

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
High blood glucose levels increase the risk of developing serious diseases affecting the heart, blood vessels, eyes, kidneys, nerves, and teeth [12]. Especially, diabetes is a leading cause of kidney failure, cardiovascular disease, blindness, and lower limb amputation [2]. Since high blood glucose levels are a probable cause of damage to the cardiovascular system, maintaining normal glucose levels could help delay or prevent diabetic complications. Damage to large blood vessels could lead to heart attacks, stroke, and peripheral arterial diseases [34]. Microvascular diseases, which refer to injuries to small vessels, can adversely affect the eyes, kidneys, and nerves [45]. Diabetic nephropathy (DN) is characterized by failure of nephron functioning and albuminuria, which are primarily important risk factors for kidney diseases [6]. Diabetic retinopathy (DR) causes microvascular alterations in the retina, eventually leading to visual impairment or vision loss [78]. Moreover, diabetic neuropathy is a diabetic complication that reduces the quality of life due to pain, sensory loss, gait instability, and foot ulceration and amputation [9].
DN is a multifactorial kidney disorder associated with glomerulosclerosis and fibrosis, along with mesangial expansion and glomerular basement membrane thickening [10]. In progressive DN, the glomerular filtration barrier is seriously damaged, causing persistent proteinuria and abnormal renal function [1112]. Plasma proteins filtered into the urine affect the renal tubules and instigate interstitial scarring, leading to tubular fibrosis and, ultimately renal failure [13]. Renal fibrosis occurs due to prolonged injury and wound-healing dysregulation of kidneys, resulting in excess accumulation of extracellular matrix (ECM) proteins, including collagens [14]. Thus, aberrant fibrosis is positioned as a potential mechanism for chronic renal failure in DN. Blocking signaling pathways that evoke unfavorable renal fibrosis could slow down or prevent DN. Several signaling pathways that involve transforming growth factor-β, connective tissue growth factor (CTGF), and Wnt/β-catenin have been suggested [1516]. Understanding the cellular and molecular mechanisms of renal fibrosis provide versatile visions for the development of new therapeutic strategies for DN [15].
Hyperglycemia induces loss of intramural pericytes and thickening of basement membranes in retinal vessels, leading to the dysfunction of the blood-retinal barrier [717]. During DR progression, the damaged vessels are leaky, resulting in penetration of blood and fluid exudates into the macula and retina, leading to diabetic macular edema and blurry vision [718]. Vitreous hemorrhage, fibrovascular tissue formation, and retinal detachments are often associated with abnormal neovascularization in the retina [718]. Retinal neovascularization is also known to occur due to damage of retinal vessels under retinal ischemic conditions [19]. Retinal tissue hypoxia leads to the upregulation of several hypoxia-regulated proteins, including vascular endothelial growth factor (VEGF) and angiopoietins, which may be involved in increased vascular permeability and pathological angiogenesis [20]. Thus, VEGF has been identified as a validated target for several retinal vascular diseases, and anti-angiogenic neutralization of VEGF by antagonists may be beneficial in the treatment of vascular leakage and macular edema in DR [21]. However, anti-VEGF agents may also cause ocular side effects after intraocular administration.
So far, several efforts have been made to treat diabetic complications by developing experimental therapeutic targets. The causal role and signaling sites of oxidative stress in the development and progression of diabetic complications have been emphasized [22]. Polyphenols present in natural products have gained much attention in preventing diabetes-associated pathologies. Our previous study revealed that the multifunctional chrysin (5,7-dihydroxyflavone, Fig. 1A) inhibited tubulointerstitial fibrosis in diabetic kidneys by blocking the epithelial to mesenchymal transition [23]. Chrysin administration lowered the fasting blood glucose and 24 h urine volume in db/db mice [23]. In addition, exposure to chrysin antagonized retinal neovascularization in diabetic mouse models [24]. Chrysin is soluble in a few organic solvents but is practically insoluble in water [25]. Therefore, effective solubilization of chrysin is critical for its pharmaceutical application [26]. The current study attempted to provide a preventive effect of the newly-synthesized 5,7-di-O-acetylchrysin (diacetylchrysin, Fig. 1A) having increased water solubility, on intervening with diabetic complications of DN and DR [26].
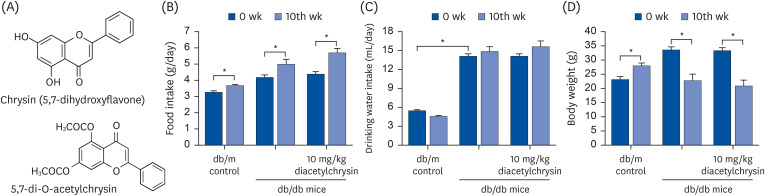 | Fig. 1Chemical structures of chrysin and diacetylchrysin (A), and changes in food intake (B), drinking water intake (C), and body weight (D) of db/m controls and db/db mice after the 10 week-supplementing challenge with diacetylchrysin. The db/db mice were supplemented with 10 mg/kg diacetylchrysin for 10 weeks. The db/m mice were introduced as control animals. The data are presented as mean ± SEM for each treatment group (n = 9).SEM, standard error of mean.
