

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Chronic renal failure (CRF) describes the slowly progressing damage to the kidney as a complication of certain primary or secondary chronic kidney diseases, which could disrupt the essential functions of the kidney [1]. The clinical manifestations of CRF include the following: the accumulation of metabolites, toxins, and electrolytes; acid–base balance disorders; a group of syndromes characterized by abnormal endocrine function, which cause symptoms, such as nausea, vomiting, fatigue, and limb swelling; and at the terminal stage, uremia. Its diagnosis is often delayed because the symptoms of CRF are less evident in the early stages, by which time it has already caused severe harm to the patient. Furthermore, the current treatment of CRF is focused mainly on controlling the deterioration of the disease, e.g., via intestinal adsorbents and regulation of the acid–base balance, but they cannot cure the disease or reverse the damage. For end-stage patients, additional measures, such as dialysis or kidney transplantation, are required, which are expensive and have many side effects. The better treatment of CRF requires a more in-depth understanding of its pathogenesis and regulatory mechanisms.
Podocytes are highly differentiated epithelial cells covering the outer surface of glomerular capillaries. The damage to podocytes, which manifests as foot process fusion, cell density reduction, apoptosis, epithelial-mesenchymal transition, and hypertrophy, is an important marker of CRF and a potential target for its treatment [2]. Autophagy is a protective measure against podocyte damage under various pathological conditions and can help inhibit many renal diseases [34]. On the other hand, the effect of autophagy on CRF remains controversial. The mammalian target of rapamycin (mTOR) is an atypical serine/threonine protein kinase whose activity depends on the nutrient status and redox levels [5]. mTOR is a main regulator of cellular growth, proliferation, protein and lipid synthesis, autophagy, and metabolism. Promoting autophagy by inhibiting the mTOR pathway has renoprotective effects in rats with CRF [6]. On the other hand, inhibiting autophagy by activating the mTOR pathway alleviates renal fibrosis and protects the renal function in rats with CRF [7]. Further studies are needed to clarify the exact functions of autophagy in podocytes and their effects on CRF.
Traditional Chinese medicine has shown remarkable effects in treating CRF patients and has been found to alleviate symptoms, protect residual renal function, and delay the progress of the disease in early and intermediate-stage patients [8]. For example, Astragali Radix (Huangqi) is an essential component in the dry roots of Astragalus membranaceus (Fisch.) Bge that reduces the total cholesterol, triglyceride, and low-density lipoprotein levels, increases the plasma protein levels, and improves eGFR in CRF patients [89]. Abelmoschi Corolla (Huangshukui), the flower of Abelmoschus manihot (L.) Medic., is also used widely for CRF in China and has been shown to suppress proteinuria and maintain stable kidney functions in patients [10].
Bee venom (BV) is used to treat inflammatory diseases, such as rheumatoid arthritis [11]. One of the major active components in BV is melittin, a basic peptide consisting of 26 amino acids. Melittin can potentially treat renal diseases, such as renal fibrosis and acute kidney injury [1213], suggesting that melittin may also have therapeutic effects on CRF. Melittin induces autophagy to prevent against rheumatoid arthritis [14] and protozoan pathogens [15]. Melittin also attenuates autophagy to alleviate cerebellar ataxia [16] and has shown strong potential in treating tumors [17]. Nevertheless, the roles of melittin-mediated autophagy in CRF are unclear.
This paper provides in vivo and in vitro evidence that melittin could facilitate autophagy in podocytes and alleviate the podocyte damage caused by CRF. These results shed light on the cellular mechanism of the therapeutic effects of melittin on CRF and also provide compelling evidence supporting the ameliorating role of autophagy in CRF, highlighting it as a promising target for future drug development in the treatment of CRF.
MATERIALS AND METHODS
Animal study
Male Sprague–Dawley rats were purchased from the Zhejiang Academy of Medical Sciences and housed under standard conditions, as described elsewhere [18]. After 1 week of acclimation, the rats were subjected to a 5/6 nephrectomy [19] and divided randomly into the following groups: control group, 5/6-nephrectomized group (5/6 Nx), and melittin groups (n = 6 per group). The melittin groups underwent 5/6 nephrectomy but were then treated with melittin (25, 50, and 100 μg/kg/day; Sigma-Aldrich, St. Louis, MO, USA) for 28 days by intraperitoneal administration. The doses of melittin used in the in vivo experiments were determined according to previous studies [2021]. On days 7, 14, and 28, the rats in each group were inspected for weight, total urine volume within 24 h, urine protein content (C035‐2‐1), serum creatinine (C011‐2‐1), and blood urea nitrogen (C013‐2‐1; all from Nanjing Jiancheng Bioengineering Institute, Nanjing, China). On Day 28, all animals were euthanized, and the remnant kidney tissues were harvested for analysis. All animal procedures followed the National Institutes of Health’s Guide for the Care and Use of Laboratory Animals. The experimental procedures were approved by the Research Ethics Committees of the Shanghai Yangpu District Kongjiang Hospital (LL-2020-KY-24).
