

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Nasopharyngeal carcinoma (NPC) is a distinctive type of head and neck cancer due to its geographic distribution. NPC affects an estimated 130,000 patients worldwide, accounting for 0.7% of all cancer cases in 2018. The highest rates are in Southern China, Southeast Asia, and North Africa, where NPC is consistently associated with Epstein‒Barr virus infection [1].
Radiotherapy alone is adequate for the early stages of NPC, but radiation combined with platinum-based chemotherapy is the mainstay treatment for locally advanced NPC [2-5]. Distant metastasis is the predominant cause of treatment failure and accounts for cancer-specific mortality among approximately 70% of cancer patients [6,7]. Current chemotherapy agents used to treat advanced NPC have limitations regarding therapeutic potential and the duration of response. Malignant tumors eventually develop resistance. The search for the most effective drugs and optimal treatment regimens remains a significant challenge.
Oceans cover more than 70% of the Earth’s surface. The diversity of marine flora and fauna is an untapped source of medical discovery. Previous studies have reported the broad-spectrum biological effects of natural compounds isolated from Taiwanese soft corals. For example, capnellene and lemnalol have anti-inflammatory and neuroprotective properties [8,9]. Cembrane-type diterpenes, such as sinularin and 5-epi-sinuleptolide, also show anticancer activity [10-12].
Dihydroaustrasulfone alcohol (DA) is the synthetic bioactive precursor of austrasulfone, which is isolated from the Formosan (Taiwanese) soft coral Cladiella australis [13]. Previous studies have shown the therapeutic properties of DA, including anti-inflammatory and neuroprotective properties, the attenuation of neuropathic pain, the ability to treat multiple sclerosis and non-small cell lung cancer [13,14], the alleviation of atopic dermatitis [15], and inhibiting angiogenesis in atherosclerosis [16]. However, it is unknown whether DA has antitumor effects on NPC. Previously, researchers found that the PI3K and MAPK pathways play critical roles in the anticancer effect of DA on A549 cells [13]. In addition, we further investigated the apoptosis-related and reactive oxygen species (ROS)-dependent mechanisms underlying the therapeutic effects of DA while evaluating its anticancer activity against NPC and the PI3K pathway in this study.
METHODS
Cell lines and cultures
The NPC-39 and NPC-BM cell lines were used in this study, which are NPC cells and were obtained from Dr. Mu-Kuan Chen at Changhua Christian Hospital (Taiwan). Both cell lines were cultured in RPMI-1640 medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific) and 1% penicillin‒streptomycin (Biological Industries) and maintained in a 5% CO2 and 37°C incubator.
Dihydroaustrasulfone alcohol
DA was synthesized as reported previously [13]. In brief, the initial I,4-addition of 2-mercaptoethanol to methyl vinyl ketone to generate the corresponding sulfide compound and the subsequent oxidation of this sulfide with m-chloroperoxybenzoic acid were performed. Nuclear magnetic resonance spectroscopy was used to characterize the product’s structure (Supplementary Fig. 1), and the sample was stored at −20°C following dissolution in dimethyl sulfoxide (DMSO).
Cell viability assays
Cells in culture plates were treated with DA for 24 h, followed by an MTT assay. MTT solution (5 mg/ml; Merck Millipore) was directly added to the culture wells and incubated for 4 h. The formazan was dissolved in 300 µl of DMSO/well, and the absorbance was measured at 540 nm with an ELISA device.
Colony formation assay
The protocol of the colony-formation assay in this study was adapted from previous reports with necessary modifications [17]. NPC cells in six-well plates were incubated for 24 h, followed by incubation with DA-containing complete medium for seven days to visualize the colonies. The colonies were fixed with acetic acid and methanol (1:3) and stained with 2% crystal violet before being counted under an inverted microscope (Olympus).
Apoptosis and cell cycle detection
NPC cells were treated with DA-containing medium for at least 24 h before being harvested for staining with propidium iodide (PI) (50 µg/ml) and Annexin V-FITC (20 µg/ml) (Biolegend) at room temperature for 15 min. Apoptotic cells were quantified by an Accuri C5 flow cytometer (BD Biosciences).