*P < 0.05 relative to 10th week or db/m control mice.
|
Go to : 
MATERIALS AND METHODS
Materials
Chrysin was obtained from Sigma-Aldrich Chemical (St. Louis, MO, USA). Rabbit polyclonal antibodies of collagen type IV, membrane type 1-matrix metalloproteinase (MT1-MMP) and tissue inhibitor of metalloproteinase 2 (TIMP-2), and goat polyclonal antibodies of Zona occluden (ZO)-1 and platelet endothelial adhesion molecule (PECAM)-1, were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Rabbit polyclonal CTGF antibody was provided from Peprotech (Rocky Hill, NJ, USA). Goat polyclonal VEGF antibody was obtained from R&D systems (Minneapolis, MN, USA), and rabbit monoclonal VEGF receptor 2 (VEGFR2) antibody was purchased from Cell Signaling Technology (Beverly, CA, USA). Rabbit polyclonal vascular endothelial cadherin (VE-cadherin) antibody was supplied by Abcam Biochemicals (Cambridge, UK). Mouse monoclonal β-actin antibody was obtained from Sigma-Aldrich Chemical. Horseradish peroxidase (HRP)-conjugated goat anti-rabbit immunoglobulin G (IgG), goat anti-mouse and donkey anti-goat IgG were purchased from Jackson ImmunoResearch Laboratories (West Grove, PA, USA).
Synthesis of 5,7-di-O-acethylchrysin
The compound diacetylchrysin was synthesized as described in a previous study [26]. Briefly, acetic anhydride (10 mM) was added dropwise to a solution of 10 mM chrysin in 50 mL pyridine. After 2 h of reaction at room temperature with continuous stirring, the solvent was removed using a rotary evaporator at 40°C. The residue was dissolved in dichloromethane, washed three times in 1 M HCl, followed by washing with saturated sodium bicarbonate solution and water to neutralize. The organic phase was separated, dried over MgSO4, and vacuum-concentrated. The residue was purified by silica gel column chromatography, eluting with dichloromethane/MeOH (10:0 to 9.5:1.5, v/v) to obtain diacetylchrysin.
The solubilities of chrysin and diacetylchrysin in water were determined to be 0.03 mM and 0.265 mM, respectively, indicating that the diacetylchrysin solubility was 8.83-fold higher than chrysin [26].
In vivo animal experiments
Adult male db/db mice (C57BLKS/+Leprdb Iar; Jackson Laboratory, Sacramento, CA, USA) and their age-matched non-diabetic db/m littermates (C57BLKS/J; Jackson Laboratory) were employed in the current study. Mice were housed at the animal facility of Hallym University and kept under specific pathogen-free conditions on a 12 h light/12 h dark cycle at 23 ± 1°C with 50 ± 10% relative humidity. All animals were fed a standard pellet laboratory chow diet (Cargill Agri Purina, Inc., Seongnam, Korea) and were provided with water ad libitum. Since the mice began to develop diabetes (hyperglycemia) at 7–8 weeks, the db/db mice used in this study were at 7 weeks of age. The animals were allowed to acclimatize for a week before beginning the experiments. Mice were divided into 3 subgroups (n = 9–10 for each subgroup): Group 1, non-diabetic db/m controls; Group 2, db/db mice orally administrated 10 mg/kg BW diacetylchrysin (dissolved in phosphate-buffered saline [PBS]) daily for 10 weeks; and Group 3, db/db mice orally administrated 10 mg/kg chrysin (dissolved in dimethyl sulfoxide [DMSO]) daily for 10 weeks.
All experiments were approved by the Committee on Animal Experimentation of Hallym University, and performed in compliance with the University Guidelines for the Care and Use of Laboratory Animals (hallym 2020-52). There was no mortality, and no apparent signs of exhaustion were observed during the experimental period.
Sampling of blood and urine
Food intake, drinking water intake, and body weight of mice were measured every week during the 10 weeks of supplementation. Food intake, drinking water intake, and body weight of db/db mice were much higher than those of db/m controls, regardless of the supplementation of 10 mg/kg diacetylchrysin (Fig. 1B-D). It should be noted that despite the increased food intake, the administration of 10 mg/kg diacetylchrysin to mice significantly lowered the body weight after 10 weeks (Fig. 1D).
A blood glucose meter (ACCU-CHEK Performa; Roche Diagnostics, Mannheim, Germany) was used to measure the fasting blood glucose in the mouse tail veins every other week. Blood glycated hemoglobin A1c (HbA1c), a biomarker indicating development of diabetic complications, was measured using the high-performance liquid chromatography technique. The levels of blood glucose and plasma HbA1C were highly elevated in db/db mice (Fig. 2A and B), but treatment of diacetylchrysin did not influence these levels. The oral glucose tolerance curve in diacetylchrysin-challenged mice was similar to that of untreated db/db mice (Fig. 2C). The 24 h urine collection was carried out using metabolic cages. The 24 h urine volume of db/db mice was much higher than the db/m controls (Fig. 2D). Urinary albumin excretion was determined using an enzyme-linked immunosorbent assay kit (Abcam Biochemicals), according to the manufacturer’s instructions. Although diacetylchrysin did not reduce the urine volume significantly, the urinary albumin level was markedly curtailed (Fig. 2E).