Hematoxylin & eosin (H&E), Masson and periodic acid-Schiff (PAS) staining
For H&E staining, the kidney tissues were fixed in a 10% formalin solution, embedded in paraffin, and cut into 4–5 μm thick slices. The slices underwent H&E staining (D006-1; Nanjing Jiancheng Bioengineering Institute). The histopathology of the renal tissues was scored as previously described [22]. For Masson staining, interstitial collagen deposition in the kidney tissues was stained using a Masson staining kit (D026-1; Nanjing Jiancheng Bioengineering Institute). Semiquantitative analysis was performed based on the percentage of collagen area stained in the visual field [23]. For PAS staining, the sections were incubated with 100 μL of a periodic acid solution for 5 min, washed using ddH2O, and incubated with 100 μL of Schiff reagent for 30 min at 37°C in the dark. Semiquantitative scoring of glomerular sclerosis in PAS-stained slides was performed using a 5-grade method described elsewhere [24]. Hematoxylin counterstaining was performed for 2 min.
Cell culture and treatment
Conditionally immortalized mouse podocyte clone cells (MPC5) (Shanghai Enzyme-linked Biotechnology Co., Ltd, Shanghai, China) were cultured in RPMI 1640 medium (HyClone; Cytiva, Marlborough, MA, USA) containing 10% fetal bovine serum, 10 U/mL of interferon-γ (both from Thermo Fisher Scientific, Waltham, MA, USA), and 1% penicillin–streptomycin (Millipore Sigma, Burlington, MA, USA) at 37°C and 5% CO2. In certain experiments, MPC5 cells were treated with 1 μM AngII (Sigma-Aldrich), followed by different concentrations of melittin (2, 4, or 8 mg/L; Selleck, Shanghai, China) in the absence or presence of 10 mM 3-MA (Selleck) for 48 h. The melittin doses used in the in vitro experiments were determined according to previous studies [14252627].
Cell counting kit 8 (CCK‐8) assay
MPC5 cells (104 cells/well) were plated overnight into 96‐well culture plates. Subsequently, 10 µL of a CCK‐8 solution (C0038; Beyotime Biotechnology, Haimen, China) was added to each well, where the cells were incubated at 37°C for 2 h. The absorbance at 450 nm was measured using a DR‐200Bs microplate reader.
RNA isolation and quantitative reverse transcription polymerase chain reaction (RT-PCR)
The total RNA was extracted from kidney tissues using Trizol reagent (Invitrogen, Carlsbad, CA, USA). The mRNA levels of podocin and nephrin were determined by quantitative RT-PCR. The primers used were as follows: podocin, 5′-CTTGGCACATCGATCCCTCA-3′ and 5′-CTCTCCACTTTGATGCCCCA-3′; nephrin, 5′-AACCGAGCCAAGTTCTCCTG-3′ and 5′-GGACGACAAGACGAACCAGT-3′; GAPDH, 5′-GGAGTCTACTGGCGTCTTCAC-3′ and 5′-ATGAGCCCTTCCACGATGC-3′.
Antibodies and western blotting
The primary antibodies used in this study included the following: podocin (Affinity; DF8593), nephrin (Affinity; AF7951), Beclin 1 (Affinity; AF5128), LC3I/II (Affinity; AF5128), phosphorylated mTOR (p-mTOR) (Affinity; AF3308), mTOR (Affinity; AF6308), and GAPDH (Proteintech; 60004-1-Ig). The secondary antibodies used in this study included horseradish peroxidase-conjugated goat anti-rabbit or goat anti-mouse secondary antibodies (Beyotime Biotechnology). The cells were lysed in ice-cold radioimmunoprecipitation assay buffer (50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1% Triton X-100, 0.5% Na-deoxycholate) containing the protease inhibitors. The protein concentration was measured using a BCA protein assay kit (Thermo Fisher Scientific). Equal amounts of cell lysates were analyzed by western blotting using an enhanced chemiluminescence system (Bio-Rad, Richmond, CA, USA).