CaspGLOW fluorescein active caspase staining kits (Biovision) were used to measure caspase activities, including caspase-3, -8, and -9. Cells were treated with DA-containing medium for 24 h, harvested, centrifuged at 1,200 rpm, washed with phosphate-buffered saline (PBS), and finally incubated with a caspase staining kit according to the manufacturer’s instructions before being analyzed by flow cytometry. In addition, Z-DEVD-FMK, an inhibitor of caspase-3, was used to treat the cells 2 h before DA treatment to rescue NPC cell viability, which was measured by MTT assays. For cell cycle analysis, intracellular PI staining was used to determine the DNA content linked to the cell cycle status.
Western blotting
DA-treated cells were collected and lysed with ice-cold RIPA buffer (MedChemExpress), and a phosphatase inhibitor (Sigma-Aldrich) was added to prevent the degradation of phosphorylated proteins. Forty micrograms of total protein was subsequently separated by SDS‒PAGE and hybridized with the following antibodies after being transferred to PVDF membranes (Merck Millipore) and blocked with blocking buffer (Visual Protein). The primary antibodies were diluted 1:100 and included anti-GAPDH (Abcam, cat. no. ab9484), anti-Bax (Biolegend, cat. no. 633602), anti-Bcl-2 (Cell Signaling, cat. no. 2870s), anti-cleaved-PARP (Cell Signaling, cat. no. 9541s), anti-pPI3K (Cell Signaling, cat. no. 13857s), anti-PI3K (Santa Cruz, cat. no. sc-136208), anti-AKT (Sigma-Aldrich, cat. no. 05-1003), and anti-pAKT (Santa Cruz, cat. no. sc-7985); then, a 1:2,000 dilution of horseradish peroxidase-conjugated secondary antibodies (cat. no. 111-035-003, Jackson ImmunoResearch Laboratories, Inc.) was added and incubated with enhanced chemiluminescence (GE Healthcare Life Sciences) in a Hansor Luminescence Image system (Hansor).
Mitochondrial membrane potential and cytochrome c release assays
DA-treated NPC cells were analyzed with a JC-1 Assay Kit (Invitrogen Life Technologies) and FITC-anti-cytochrome c antibodies to determine mitochondrial membrane potential and cytochrome c release, respectively. Briefly, the cells were incubated with medium containing JC-1 fluorescent dye or cytochrome c solution, including the specific antibody, for 10 min in the dark at 37°C prior to analysis with an Accuri C5 flow cytometer. The results were analyzed using BD Accuri C6 Software version 1.0.264.21.
Detection of death receptors
DA-treated cells were trypsinized and stained with anti-DR4-FITC, anti-DR5-FITC, and anti-Fas-FITC (eBioscience) antibodies at 4°C for 30 min. The expression was detected using an Accuri C5 flow cytometer and analyzed using BD Accuri C6 Software version 1.0.264.21.
Preparation of retroviral particles
The pBabe.puro-Myc-Flag-PKBA/AKT plasmid was obtained from Addgene (Watertown) and transfected into HEK-293T cells with jetPEM transfection reagent for 24 h to generate viral particles, including gag-pol and VSV-G proteins, as instructed by the manufacturer. The viral particles were collected to infect NPC-39 cells (5 × 105/well) for 48 h, and the harvested cells were plated in culture plates for further experiments.
Measurement of ROS
Intracellular ROS concentrations were determined using the peroxide-sensitive fluorescent probe DCFH–DA. In brief, DA-treated NPC cells were collected, washed twice with PBS, and incubated with 1 µl of DCFDA in 1 ml of HBSS buffer (Invitrogen) for 30 min in an incubator. After further washing with PBS, the level of ROS was determined by flow cytometry and the corresponding software. N-acetylcysteine (NAC, Sigma-Aldrich)-treated cells were used as a positive control to compare the ROS scavenging activity of DA, and MTT assays were used to determine cell viability.
Statistical analysis
Unless indicated elsewhere, all data are presented as the mean ± standard deviation. Data in groups were further analyzed for significant differences by one-way ANOVA followed by Tukey’s posttest or unpaired two-tailed t-test. A p-value less than 0.05 was considered to be statistically significant.
RESULTS
DA decreased cell viability and colony formation in human NPC cell lines
As shown in Fig. 1B, treatment of NPC-39 and NPC-BM cells with DA (Fig. 1A) (0−100 µM) for at least 24 h reduced their proliferation in a dose-dependent manner. The MTT assay showed that the IC50 values of DA were 27.1 ± 4.6 and 45.4 ± 7.3 µM in NPC-39 and NPC-BM cells, respectively. In parallel, colony formation assays demonstrated that DA treatment for one week inhibited NPC-39 and NPC-BM cells from forming more colonies compared to control cells (Fig. 1C, D). These results suggest that DA could reduce the growth of NPC cells. We observed that NPC-39 cells appeared more susceptible to DA treatment than NPC-BM cells. As a result, we assume that the NPC-39 cell line is more adequate for the following experiments to better illustrate the anticancer effects of DA in vitro.