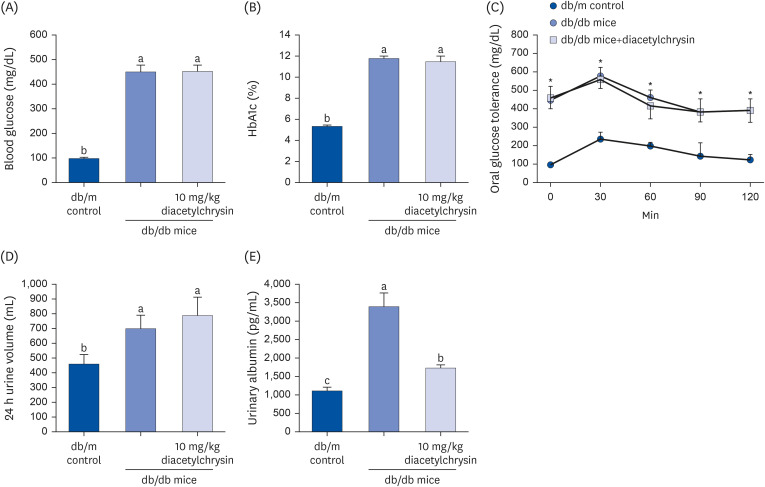 | Fig. 2Blood levels of fasting glucose (A) and glycated hemoglobin level (HbA1c, B), oral glucose tolerance curve (C), 24 h urine volume (D), and urinary albumin excretion (E) of db/db mice treated with diacetylchrysin. The db/db mice were supplemented with 10 mg/kg diacetylchrysin for 10 weeks. The db/m mice were introduced as control animals. The blood glucose levels and HbA1c, 24 h urine volume, and urinary albumin excretion were measured after the 10 weeks treatment of diacetylchrysin. The values (mean ± SEM, n = 9) not sharing a common small letter are significantly different at P < 0.05.HbA1C, hemoglobin A1c; SEM, standard error of mean.
*P < 0.05 relative to db/m control mice.
|
Masson’s trichrome staining
Paraffin-embedded renal tissues of glomeruli and tubules were sectioned at 5 μm thickness, de-paraffinized, and stained with Masson trichrome for the histological visualization of collagen fibers and muscle fibers. The stained tissue sections were examined using an optical Axiomager microscope (Zeiss, Göttingen, Germany), and 5 images were taken for each section.
Western blot analysis
Western blot analysis was conducted using kidney tissue extracts. Briefly, tissue extracts of kidneys and eyes were prepared in a lysis buffer containing 1 M β-glycerophosphate, 1% β-mercaptoethanol, 0.5 M NaF, 0.1 M Na3VO4, and protease inhibitor cocktail. Tissue extracts containing equal amounts of proteins were electrophoresed on 6–15% SDS-PAGE and transferred onto a nitrocellulose membrane. Nonspecific binding was blocked with 5% skim milk for 3 h. The membrane was incubated overnight at 4°C with each primary antibody of target proteins being examined, followed by washing in a Tris-buffered saline-Tween buffer for 10 min. The membrane was then incubated for 1 h with a secondary antibody of goat anti-rabbit IgG, goat anti-mouse IgG, or donkey anti-goat IgG conjugated to horseradish peroxidase. Each target protein level was determined using the immobilon western chemiluminescent HRP substrate (Millipore Corp., Billerica, MA, USA) and Agfa X-ray film (Agfa-Gevaert, Mortsel, Belgium). Incubation with monoclonal mouse β-actin antibody was also performed for comparative controls.
Periodic acid-Schiff (PAS) staining
For the histological analyses of glomerular tissues and renal tubule tissues, tissue specimens were collected at the end of the experiments and fixed in 10% buffered formaldehyde. The paraffin-embedded tissue specimens were sectioned at 5 μm thickness, deparaffinized, and stained with PAS stain to assess interstitial fibrosis. The stained tissue sections were observed using an optical Axioimager microscope system (Zeiss), and 5 images were taken for each section.
Immunohistochemical staining
For the immunohistochemical analyses of ZO-1, and VEGFR2, this study employed paraffin-embedded, 5 μm thick mouse eye tissue sections. The tissue sections were placed on glass slides, de-paraffinized, and hydrated with xylene and graded alcohol. The sections were then preincubated in a boiled sodium citrate buffer (10 mM sodium citrate, 0.05% Tween 20, pH 6.0) for antigen retrieval. A specific primary antibody against ZO-1 or VEGFR2 was incubated with the tissue sections overnight at room temperature. For ZO-1 visualization, the tissue section was developed with Cy3-conjugated anti-rabbit IgG. In other experiments for visualizing ZO-1, the tissue sections were incubated for 1 h with HRP-conjugated anti-rabbit IgG. Subsequently, the sections were visualized with 3,3′-diaminobenzidine (DAB) to produce a brown staining, and counterstained with hematoxylin. For the localization of VEGFR2, the tissue section was developed with fluorescein isothiocyanate (FITC)-conjugated anti-rabbit IgG. Nuclear staining was performed using 4′,6-diamidino-2-phenylindole (Santa Cruz Biotechnology). The stained tissue sections were examined using an optical Axiomager microscope system, and 5 images were taken for each section.