Data analysis
All experiments were repeated 3 times. All data were analyzed using GraphPad Prism 8.4.2 (GraphPad Software Inc., San Diego, CA, USA) and plotted as the mean ± SD. An analysis of the variance was used to compare the averages of multiple groups. P-values < 0.05 were considered significant.
RESULTS
Melittin ameliorates body weight and renal function in a rat 5/6 Nx
The functional effects of melittin in CRF were examined by establishing a 5/6 Nx of renal failure in rats [19]. The rats were divided into 5/6 Nx group (no treatment) and 3 treatment groups and administered with low (25 μg/kg body weight), intermediate (50 μg/kg body weight), or high (100 μg/kg body weight) dose melittin via an intraperitoneal injection. After 7 days, the rats in the 5/6 Nx group started to show a lower body weight (Fig. 1A) and higher serum creatinine (Fig. 1B), urea nitrogen (Fig. 1C), and urine protein levels (Fig. 1D) than the rats that did not undergo a nephrectomy. These observations persisted over the 4-week observation window, indicating renal damage. The administration of melittin partially restored the body weight of the rats under all 3 doses after 4 weeks (Fig. 1A). Furthermore, it also dose-dependently lowered the serum creatinine, urea nitrogen, and urine protein levels as early as one week after the nephrectomy, the effect of which also persisted throughout the 4-week observation window (Fig. 1B-D).
Fig. 1
Melittin ameliorated the body weight and renal function in a rat 5/6 nephrectomy model. A 5/6 Nx of renal failure was established on rats. The rats were then administered low (25 μg/kg body weight), intermediate (50 μg/kg body weight), or high (100 μg/kg body weight) doses of melittin via intraperitoneal injection. One, 2, and 4 weeks after the administration, serum, urine, and kidney tissues were collected from the rats for assessment. (A) Body weight of each group of rats one, 2, and 4 weeks after administration. (B-D) Serum creatinine (B), urea nitrogen (C), and urine protein level (D) of each group of rats 1, 2, and 4 weeks after the administration.
5/6 Nx, 5/6 nephrectomy model.
***P < 0.001 vs. control; #P < 0.05, ##P < 0.01, ###P < 0.001 vs. 5/6 Nx.
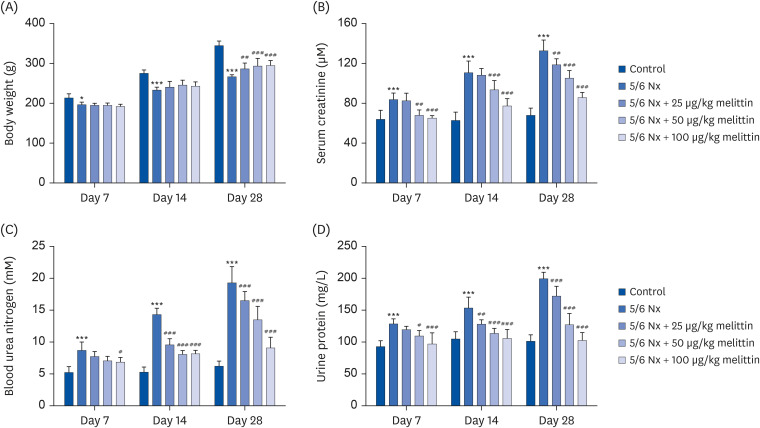
Melittin alleviates renal damage in a rat 5/6 Nx
Kidney tissue sections were acquired 4 weeks after the nephrectomy to observe the effects of melittin. H&E, Masson, and PAS staining were then performed on the tissue samples. Melittin dose-dependently decreased the kidney injury and histopathology scores in 5/6 Nx mice (Fig. 2A and B). Masson staining revealed massive interstitial collagen deposition and significantly increased fibrosis in the 5/6 Nx mice, which was reduced by melittin in a dose-dependent manner (Fig. 2A and C). Hence, melittin reduced renal injury in the 5/6 Nx mouse model. PAS staining showed that the 5/6 Nx mice had the typical characteristics of glomerulosclerosis, including glomerular hypertrophy, capsular adhesion, basement membrane thickening, and pedicle fusion. Melittin dose-dependently reduced the glomerulosclerosis index (Fig. 2A and D).