DA treatment enhanced apoptosis in NPC-39 cells
To determine whether the death of NPC cells induced by DA resulted from apoptosis, we stained cells with Annexin V conjugated FITC and PI to label DNA contents, followed by flow cytometry to analyze the degrees of apoptosis and cell-cycle distribution with or without DA treatment for 24 h. The results in Fig. 2A and 2C illustrate that dead cells, representing the sub-G1 population, were significantly increased following DA treatment. Annexin V-positive cells were also increased by DA, indicating higher apoptotic ratios of NPC-39 cells (Fig. 2B, D). These results were further confirmed by determining poly (ADP-ribose) polymerase (PARP) protein expression levels via Western blot analysis, and we observed higher expression of PARP in NPC-39 cells, suggesting that the apoptotic protein marker was activated and elevated after DA treatment in a dose-dependent manner (Fig. 2E, F).
Furthermore, we attempted to explore the apoptotic pathways involved in the effect of DA treatment on NPC-39 cells. We directly measured the activities of caspase-9, caspase-8, and caspase-3 and found that DA dose-dependently increased the activities of all three caspases, including caspase-9 (Fig. 3A, B), caspase-8 (Fig. 3A, C), and caspase-3 (Fig. 3A, D), in NPC-39 cells. Consistently, we added the pancaspase inhibitor Z-VAD-FMK to DA-induced cells to determine whether the inhibitor could reverse apoptosis levels. The MTT assays demonstrate that Z-VAD-FMK significantly rescued the viability of NPC-39 cells after DA treatment (Fig. 3E), proving that the growth of NPC cells was slowed and that caspase-mediated apoptosis was promoted.
DA induced mitochondrial dysfunction in NPC-39 cells
Mitochondrial dysfunction is generally associated with apoptosis. We stained cells with JC-1 dye to measure the mitochondrial membrane potential to understand whether DA-induced apoptosis involves mitochondrial dysfunction. The results showed that mitochondria in cells without DA treatment retained a high level of matrix metalloproteinase (MMP)-dependent JC-1, and this JC-1 aggregation was dose-dependently decreased by DA treatment (Fig. 4A, C). Moreover, the results in Fig. 4B and 4D show that cytochrome c release into the cytosol in DA-treated mitochondria was increased to a higher level than that in mitochondria without DA treatment. In addition, the protein expression levels of Bax were increased, and the antiapoptotic protein Bcl-2 was decreased following DA treatment (Fig. 5A, B), which was consistent with the increase in truncated BH3 interacting-domain death agonist (tBid) protein levels, indicating that mitochondrial dysfunction was involved in DA-induced carcinoma cytotoxicity (Fig. 5).
DA induced death receptor expression in NPC-39 cells
Given our previous data suggesting that caspase-8 activity was activated by DA (Fig. 3A, C) and the increase in truncated Bid (Fig. 5), both of which are associated with the extrinsic apoptotic pathway and are highly correlated with death receptors such as DR4, DR5, and Fas/CD95 (death receptor protein), we attempted to confirm the expression of the aforementioned death receptor proteins with and without DA treatment. We stained the indicated cell surface markers and analyzed the numbers of positive cells. The results revealed that the surface levels of DR4 (Fig. 6A, B), DR5 (Fig. 6A, C), and Fas (Fig. 6A, D) protein on the surface of NPC-39 cells increased in a concentration-dependent manner after DA treatment.
DA suppressed the PI3K/AKT signaling pathway in NPC-39 cells
We wondered if the survival signaling pathway in NPC cells was interfered with and inhibited by DA, and so we examined the phosphorylation levels of the PI3K protein and its downstream protein AKT to examine the mechanisms of DA. The results in Fig. 7A and 7B show that phosphorylated PI3K and AKT were profoundly and dose-dependently decreased by DA, while the total protein levels of PI3K and AKT remained constant, suggesting that DA could inhibit the PI3K/AKT pathway in NPC-39 cells, leading to the cells receiving fewer survival signals. Therefore, we transfected NPC-39 cells to create cells constitutively expressing activated PI3K/AKT (Fig. 7C) to validate our findings. We found that overactivation of the AKT pathway overcame the adverse and cytotoxic effects of DA treatment, as evidenced by fewer Annexin V-positive cells (Fig. 7D, E) and rescued cell viability compared with cells treated with DA alone (Fig. 7F). These findings strongly suggest that the PI3K/AKT pathway is involved in DA-mediated induction of apoptosis or cytotoxicity in NPC-39 cells.