FITC-dextran perfused retinal flat mounts
Anesthetized mice received 20 mg/ml FITC-dextran dissolved in PBS via an injection into the left ventricle. Eyes were enucleated immediately after the injection of FITC-dextran and fixed in 4% paraformaldehyde solution for 10 min. The retinas were flat-mounted and examined using an optical Axiomager microscope system.
Retinal trypsin digestion assay
The eyes were severed from mice and fixed in 4% paraformaldehyde solution for 24 h, equatorially bisected, followed by the entire removal of the retina. Subsequently, the harvested retinas were washed overnight in distilled water and incubated with 3% trypsin in 0.1 M Tris buffer (pH 7.8) for 1 h at 37°C. Non-vascular tissues were cautiously trimmed, and the vasculature was isolated and air-dried on the slide. The retinal vasculature was stained with hematoxylin and eosin (H&E) for visualization and examined using an optical Axiomager microscope system.
Statistical analysis
The data are presented as mean ± standard error of mean for each treatment group. Statistical analyses were conducted using a Statistical Analysis Systems program (SAS Institute, Cary, NC, USA). One-way analysis of variance was applied to determine the inhibitory effects of diacetylchrysin on renal fibrosis and retinal neovascularization in diabetic mice. Differences among treatment groups were analyzed with Duncan’s multiple-range test and are considered significant at P < 0.05.
Go to :
RESULTS
Inhibition of collagen accumulation by diacetylchrysin
Renal fibrosis is an inevitable consequence due to an excessive accumulation of ECM components that occurs in chronic kidney diseases [14]. This study attempted to determine whether exposure to diacetylchrysin attenuated the diabetes-associated deposition of ECM proteins. Dense collagen fiber deposition was distinctly detected (blue color) in glomerular and tubular tissues of db/db mouse kidneys, evidenced by Masson's trichrome staining (Fig. 3A and B). In contrast, oral administration of 10 mg/kg diacetylchrysin diminished the deposition of collagen fibers, indicating the inhibition of renal fibrosis.
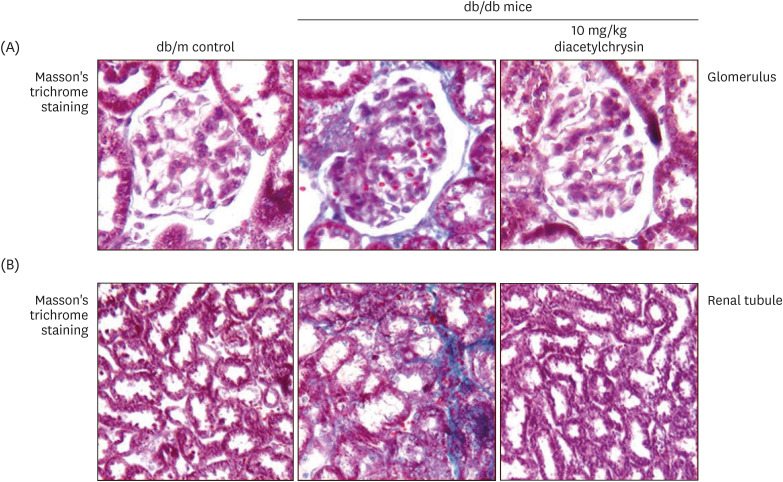 | Fig. 3Effects of diacetylchrysin on formation of collagen fibers in glomeruli (A) and renal tubules (B). The db/db mice were supplemented with 10 mg/kg diacetylchrysin for 10 weeks. The db/m mice were employed as control animals. Glomerular and tubular fibrosis in db/db mouse kidneys was observed by Masson’s trichrome staining (A and B). The collagen fibers were stained in blue. Each photograph is representative of 4 mice. Magnification: 200-fold.
|
Effects of diacetylchrysin on induction of ECM-related enzymes
This study further investigated the mechanism by which diacetylchrysin diminished the deposition of ECM components involved in the structural remodeling of kidneys. The CTGF expression was enhanced in diabetic kidneys, and such induction was markedly suppressed by oral administration of 10 mg/kg diacetylchrysin (Fig. 4A). A similar effect was observed in the increased expression of collagen IV in diabetic kidneys (Fig. 4B). On the other hand, the expression of the ECM-degrading MT1-MMP was markedly reduced in the renal tissues of diabetic mice (Fig. 4C). It was found that diacetylchrysin boosted the MT-1 MMP expression in diabetic kidneys. The TIMP-2 induction up-regulated in renal tissues of diabetic animals was significantly reduced after diacetylchrysin treatment (Fig. 4D). Thus, our results determined that diacetylchrysin attenuated the increased accumulation of ECM components in diabetic kidneys, possibly via inhibiting their excessive production and simultaneously enhancing their degradation.