Fig. 2
Melittin alleviated renal damage in a rat 5/6 Nx. The 55/6 Nx of renal failure was established on rats. The rats were then administered low (25 μg/kg body weight), intermediate (50 μg/kg body weight), or high (100 μg/kg body weight) doses of melittin via intraperitoneal injection. (A) Representative images of H&E, Masson, and PAS staining of kidney sections from mice 4 weeks after the administration. The blue arrows indicate the inflammatory infiltration or swelling of the kidney tissues. The yellow arrows indicate the fibrotic tissues. The green arrows indicate the glomerulosclerosis. Scale bar, 50 μm. (B) Quantification of the kidney injury score in H&E-stained sections. (C) Quantification of the renal interstitial fibrosis score in Masson stained sections. (D) Quantification of the glomerulosclerotic index in PAS-stained sections.
5/6 Nx, 5/6 nephrectomy model; H&E, hematoxylin & eosin; PAS, periodic acid-Schiff.
***P < 0.001 vs. control; #P < 0.05, ##P < 0.01, ###P < 0.001 vs. 5/6 Nx.
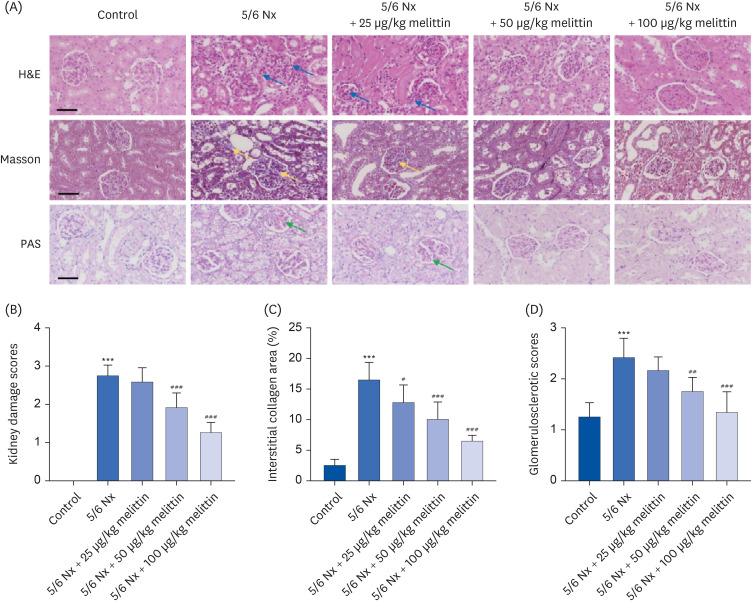
Melittin alleviates podocyte damage and induces autophagy in 5/6 Nx rats
The status of podocytes was examined to understand the mechanism of how melittin alleviated renal damage in 5/6 Nx rats. Podocin and nephrin are 2 proteins that are expressed and can serve as biomarkers of the slit diaphragm, an intercellular junction between podocyte foot processes [28]. Much lower podocin and nephrin mRNA and protein expression levels were detected in the kidney tissues, which were restored by melittin in a dose-dependent manner (Fig. 3A-E).
Fig. 3
Melittin alleviates podocyte damage and autophagy inhibition in 5/6 Nx rats. A 5/6 Nx of renal failure was established on rats. The rats were then administered low (25 μg/kg body weight), intermediate (50 μg/kg body weight), or high (100 μg/kg body weight) doses of melittin via intraperitoneal injection. Four weeks after the administration, kidney tissues were collected from the rats for assessment. (A) Podocin and (B) nephrin expression levels in the kidney tissues were assessed by qRT-PCR. Expression levels of (C, D) podocin, (C, E) nephrin, (C, F) Beclin 1, (C, G) LC3II/LC3I, and (C, H) p-mTOR/mTOR in the kidney tissues assessed by Western blot.
5/6 Nx, 5/6 nephrectomy model; qRT-PCR, quantitative reverse transcription polymerase chain reaction; p-mTOR, phosphorylated mammalian target of rapamycin; mTOR, mammalian target of rapamycin.
*P < 0.05, **P < 0.01, ***P < 0.001 vs. control; #P < 0.05, ##P < 0.01, ###P < 0.001 vs. 5/6 Nx.