ROS production involved in the effects of DA on growth inhibition and PI3K/AKT signaling
Because ROS can affect cellular redox status and mitochondrial membrane potential, we needed to confirm whether DA could promote ROS production in NPC-39 cells. As shown in Fig. 8, the DCFDA fluorescence intensity of DA-treated cells was significantly and dose-dependently increased compared to untreated cells. In addition, pretreatment of NPC-39 cells with the oxidant scavenger NAC rescued cell viability, and DA-treated cells without NAC exhibited higher cytotoxicity (Fig. 9A). Furthermore, a decrease in apoptosis was also observed in NAC-treated cells by flow cytometric analysis, suggesting that ROS were a key factor in the therapeutic effect of DA (Fig. 9B, C). Notably, NAC treatment significantly reversed phospho-AKT expression (Fig. 9D), suggesting that DA could inhibit cell growth and inactivate the phosphorylation of PI3K/AKT signaling, both of which were associated with ROS levels induced by DA.
DISCUSSION
The ocean accounts for over two-thirds of the Earth’s surface and provides valuable natural resources with biodiversity. Many marine compounds derived from organisms have been shown to have anticancer effects, demonstrating their potential for cancer treatment [18-22]. DA, which is derived from coral Cladiella australis, has been shown to exert cytotoxic effects against cancer cells. Chen et al. [14] demonstrated that DA inhibited the migration of A549 lung cancer cells and MMP-2/-9 expression. The researchers also found that this compound attenuated the phosphorylation of PI3K, AKT, p38, ERK1/2, and c-Jun N-terminal kinase 1/2, as well as PI3K expression [14]. However, the antitumor effect and detailed mechanism of DA in human NPC cells are unknown. We examined the proliferation of DA-treated NPC cells in vitro, and the results indicated a significant reduction in the viability of NPC-39 and NPC-BM cells. We found that NPC-39 cells were more sensitive to DA treatment and therefore more suitable for investigating anticancer effects in vitro.
Apoptosis is a mode of programmed cell death promoted by anticancer treatments. Our data demonstrated that DA induced NPC-39 apoptosis, which was confirmed by flow cytometric gating with Annexin V-FITC, PI staining, and examination of a sub-G1 population. Caspases are protease enzymes that play an essential role in apoptosis. The caspase cascade is an intrinsic or an extrinsic apoptosis pathway.
Members of the TNF superfamily, including TNF, Fas ligand, and TNF-related apoptosis-inducing ligand (TRAIL), initiate the extrinsic apoptosis pathway. These ligands induce apoptosis by binding to their respective receptors: Fas (CD95), TNFR1, TRAIL-R1 (DR4), and TRAIL-R2 (DR5). Trimerization of the receptor and clustering of the receptor’s intracellular death domain (DD) leads to the recruitment of the adaptor molecule Fas-associated death domain or TNF receptor-associated protein with death domain, which forms a death-inducing signaling complex (DISC) and subsequently activates caspase-8 and -10. Activated caspase-8 and -10 then cleave caspase-3, which leads to the cleavage of death substrates. DISC-mediated caspase activation is regulated by the cellular FLICE-like inhibitory protein [23-25].
The intrinsic pathway, which is also known as the mitochondrial pathway, involves activating the proapoptotic arm of the Bcl-2 gene superfamily. Mitochondrial outer membrane permeabilization and the release of cytochrome c into the cytosol occur via the intrinsic route. Cytochrome c interacts with the adaptor APAF-1 in the cytosol, producing an apoptosome that activates caspase-9, an apoptosis-initiating protease. Other executioner caspases are cleaved and activated by Caspase-9 (caspase-3, caspase-6, and caspase-7) [26-29].
Our study showed increased caspase-9, caspase-8, caspase-3, and PARP activity in DA-treated NPC-39 cells. In addition, a pancaspase inhibitor (Z-VAD-FMK) was proven to have a protective effect. These results verified that caspase-mediated apoptosis was involved in the treatment process. Increased expression of DR4, DR5, and FAS was also identified in the DA-treated cells, indicating that DA is involved in the extrinsic apoptosis pathway. Furthermore, decreased JC-1 and MMP aggregation and increased cytochrome c levels suggested mitochondrial apoptosis driven by DA.