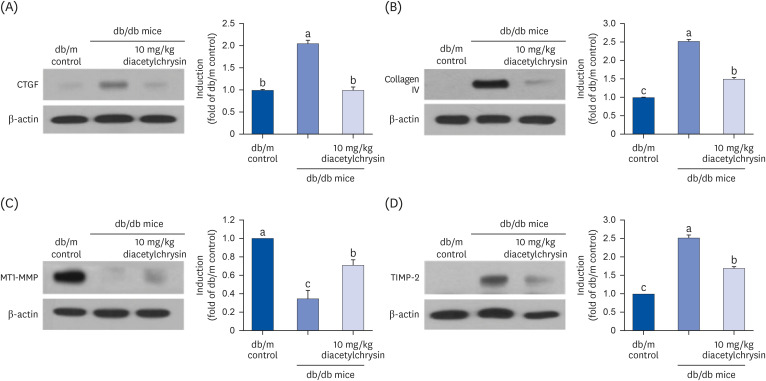 | Fig. 4Effects of diacetylchrysin on the expressions of CTGF (A), collagen IV (B), MT1-MMP (C), and TIMP-2 (D) in db/db mice. The db/db mice were orally treated with 10 mg/kg diacetylchrysin or 10 mg/kg chrysin for 10 weeks. The db/m mice were introduced as control animals. Western blot analysis with tissue extracts was conducted with a primary antibody against CTGF, collagen IV, MT1-MMP, and TIMP-2. β-Actin protein was used as an internal control. The bar graphs (mean ± SEM, n = 3) in the right panel represent quantitative results obtained from a densitometer. Values not sharing a common small letter are significantly different at P < 0.05.CTGF, connective tissue growth factor; MT1-MMP, membrane type 1-matrix metalloproteinase; TIMP-2, tissue inhibitor of metalloproteinase 2; SEM, standard error of mean.
|
Inhibitory effects of diacetylchrysin on glomerular sclerosis and tubular injury
Pathologic alterations in glomeruli contribute to glomerular filtration barrier injury and abnormal protein filtration in diabetic kidneys [2728]. The PAS staining revealed glomerulosclerosis observed in db/db mouse kidneys (arrows), which was ameliorated by 10 mg/kg diacetylchrysin supplementation (Fig. 5A). Furthermore, the oral administration of diacetylchrysin hampered tubular interstitial fibrosis leading to renal tubular injury in db/db mice (Fig. 5B). The protein escape from the glomerular capillary due to glomerular damage causes protein overload on the tubular epithelium and within the tubulointerstitium [28]. In fact, the urinary albumin was markedly elevated in diabetic kidneys, which was diminished after exposure to diacetylchrysin (Fig. 2E).
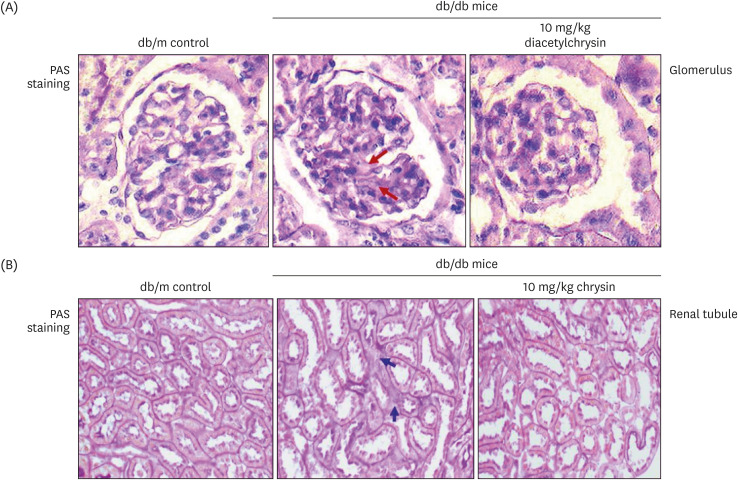 | Fig. 5Inhibition of renal interstitial fibrosis (arrows) in glomeruli (A) and renal tubules (B) of db/db mice by diacetylchrysin. The db/db mice were supplemented with 10 mg/kg diacetylchrysin for 10 weeks. The db/m mice were employed as control animals. Glomerular and tubular tissue sections were stained by PAS reagent and counterstained with hematoxylin (A and B). Each photograph is representative of four mice. Magnification: 200-fold.PAS, Periodic acid-Schiff.
|
Induction of retinal endothelial proteins by diacetylchrysin
The present study examined whether diacetylchrysin ameliorated the cellular expression and distribution of vascular endothelial junction molecules in diabetic retinas. The eye tissue expression of ZO-1, known as the tight junction protein-1, decreased in db/db mice (Fig. 6A). However, such reduction was elevated in the 10 mg/kg diacetylchrysin-challenged diabetic mice. In addition, immunohistochemical staining with red-fluorescent Cy3 confirmed the localization of ZO-1. Weak ZO-1 immunostaining was observed in the neurofiber layer/ganglion cell layer (NFL/GCL) and in the photoreceptor inner segment/outer segment (IS/OS) of the diabetic retina (Fig. 6B). In contrast, the immunostaining of ZO-1 was restored in diabetic retinas exposed to diacetylchrysin. Conversely, the retinal ZO-1 was DAB-stained in diabetic mouse retinas exposed to 10 mg/kg chrysin. The DAB-brown staining of ZO-1 was weakly detected in the NFL/GCL and IS/OS, while ZO-1 was highly localized in the retina of db/db mice treated with10 mg/kg chrysin (Fig. 6C).
