adipogenesis and lipogenesis related protein expression in 3T3-L1 cells. The 3T3-L1 adipocytes after the completion of differentiation were treated with R. multiflora root extract at 0.05 mg/mL, 0.1 mg/mL, and 0.2 mg/mL concentrations for a 24 h culture. The cultured cells were scraped using a cell scraper, then centrifuged at 5,000 rpm for 10 min and lysed using RIPA buffer. From the lysed cells, proteins of equivalent quantity were mixed with sample buffer containing sodium dodecyl sulfate (SDS) and β-mercapto-ethanol at a 3:1 ratio using the Bradford method. The mixture was heated at 100°C for 10 min. The protein samples underwent electrophoresis in SDS-polyacrylamide gel electrophoresis and were transferred to a polyvinylidene fluoride membrane (0.45 μm, PVDF transfer membrane, Thermo, Rockford, IL, USA). The membrane was placed in tris-buffered saline (TBS) containing 0.1% tween 20 and 5% skim milk to block for 2 h. Next, the buffer with the following primary antibodies: peroxisome proliferator-activated receptor γ (PPARγ, 1:1,000), cytidine-cytidine-adenosine-adenosine-thymidine (CCAAT)/enhancer binding proteins (C/EBPα, 1:1,000), and sterol regulatory element-binding transcription factor 1 (SREBP-1c, 1:1,000), pSer79 and total acetyl-CoA carboxylase (pACC and ACC, 1:1,000) and fatty acid synthase (FAS, 1:1,000) (Cell Signaling Technology, Danvers, MA, USA) was applied for an overnight reaction, and then followed by washing with TBS-T (TBS containing 0.1% tween 20) for 5 min, which was repeated three times. Next, the buffer containing horseradish peroxidase-conjugated secondary antibody (1:1,000) (Cell signaling technology) was applied for 1 h, and the enhanced chemiluminescence method was used for X-ray film exposure. The intensity of the detected band was analyzed using ImageJ (National Institute of Health, Bethesda, MD, USA) software.
Animal experiments
To test the anti-obesity effect of KWFD-H01, Four-week-old male SD rats weighing 130 and 150 g (Central Lab Animal Inc., Seoul, Korea) were maintained at room temperature (221 ± 1°C) with 55 ± 3% relative humidity with a 12-h photoperiod (8 a.m./8 p.m.) and given solid feed and drinking water ad libitum for one week of acclimation. Thereafter, a HFD was provided for 4 weeks to induce obesity. Animal models of induced obesity were randomly assigned to four groups (n = 7 per group and bred for 8 weeks while consuming normal diet (ND), HFD, HFD + hydroxycitric acid 200 mg/kg BW, and HFD + KWFD-H01 100 mg/kg BW (KWFD-H01). All procedures and methods for animal experiments proceeded according to the guidelines of the Institutional Animal Care and Use Committee at the Chuncheon Bioindustry Foundation (CBF IACUC No. 2020-018).
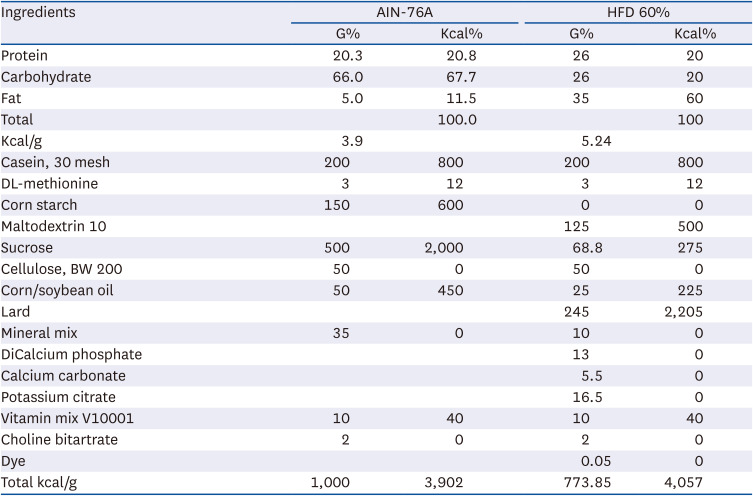
The experiment feed was AIN-76 and HFD 60% (Table 2), and KWFD-H01 was diluted in carboxymethyl cellulose sodium salt and were treated orally (gavage) once daily. The rats were weighed at specific times 2 days per week throughout the experimental period, and feed was removed 1 h before weighing to reduce errors due to weight fluctuations caused by feed intake.