Intrinsic and extrinsic apoptosis signaling pathways communicate with one another. The proapoptotic Bcl-2 family member Bid is activated by caspase-8-mediated cleavage. The interaction of Bax and Bak with caspase-8 leads to the separation of Bid and translocation of tBid to mitochondria, resulting in increased cytochrome c release, providing a plausible molecular link between the extrinsic and intrinsic pathways [28].
Bcl-2 (B-cell lymphoma 2), which is encoded by the BCL2 gene, is a critical protein in the Bcl-2 family that regulates cell death. Bcl-2 drives this process by inhibiting or inducing apoptosis. The proapoptotic proteins in the BCL-2 family, including Bax and Bak, promote mitochondrial outer membrane permeability and the release of cytochrome c and ROS, which are essential signals in the apoptosis cascade. These proapoptotic proteins are activated by BH3-only proteins but inhibited by antiapoptotic BCL-2 and its relative BCL-XL [27,30,31]. This study demonstrated increased proapoptotic Bax expression and decreased antiapoptotic BCL-2 expression in DA-treated NPC-39 cells. The findings suggest that DA promotes apoptosis in the balance between proapoptotic and antiapoptotic effects.
The PI3K/AKT signaling pathway is considered the “survival pathway” because of its essential role in cell differentiation, proliferation, and survival. AKT is upregulated in various malignant tumors, including gastric, colon, breast, ovarian, and prostate cancers [32,33]. The PI3K/AKT pathway has become an important target in cancer therapy. We demonstrated the inhibitory effects of DA on the expression of pPI3K and pAKT in NPC-39 cells, suggesting that DA could block the activation of the PI3K/AKT pathway. Moreover, we transfected DA-treated NPC-39 cells with an active form of AKT cDNA, and showed reduced apoptosis and the return of cell viability. This event aided the inactivation of the PI3K/AKT pathway by DA.
While we did not investigate the autophagy pathway, it is worth noting that an increase in ROS can downregulate the autophagic pathway, particularly ULK1 signaling, and the phosphorylation of PI3K mediated by ULK1 was also suppressed in a previous study [34]. In line with previous findings, we observed that the downregulation of PI3K and activation of apoptotic signaling by DA were highly dependent on ROS production. In summary, a model of the mechanism by which DA inhibits the growth of NPC could be proposed: DA treatment induces ROS production, which mitigates autophagy and activates apoptosis in carcinoma cells, eventually leading to the downregulation of PI3K.
Because ROS play a vital role in the survival of cancer cells, we explored ROS production in DA-treated NPC-39 cells. Cancer cells have higher ROS levels than normal cells because of their enhanced metabolism and persistent pro-oxidative state, which leads to intrinsic oxidative stress. Increased ROS levels are closely related to cancer initiation, metastasis, and drug resistance. However, elevated ROS also increase the susceptibility of tumor cells to oxidative stress and trigger cell death, which could be a potential strategy for cancer therapy [35,36]. Mitochondria are a possible ROS generation site that can leak single electrons to oxygen, convert them into superoxide anions and reduce mitochondrial transmembrane potential (ΔΨm). The mitochondrial apoptosis pathway is then triggered. In our study, ROS production was promoted in DA-treated NPC-39 cells, and the oxidant scavenger NAC was found to reduce DA-induced cell viability. This evidence suggests ROS-mediated cell death in DA-treated NPC-39 cells.
Previous studies have shown that ROS can repress the PI3K/AKT pathway [37-42]. To determine the relationship between PI3K/AKT activation and ROS production in response to DA treatment, we used the ROS scavenger NAC to investigate the activation of PI3K/Akt in DA-treated NPC-39 cells. We found that NAC significantly reduced DA-induced apoptosis and reversed pPI3K/AKT expression. These results indicated that ROS mediated DA-induced apoptosis and the inactivation of PI3K/AKT signaling in NPC-39 cells.