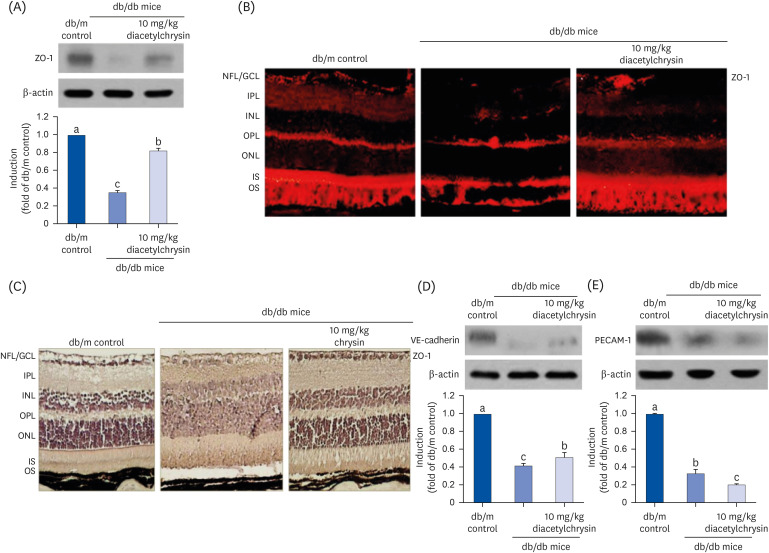 | Fig. 6Effects of diacetylchrysin on the expression of endothelial junction markers. The db/db mice were orally supplemented with 10 mg/kg diacetylchrysin daily for 10 weeks. The db/m mice were introduced as control animals. Mouse retinal tissue extracts were subject to Western blot analysis with a primary antibody against ZO-1 (A), VE-cadherin (D), and PECAM-1 (E). β-Actin protein was used as an internal control. Bar graphs (mean ± SEM, n = 3) in the right panel represent densitometric results of left blot bands. Values not sharing a common small letter differ, P < 0.05. Histological sections of mouse retina were immunohistochemically stained using a primary antibody of ZO-1 and red Cy3-conjugated secondary antibody for visualizing the ZO-1 induction (B). In another experiment with 10 mg/kg chrysin administration, the ZO-1 expression was identified as 3,3′-diaminobenzidine staining (brown) and the sections were counterstained with hematoxylin (C). Each photograph is representative of four mice. Magnification: 200-fold. Retinal layers are labeled as follows: NFL/GCL, IPL, INL, OPL, ONL, and photoreceptor IS/OS.ZO, Zona occluden; VE, vascular endothelial; PECAM, platelet endothelial adhesion molecule; SEM, standard error of mean; NFL, neurofiber layer; GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; IS, inner segment; OS, outer segment.
|
This study further examined whether diacetylchrysin influenced the expression of VE-cadherin and PECAM-1 localized at the adherens junctions in diabetic retinas. The retinal expression of VE-cadherin was drastically reduced in diabetic mice (Fig. 6D), but was significantly enhanced in the db/db mice treated with 10 mg/kg diacetylchrysin. However, the treatment with diacetylchrysin did not restore the expression of the endothelial cell-cell adhesion molecule PECAM-1 diminished in diabetic retinas (Fig. 6E).
Inhibition of retinal neovascularization by diacetylchrysin
Hyperglycemia results in pathological neovascularization and vascular dystrophies in the retina [2429]. This study examined whether oral administration of diacetylchrysin attenuated the expression of proangiogenic VEGF in diabetic retinas. The VEGF expression was induced in diabetic eye tissues, while the VEGF induction returned to levels of db/m control mice after treatment (Fig. 7A). Furthermore, the oral supplementation of diacetylchrysin near-completely inhibited the highly increased VEGFR2 induction seen in diabetic eyes (Fig. 7B). The FITC staining for retinal VEGFR2 was lacking in non-diabetic mice (Fig. 7C), but was apparently induced in the retinal tissues of db/db mice. A heavy green staining denoting VEGFR2 was detected in the GCL, outer nuclear layer (ONL) of the diabetic retina, and OS of the diabetic retina (Fig. 7C). Oral administration of 10 mg/kg chrysin substantially diminished the VEGFR2 staining in the GCL, ONL, and OS, and levels were almost indistinguishable from that of db/m controls (Fig. 7C). These results indicate that both diacetylchrysin and chrysin potentially allay the diabetes-associated retinal neovascularization.