Table 2
The experiment diet of normal group and HFD group
Sample collection
Animal models fasted for 18 h and were anesthetized zoletil (25 mg/kg) and xylazine (5 mg/kg) mixture after completion of the experiment. Blood samples were collected from the hearts of experimental animals and centrifuged at 3,200 × g for 20 min at 4°C. Serum was separated from blood stored at −80°C. The epididymal fat were extracted from animal models, weighed and stored at −80°C.
Plasma biochemical analysis
Serum was evaluated using a Konelab 20B clinical chemistry analyzer (Thermo Fisher Scientific Inc., Vantaa, Finland) to analyze key liver marker enzymes and liver lipid, γ-glutamyl transferase (GGT), aspartate aminotransferase (AST) and alanine aminotransferase (ALT), lactate dehydrogenase (LDH), TG, total cholesterol, and high high-density lipoprotein (HDL), and low-density lipoprotein (LDL) cholesterol. The concentrations of blood serum insulin, leptin, and adiponectin were measured using Rat Insulin (#ERINS, Invitrogen, Waltham, MA, USA), Rat Leptin (ab100773), and Adiponectin (Ab239421; both from Abcam, Cambridge, UK).
Western blot analysis on epididymal adipose tissue
An 1 g of epididymal adipose tissue (blood removed) was mixed with 5 times the RIPA buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1 mM Na2 EDTA, 1 mM EGTA, 1% NP-40, 1% sodium deoxycholate, 2.5 mM sodium pyrophosphate, 1 mM beta-glycerophosphate, 1 mM Na3VO4) 1 g/mL and homogenize the epididymal adipose tissue for 10 min at 4°C using a homogenizer (HS30E, DAIHAN, Seoul, Korea). The supernatant was then recovered by centrifugation at 10,000 g for 10 min. Following protein quantification of the recovered supernatant, the western blot experiment was carried out in the same way as with 3T3-L1 adipocytes.
Statistical analysis
Data are expressed as means ± SD and were analyzed using SAS statistical software (SAS Institute Inc., Cary, NC, USA). One way analysis of variance was performed following Levene’s test, and the significance of the be-tween-group variation was confirmed. A post hoc test was performed depending on the homogeneity of dispersion (Tukey test for multiple comparison in case of homogeneous dispersion; Dunnett’s T3 in the case of heterogeneous dispersion). The confidence interval was set at 95%. All data were expressed as means ± standard error of the mean.
RESULTS
Euscaphic acid in KWFD-H01
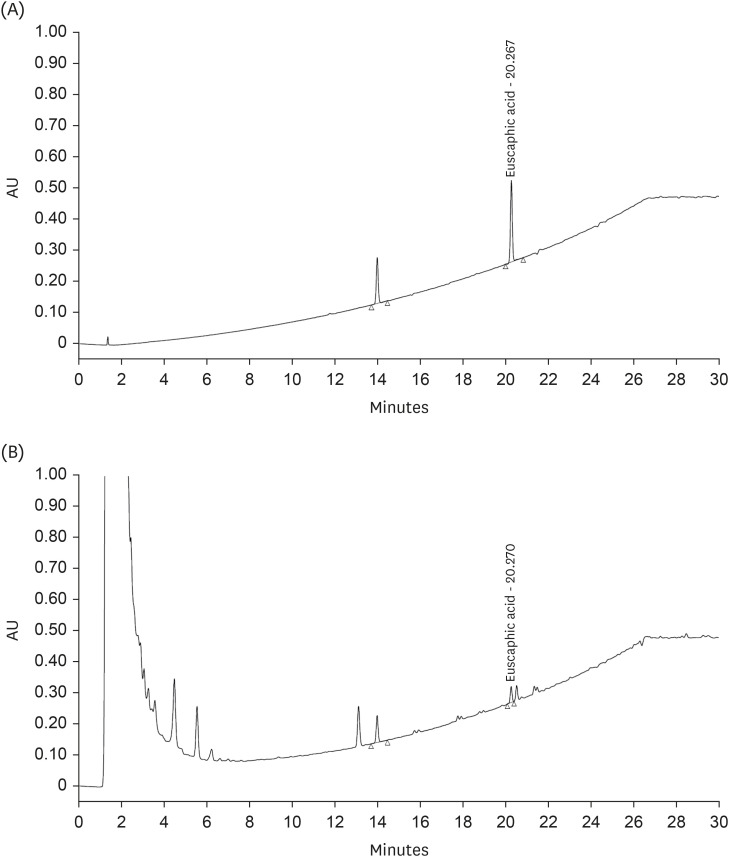
The active substances in KWFD-H01 were determined using HPLC analysis of euscaphic acid, which contains antioxidant and hepatoprotective compounds. The retention time of the euscaphic acid standard was 20.267 min while that of euscaphic acid in KWFD-H01 was 20.270 min. The euscaphic acid content was determined to be 5.97 mg/g (Fig. 2).