The extent of autophagy in this context was further investigated, which took advantage of several autophagy biomarkers: Beclin 1, LC3I, LC3II, and p-mTOR. Beclin 1 is an important signaling hub in cell autophagy [29]. Autophagy induces LC3 cleavage by Atg4 to become LC3I, which undergoes further cleavage and lipidation to become LC3II [30]. On the other hand, mTOR is a serine/threonine kinase that inhibits autophagy upon phosphorylation-induced activation [31]. Western blot showed that the nephrectomy caused a drastic decrease in Beclin 1 expression and the LC3II/LC3I expression ratio and a significant increase in p-mTOR expression, all indicating the inhibition of autophagy. In contrast, treatment with melittin effectively increased Beclin 1 expression and the LC3II/LC3I expression ratio and suppressed mTOR phosphorylation (Fig. 3C-H). These results showed that melittin could alleviate the inhibition of autophagy induced by the nephrectomy.
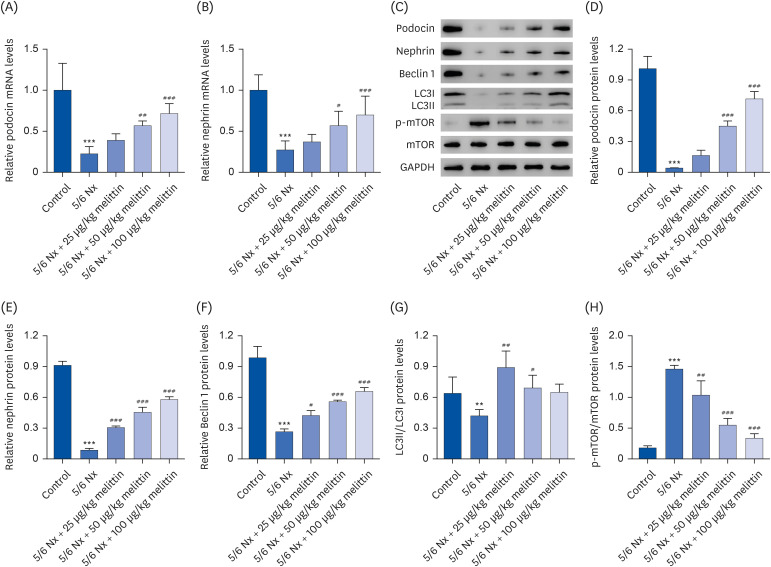
Melittin alleviates AngII-induced podocyte damage by regulating the autophagy signaling pathway
An in vitro podocyte damage model that mimics CRF was established by treating the cells with AngII to investigate the cellular mechanism of the therapeutic effects of melittin. A CCK‐8 assay was used to evaluate the cytotoxicity of melittin on MPC5 cells. As shown in Fig. 4A, 2, 4, and 8 mg/L of melittin did not affect MPC5 cell viability. Furthermore, AngII treatment markedly reduced the MPC5 cell viability compared to the control group (Fig. 4B). On the other hand, melittin restored the viability. In particular, the AngII treatment significantly decreased podocin, nephrin, and Beclin 1 expression and the LC3II/LC3I expression ratio but increased p-mTOR expression. Melittin reversed these in a dose-dependent manner (Fig. 4C-H).
Fig. 4
Melittin alleviates podocyte damage and autophagy inhibition induced by AngII. (A) MPC5 mouse podocyte cells were treated with 2, 4, or 8 mg/L of melittin. The cell viability was evaluated using CCK‐8 assay. MPC5 mouse podocyte cells were treated with 1 μM AngII followed by 2, 4, or 8 mg/L of melittin. Then (B) CCK‐8 assay was used to assess the cell viability and Western blot was used to assess the expression levels of (C, D) podocin, (C, E) nephrin, (C, F) Beclin 1, (C, G) LC3II/LC3I, and (C, H) p-mTOR/mTOR in the cells.
AngII, angiotensin II; MPC5, mouse podocyte clone 5; CCK‐8, cell counting kit 8; p-mTOR, phosphorylated mammalian target of rapamycin; mTOR, mammalian target of rapamycin.
*P < 0.05, **P < 0.01, ***P < 0.001 vs. control or 0 mg/L Melittin; ##P < 0.01, ###P < 0.001 vs. AngII.