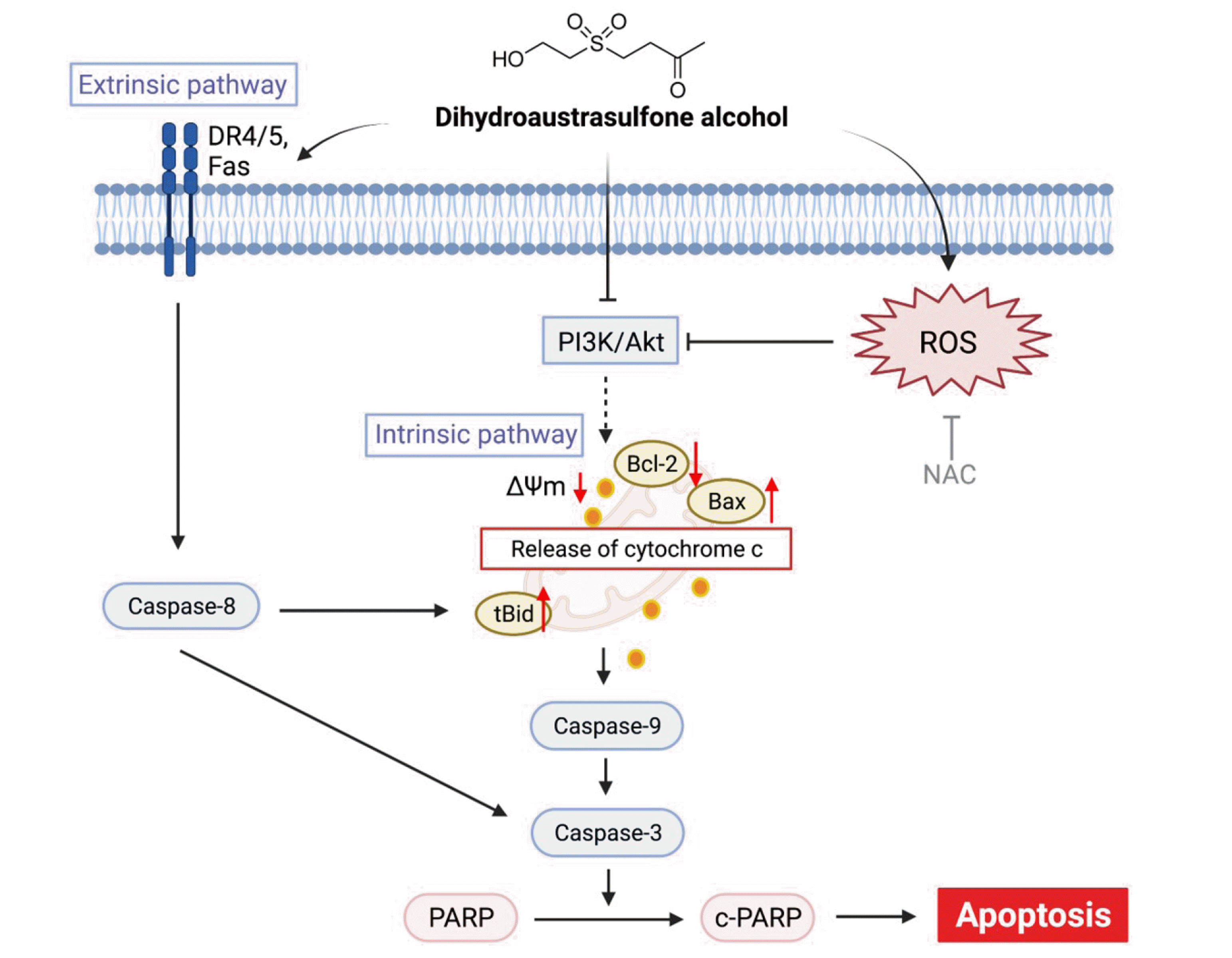
We demonstrated that DA inhibits the growth and proliferation of NPC-39 cells by inducing G1 phase cell cycle arrest. In addition, DA induces intrinsic and extrinsic pathways involving caspase-sdependent apoptosis in NPC-39 cells through ROS accumulation and the repression of pPI3K and pAKT. We examined ROS-mediated inactivation of the PI3K/AKT pathway in apoptosis in DA-treated NPC-39 cells. Fig. 10 shows the proposed mechanism of DA-induced apoptosis in NPC-39 cells. This is the first report showing the inhibitory effect of DA on cell proliferation and the induction of apoptosis in NPC cells in vitro. Our findings suggest that DA has therapeutic potential for treating NPC.
SUPPLEMENTARY MATERIALS
Supplementary data including one figure can be found with this article online at https://doi.org/10.4196/kjpp.2023.27.4.383.



 XML Download
XML Download