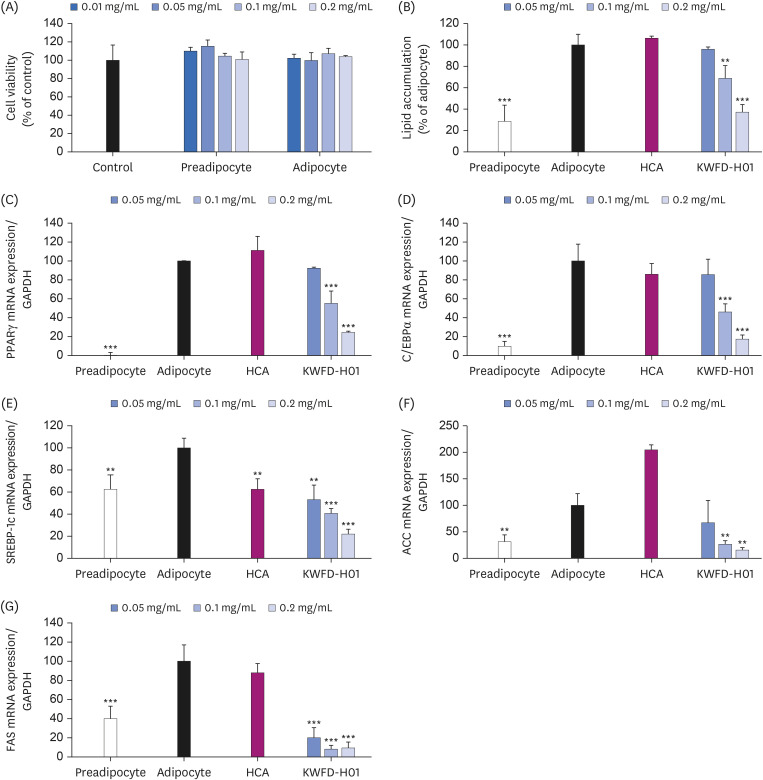
Effect of KWFD-H01 on cell viability and lipid accumulation
The effect of KWFD-H01 on 3T3-L1 preadipocyte and adipocyte viability was confirmed using an MTT assay. The results confirmed that KWFD-H01 did not affect cell viability up to a dosage of 0.2 mg/mL (Fig. 3A). Oil red O staining was performed to analyze the effect of KWFD-H01 on lipid accumulation in 3T3-L1 adipocytes. Upon induction of adipocyte differentiation, 3T3-L1 preadipocytes showed intracellular lipid accumulation (Fig. 3B). Treatment with KWFD-H01 during the induction of differentiation decreased the lipid accumulation in a concentration-dependent manner. The effect of KWFD-H01 on lipid accumulation in 3T3-L1 adipocytes was significantly decreased by 6.0%, 35.7%, and 64.6% at 0.05 mg/mL, 0.1 mg/mL, and 0.2 mg/mL, respectively.
Fig. 3
Cell viability, lipid accumulation, and real-time PCR showing the mRNA expression levels in 3T3-L1 cells after treatment with KWFD-H01. Real-time PCR showing the mRNA levels of PPARγ, C/EBPα, SREBP-1c, ACC, and FAS in 3T3-L1 cells after treatment with KWFD-H01. Preadipocyte: undifferentiated 3T3-L1; Adipocyte: differentiated 3T3-L1; HCA: adipocyte treated with 0.2 mg/mL of HCA; KWFD-H01: adipocyte treated with 0.05, 0.1 and 0.1 mg/mL of KWFD-H01.