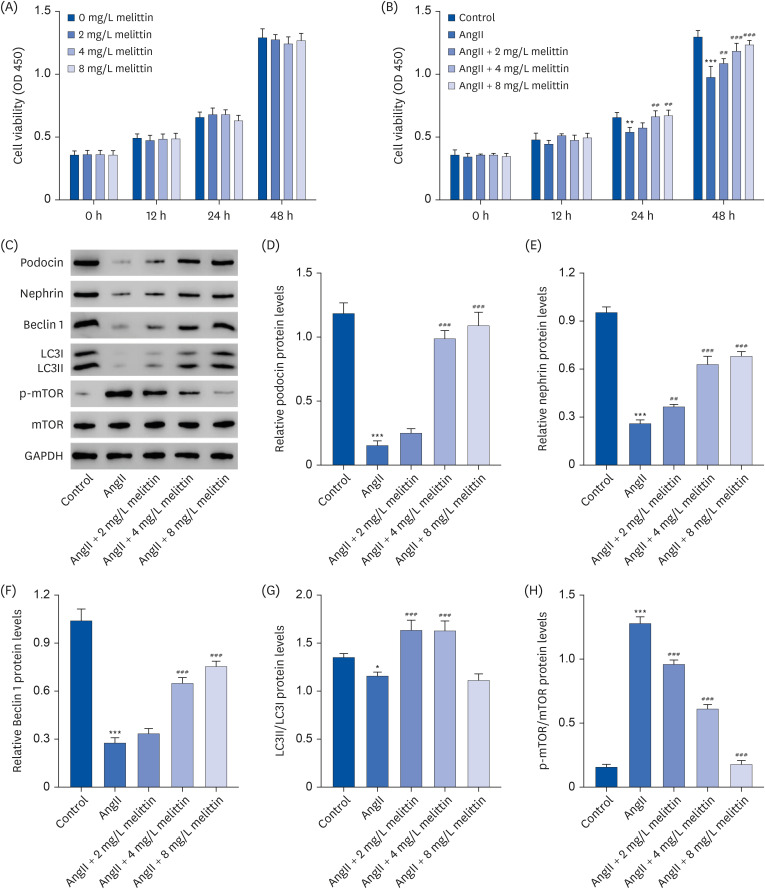
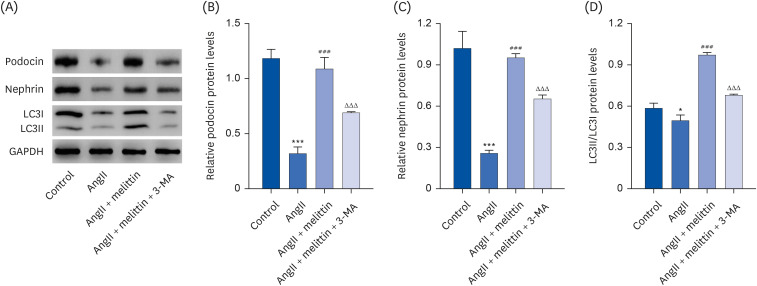
Interestingly, when an autophagy inhibitor 3-MA was added to the cells treated with AngII followed by melittin, it suppressed the extent of autophagy in the cells, as reflected by the decrease in the LC3II/LC3I expression ratio and resulted in lower viability of the podocytes, as reflected by the reduced expression of podocin and nephrin (Fig. 5). These results suggest that AngII induces podocyte damage, at least partially, by suppressing autophagy. Moreover, melittin alleviates AngII-induced podocyte damage by facilitating autophagy.
Fig. 5
Melittin alleviates AngII-induced podocyte damage by regulating the autophagy signaling pathway. Mouse podocyte clone 5 mouse podocyte cells were treated with 1 μM AngII followed by 4 mg/L melittin and 10 mM autophagy inhibitor 3-MA. Western blot was then performed to assess the expression levels of (A, B) podocin, (A, C) nephrin, and (A, D) LC3II/LC3I in the cells.
AngII, angiotensin II.
*P < 0.05, ***P < 0.001 vs. control; ###P < 0.001 vs. AngII; ΔΔΔP < 0.001 vs. AngII + Melittin.