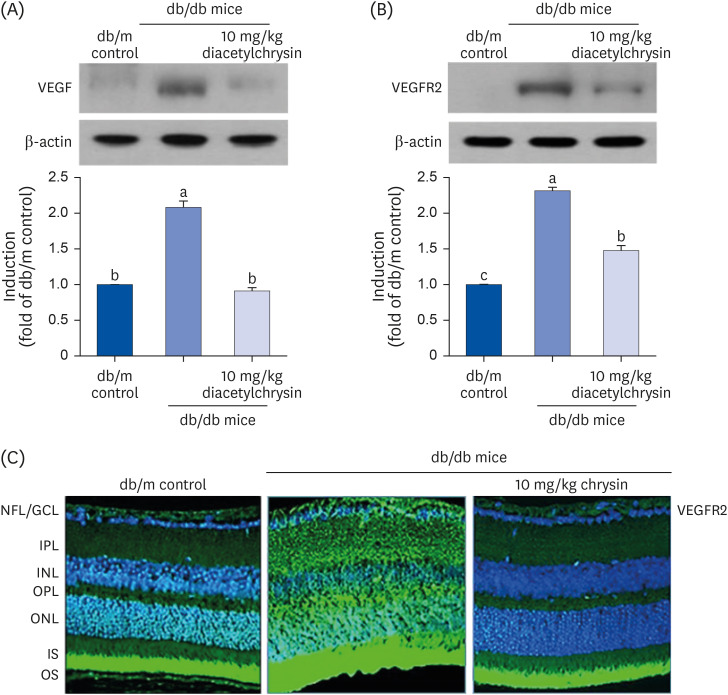 | Fig. 7Inhibition of retinal tissue induction of VEGF (A) and VEGFR2 (B) by diacetylchrysin. The db/db mice were orally administered 10 mg/kg diacetylchrysin daily for 10 weeks. The db/m mice were employed as control animals. Mouse retinal tissue extracts were subject to Western blot analysis with a primary antibody against VEGF and VEGFR2 (A and B, respectively). β-Actin protein was used as an internal control. Bar graphs (mean ± SEM, n = 3) in the right panel represent densitometric results of left blot bands. Values not sharing a common small letter differ, P < 0.05. Immunocytochemical analysis of VEGFR2 in db/db mice treated with 10 mg/kg chrysin was achieved by histological staining of mouse retina sections using a primary antibody of VEGFR2. FITC-conjugated secondary antibody was used for visualizing the VEGFR2, and the nuclear staining was done with 4′,6-diamidino-2-phenylindole (C). Each photograph is representative of 4 mice. Magnification: 200-fold. Retinal layers are labeled as follows: NFL/GCL, IPL, INL, OPL, ONL, and photoreceptor IS/OS.VEGF, vascular endothelial growth factor; VEGFR2, VEGF receptor 2; SEM, standard error of mean; FITC, fluorescein isothiocyanate; NFL, neurofiber layer; GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; IS, inner segment; OS, outer segment.
|
Suppressive effect of diacetylchrysin on retinal vessel leakage
Hyperglycemia stimulates retinal neovascularization with the abnormal formation of new blood vessels, which leads to the leakage and dystrophies of retinal vessels [30]. This study examined whether diacetylchrysin inhibited the acellular capillary formation in db/db mouse retina, as evidenced by retinal trypsin digestion assay and subsequent H&E staining. Acellular capillaries were observed in diabetic retinas (Fig. 8A), which were typically devoid of both pericytes and endothelial cells [17]. Treatment with 10 mg/kg diacetylchrysin resulted in reduced formation of acellular capillaries in the retinas of diabetic mice (Fig. 8A).
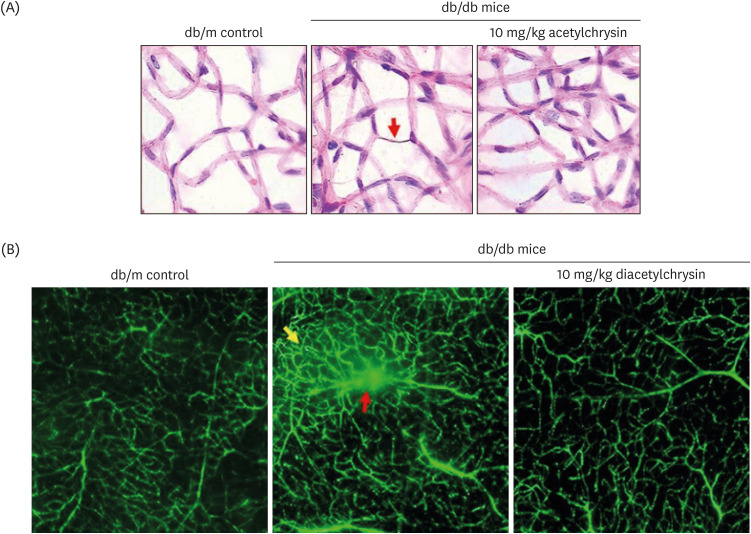 | Fig. 8Inhibition of abnormal neovascularization by diacetylchrysin. The db/db mice were orally supplemented with 10 mg/kg diacetylchrysin daily for 10 weeks. The db/m mice were employed as control animals. To reveal acellular capillaries (red arrow) in db/db mice treated with 10 mg/kg diacetylchrysin, retinal vessels were subjected to H&E staining (A). Magnification: 200-fold. To detect new capillary bed formation (yellow arrow) and vessel leakage (red arrow) by diacetylchrysin exposure (B), typical appearance of retinal capillaries was observed by FITC-dextran-perfused retinal flat-mounts. Retinas were dissected, flat mounted, and observed by confocal microscopy. Magnification: 400-fold. Each photograph is representative of 4 mice.H&E, hematoxylin and eosin; FITC, fluorescein isothiocyanate.
|
To reveal the suppressive effects of diacetylchrysin on vascular leakage in the retinas, the FITC-conjugated dextran perfusion assay was performed by using flat-mounted mouse retina. No vascular abnormality was detected in the retina of db/m mice (Fig. 8B). However, diffuse staining was observed in the retina of db/db mice (red arrow). In addition, there were increased neovascular tufts in the diabetic retina (yellow arrow). Oral administration of diacetylchrysin curtailed the retinal neovascularization with vessel leakage in the db/db mice. Thus, diacetylchrysin has the potential to optimally maintain retinal vasculature via induction of junction proteins in the retinal endothelium (Fig. 6C).