PCR, polymerase chain reaction; KWFD-H01, Rosa multiflora root extract; HCA, hydroxycitric acid; PPARγ, peroxisome proliferator-activated receptor γ; C/EBPα, cytidine-cytidine-adenosine-adenosine-thymidine (CCAAT)/enhancer binding proteins; SREBP-1c, sterol regulatory element-binding transcription factor 1; ACC, acetyl-CoA carboxylase; FAS, fatty acid synthase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
**P < 0.01 and ***P < 0.001, when compared to the adipocyte group.
Inhibitory effect of KWFD-H01 on adipogenic gene expression
The expression of Pparγ and C/ebpα mRNAs in 3T3-L1 cells was analyzed to determine the effects of KWFD-H01 on adipocyte differentiation-regulating genes. Fig. 2C and D show that the mRNA expression of Pparγ and C/ebpα, which regulate adipocyte differentiation in 3T3-L1 adipocytes, was inhibited by 7.1%, 43.5%, and 75.7%, and 14.4%, 53.9%, and 82.7%, respectively in cells incubated with KWFD-H01 0.05, 0.1, and 0.2 mg/mL compared with that in the control adipocytes.
Effect of KWFD-H01 on lipogenesis-associated gene expression
We examined the mRNA expression of Srebp-1c, Acc, and Fas, which regulate lipid diffusion and TG synthesis in 3T3-L1 cells incubated with 0.05, 0.1, and 0.2 mg/mL of KWFD-H01 to determine the effects of KWFD-H01 on lipogenesis-associated genes (Fig. 3E-G). Srebp-1c mRNA expression in 3T3-L1 adipocytes was inhibited by 46.9%, 59.4%, and 78.1%, respectively, compared with that in control adipocytes. Acc and Fas mRNA expression was inhibited by 47.0%, 83.6%, and 91.3% and by 82.1%, 92.9%, and 91.7%, respectively, compared with that in the control adipocytes.
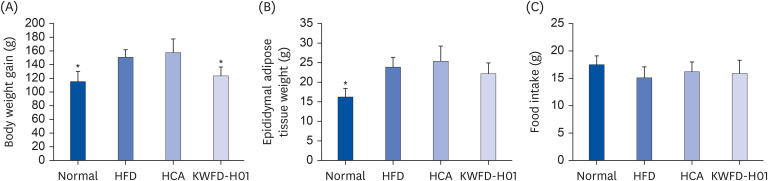
Effects of KWFD-H01 on BW and food intake
Fig. 4 shows the BW changes and weight gain in the rat models of obesity. A significant amount of weight was gained Rats in the HFD group showed significant weight gain compared with that in the ND group during the experimental period, whereas weight gain was reduced in the HFD + KWFD-H01 (100 mg/kg) group compared with that in the HFD group. Overall. weight gain was 130.9% higher in the HFD group than in the ND group and was reduced by 18.0% in the HFD + KWFD-H01 100 mg/kg group. The epididymal adipose tissue weight was 147.1% higher in the HFD group than in the ND group, and was reduced by 7.1% in the HFD + KWFD-H01 100 mg/kg group. However, food intake did not differ among the groups.
Fig. 4
Effects of KWFD-H01 on BW gain, epididymal adipose tissue weight, and food intake in high-fat diet-induced-obese rat for 8 weeks. (A) BW gain. (B) Epididymal adipose tissue weight. (C) Food intake. The data are presented as means ± standard error of the mean.
HFD, high-fat diet; HCA, hydroxycitric acid; KWFD-H01, Rosa multiflora root extract.
*P < 0.05, when compared to the HFD group (n = 7 per group).
Effects of KWFD-H01 on plasma biochemical levels
Table 3 shows the effects of KWFD-H01 on the plasma biochemical levels in the SD rat models. TGs increased by 206.0% in the HFD group compared with those in the ND group. Total and LDL cholesterol levels increased by 129.5% and 120.0%, respectively, whereas HDL cholesterol levels decreased by 38.5%. The plasma levels of TGs, total cholesterol, and LDL-cholesterol were decreased by 47.6%, 33.4%, and 48.7%, respectively, while HDL-cholesterol was increased by 122.9% in the HFD + KWFD-H01 (100 mg/kg) vs. the HFD group.
Table 3
Effects of KWFD-H01 supplementation on serum profile in SD-rat
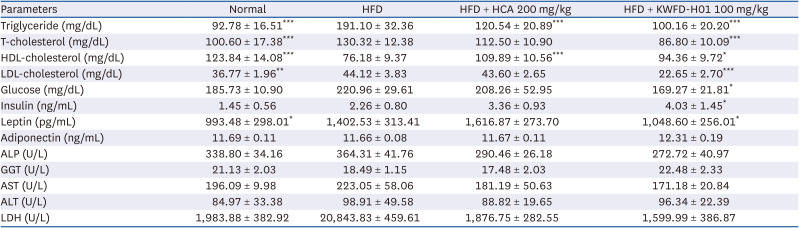
The data are presented as means ± standard error of the mean.