DISCUSSION
CRF is a disease with a complicated pathology, where many different factors play a role, e.g., microvascular environment, blood pressure, and inflammation. This may explain why it is difficult to treat. No new drug has been invented for CRF in the past 10 yrs [32]. Several drug development directions for CRF are ongoing, mainly including modulation of the renal hemodynamics, restoration of vascular integrity, restoration of slit diaphragm integrity, inhibition of inflammation, inhibition of fibrosis, and suppression of metabolic dysregulation, specifically in diabetes patients [32]. Among them, restoring the slit diaphragm integrity could benefit patients by improving the filtration function of the kidney. slit diaphragms are essentially a junction between podocyte foot processes. Therefore, the integrity of slit diaphragms is determined mainly by the proliferation and vitality of podocytes, highlighting podocytes as a potential target for CRF treatment. Indeed, previous studies have shown that podocyte loss contributes directly to kidney function decline, and compounds that protected the podocyte actin cytoskeleton integrity reduced albuminuria in a mouse kidney disease model [3233]. The homeostasis of podocytes relies on the autophagy process, which helps eliminate dysfunctional cells and provides nutrition to healthy cells [34]; a dysfunction of both would lead to multiple renal diseases. On the other hand, whether the podocyte autophagy facilitates or suppresses CRF is unclear. This study examined the mechanism of melittin in alleviating CRF. This paper provides direct evidence showing that podocyte autophagy directly inhibits the symptoms of CRF and that melittin induces podocyte autophagy to alleviate CRF. This result provides cellular-level insights into the therapeutic effects of melittin on CRF and underscores podocyte autophagy as a potential target for treating CRF.
Autophagy describes the process where a cell decomposes and destroys old, damaged, or abnormal molecules in its cytoplasm. As an important factor in podocyte homeostasis, it is maintained at a high level in normal podocytes [35] but is downregulated by renal diseases, such as diabetic nephropathy. Reduced levels of podocyte autophagy can lead to damage in organelles, accumulation of proteins, and blockage of lysosomes. The mTOR is an atypical serine/threonine protein kinase whose activity depends on the nutrient status and redox levels [5]. mTOR is the main regulator of cellular growth, proliferation, protein and lipid synthesis, autophagy, and metabolism. Promoting autophagy by inhibiting the mTOR pathway has renoprotective effects in rats with CRF [6]. On the other hand, inhibiting autophagy by activating the mTOR pathway alleviates renal fibrosis and protects renal function in rats with CRF [7]. This study provided evidence supporting the view that podocyte autophagy alleviates CRF [363738]. This is consistent with the role of podocyte autophagy in other renal disease models and emphasizes podocyte autophagy as a promising target for treating CRF in addition to other related renal diseases.
Melittin has been used widely in treating renal diseases because of its anti-inflammatory and anti-fibrotic effects. Nevertheless, melittin is a cytolytic peptide, and its intravenous administration causes non-specific cell toxicity (such as cell membrane pore formation) and hemolysis of red blood cells [39]. In the present study, rats were treated with melittin (25, 50, and 100 μg/kg) via intraperitoneal administration. Similarly, intraperitoneal administration of 0.75 and 1.5 mg/kg doses of melittin could not induce significant changes in the immune parameters. Hence, melittin was safe for the immune system [40].
The function of melittin in autophagy is controversial in biological systems. A facilitating role of melittin in autophagy has been found in human fibroblast-like synoviocytes and cancer cells, such as hepatocellular carcinoma cells and lung cancer cells, which contributes to an anti-arthritic and anti-tumor effect [144142]. Nevertheless, melittin attenuated 3-AP-induced autophagy and improved the motor function, implying a potential for treating cerebellar ataxia [16]. Melittin inhibits the proliferation and migration of cancer cells by inhibiting the PI3K/AKT/mTOR signaling pathway [434445]. It ameliorates inflammation in acute liver failure and acne vulgaris by inactivating the mTOR signaling pathway [4647]. This study confirmed that melittin could facilitate autophagy and inhibit the mTOR signaling pathway in podocytes, alleviating CRF. Moreover, 2, 4, and 8 mg/L of melittin did not affect the MPC5 cell viability, but it inhibited the AngII-induced decrease in the viability of MPC5 cells.
Melittin has shown cytotoxic effects on other cells [1448]. Considering that multiple factors contribute to CRF and the varied effects of melittin, it would be interesting to test whether the therapeutic effects of melittin on CRF also involve other mechanisms, e.g., inhibiting inflammation and fibrosis.
In conclusion, melittin has therapeutic effects on CRF related to its facilitating function on podocyte autophagy. This study revealed the cellular mechanism for how melittin can treat CRF and highlights podocyte autophagy as a target for drug development against CRF.
 XML Download
XML Download