KWFD-H01, Rosa multiflora root extract; SD, Sprague-Dawley; HFD, high-fat diet; HCA, hydroxycitric acid; T-cholesterol, total cholesterol; LDL, low density lipoprotein; HDL, high density lipoprotein; ALP, alkaline phosphatase; GGT, γ-glutamyl transferase; AST, aspartate aminotransferase; ALT, alanine aminotransferase; LDH, lactate dehydrogenase.
*P < 0.05, **P < 0.01, and ***P < 0.001, when compared to the HFD group (n = 7 per group).
Blood glucose and hormone levels were also changed after KWFD-H01 administration. Glucose, insulin, and leptin levels increased by 118.9%, 155.9%, and 141.2%, respectively in the HFD group compared with those in the ND group. In contrast, glucose, insulin, and leptin levels in the HFD + KWFD-H01 100 mg/kg group decreased by 23.3%, 198.3%, and 25.3%, respectively, whereas adiponectin levels increased by 105.6% compared with those in the HFD group. The biochemical indicators of hepatotoxicity, including alkaline phosphatase, GGT, AST, ALT, and LDH did not change in any of the experimental groups.
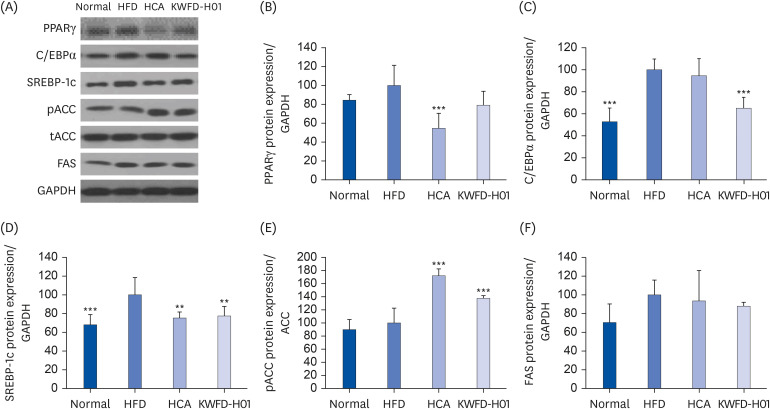
Effects of KWFD-H01 on adipogenesis- and lipogenesis-associated gene expression in rat models of obesity
Protein expression in the epididymal adipose tissues of the rat models was measured to confirm the effects of KWFD-H01 on adipose differentiation- and fat synthesis-regulating gene expression. Fig. 5 shows that the levels of Pparγ and C/ebpα proteins. which control adipose differentiation in epididymal adipose tissues, were significantly reduced in the HFD + KWFD-H01 100 mg/kg group compared with those in the HFD group. The protein levels of Srebp-1c and Fas, which regulate fat synthesis, were reduced, whereas those of phosphorylated Acc were increased in the HFD + KWFD-H01 100 mg/kg group compared with those in the HFD group.
Fig. 5
Effects of KWFD-H01 on the expression levels of proteins related to adipogenesis/lipogenesis in epididymal adipose tissue. Changes in the adipogenesis/lipogenesis-related protein levels were expressed as the fold-increase relative to their expression in the HFD group after normalization to the GAPDH protein expression level. The data are presented as means ± standard error of the mean.
PPARγ, peroxisome proliferator-activated receptor γ; C/EBPα, cytidine-cytidine-adenosine-adenosine-thymidine (CCAAT)/enhancer binding proteins; SREBP-1c, sterol regulatory element-binding transcription factor 1; pACC, phosphorylated acetyl-CoA carboxylase; tACC, total acetyl-CoA carboxylase; FAS, fatty acid synthase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; HFD, high-fat diet; HCA, hydroxycitric acid; KWFD-H01, Rosa multiflora root extract.
**P < 0.01, and ***P < 0.001, compared to the HFD group.
DISCUSSION
We previously confirmed that KWFD-H01 reduces lipid accumulation and TG levels in 3T3-L1 adipocytes as well as the protein levels of PPARγ and C/EBPα, which induce adipose differentiation. In this study, the anti-obesity effects of KWFD-H01 were confirmed both in vitro and in vivo, qualifying it for registration as an individually authorized functional health food.
Multiflorins A and B, multinoside A, multinoside A acetate, isoquercitrin, hyperin, and heliocioside A have been identified in R. multiflora [1418]. In this study, euscaphic acid was predicted to be the active component of KWFD-H01because other studies have demonstrated that euscaphic acid regulates body fat [20]. The euscaphic acid content of KWFD-H01 was 5.97 mg/g. Euscaphic acid also exerts anti-diabetic, anti-oxidative, anti-inflammatory, and anti-coagulant effects [1418].
KWFD-H01 was found to exert regulatory effects on genes related to adipogenesis and lipogenesis in 3T3-L1 adipocytes. These cells differentiate into adipocytes depending on signal transduction pathways and transcription factors, such as the adipogenesis-regulating genes Pparγ and C/ebpα. Although Pparγ and C/ebpα are master regulator genes of adipocyte differentiation, PPARγ protein is a ligand-activated transcription factor that mediates the expression of adipose-related genes and activates adipogenesis [21]. C/ebpα expression induces adipocyte differentiation and adipogenesis only in the presence of Pparγ [515]. Both Pparγ and C/ebpα can induce the expression of genes related to adipogenesis, such as ap2 and LPL [16]. Lipogenesis is controlled by several proteins including Srebp-1c, Acc, and Fas; Srebp-1c stimulates fatty acid synthesis and nuclear receptor ligands as well as activates Pparγ factors; Acc, and Fas are key enzymes in de novo lipogenesis and regulate substrate production for lipogenesis through acetyl-CoA, malonyl-CoA, and palmitic acid synthesis [121719]. The present study showed that KWFD-H01 inhibited adipogenesis- and lipogenesis-related genes. The reduced lipid accumulation and TG content observed in a previous study can be attributed to the ability of KWFD-H01 to inhibit adipose tissue differentiation and synthesis.
Obesity is caused by an imbalance between energy intake and consumption, and the surplus energy is stored as TGs in fat cells as TGs [13]. We confirmed the anti-obesity effects of KWFD-H01 in HFD-induced obese rats. A HFD easily and concurrently induces hyperlipidemia and obesity as well as increases BW and adipose tissue in SD rats [22]. Weight gain was reduced in rats fed HFD + KWFD-H01 (100 mg/kg) for 8 weeks, without changes in food intake, compared with that in the HFD group. The anti-obesity effects of KWFD-H01 were consistent with those of our previous results. Blood samples from the HFD and KWFD-H01 (100 mg/kg) groups contained significantly less TF, T-cholesterol, and LDL-C, and more HDL-C than those from the HFD group. Overall, KWFD-H01 can help reduce HFD-induced hyperlipidemia and dyslipidemia [23]. The HFD + KWFD-H01 (100 mg/kg) group showed lower blood glucose and higher insulin levels. According to Lu et al. [24], increased liver enzyme activity should be associated with blood glucose reduction and increased insulin secretion from pancreatic β-cells. Intake of KWFD-H01 also resulted in reduced leptin and increased adiponectin levels. Adipose tissue is an endocrine organ that regulates the hormonal system and controls adipokine secretion, including leptin and adiponectin; further, the amount of adipose tissue determines the adipokine secretion levels [25]. Leptin and adiponectin regulate lipolysis in adipocytes through hormone-stimulated insulin sensitivity, glucose uptake, and fatty acid oxidation via hormone-stimulated lipolysis [252627]. In this study, we found that KWFD-H01 reduced the elevated blood glucose and leptin secretion caused by HFD-induced obesity and increased adiponectin secretion, which alleviated or improved obesity.
Adipogenesis- and lipogenesis-related protein levels were measured in the epididymal adipose tissue of HFD-induced obese rats to determine how KWFD-H01 reduced body fat. The levels of adipogenesis-regulating proteins like PPARγ and C/EBPα were decreased consistent with their mRNA expression in 3T3-L1 adipocytes. The levels of SREBP-1c and FAS proteins, which regulate lipogenesis, were also decreased, whereas those of phosphorylated ACC were increased. Notably, KWFD-H01 inhibited adipogenesis and lipogenesis both in cells and in rat models of obesity. The ability of KWFD-H01 to control obesity was attributed to BW and adipocyte reduction via the inhibition of adipogenesis and lipogenesis.
Overall, the present study confirmed the inhibitory effects of KWFD-H01 on adipogenesis and lipogenesis, both in vitro and in vivo. These anti-obesity effects were evident through weight loss and improved blood profiles in rat models. Thus, KWFD-H01can be applied as a functional health food to address obesity. However, future clinical studies examining and verifying the anti-obesity activity and safety of KWFD-H01 are needed before its application as a functional health food.
 XML Download
XML Download