Go to :
DISCUSSION
Eight significant findings were determined from this study. 1) Oral administration of 10 mg/kg diacetylchrysin did not lower fasting blood glucose level and 24 h urine volume in diabetic animals, but the urinary albumin was markedly reduced. 2) Supplementation of diacetylchrysin diminished the deposition of collagen fibers in diabetic glomeruli and tubules. 3) The inductions of CTCF and collagen IV in diabetic kidneys were markedly suppressed by diacetylchrysin treatment. 4) Diacetylchrysin administration enhanced the MT-1 MMP expression reduced in diabetic kidneys, while the TIMP-2 induction was attenuated in renal tissues of diacetylchrysin-challenged diabetic mice. 5) Supplementing diacetylchrysin to diabetic mice ameliorated glomerulosclerosis and tubular interstitial fibrosis which result in renal injuries. 6) Reduced retinal induction of ZO-1, VE-cadherin, and PECAM-1 was observed in db/db mice, whereas retinal tissue levels of ZO-1and VE-cadherin were elevated by supplying diacetylchrysin to the animals. 7) Oral administration of diacetylchrysin hampered the induction of VEGF and VEGFR2 in db/db mice, ultimately retarding the diabetes-associated retinal neovascularization. 8) Orally administered diacetylchrysin curtailed the appearance of acellular capillaries with leaky vessels in db/db mice. Taken together, these results indicate that diacetylchrysin probably inhibited renal fibrosis through inhibition of accumulation of ECM components, and also allayed abnormal retinal neovascularization through increased induction of retinal junction proteins (Fig. 9).
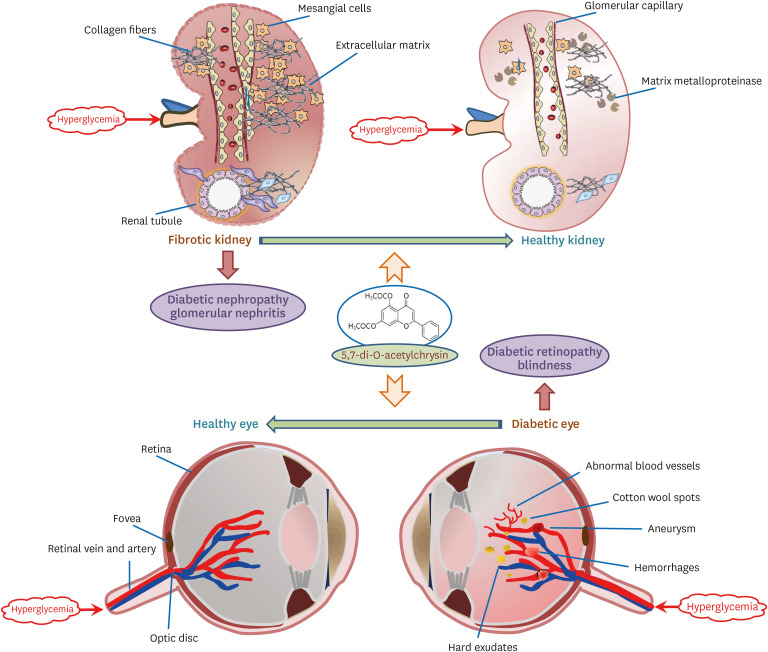
Much effort has focused on developing the management and elucidating experimental therapeutic targets in order to treat diabetic complications. Several signaling sites of oxidative stress are currently being investigated as a baseline therapy for the prevention and treatment of diabetes and its associated complications [22]. The hyperglycemia-induced imbalance between the cellular antioxidant system and reactive oxygen species (ROS) production results in oxidative stress, which contributes to macrovascular and microvascular diabetic complications [31]. Thus, taking antioxidants such as natural products and their derived bioactive compounds has been postulated as possible therapeutic strategies targeting ROS production in diabetic complications [323334]. Since the antioxidant resveratrol effectively lowers the blood glucose level without any side effects, there has been a growing interest in its application for DN and DR, together with the development of new molecular targets [3536]. In addition, the therapeutic potential of targeting signaling pathways of hypoxia-inducible factors has been suggested for the prevention and treatment of associated complications [37]. Our previous studies revealed that the antioxidant chrysin reduced the blood glucose levels and retarded oxidative stress in type 2 diabetes-triggered DN and DR [232438]. However, the water-insoluble chrysin was dissolved in DMSO. Accordingly, the current study was conducted to determine the in vivo preventive effects of diacetylchrysin, which has higher water solubility, on DN and DR. A previous study reported that acetylchrysin inhibits the formation of advanced glycation end products and exhibits anti-inflammatory activity [26]. The current study found that diacetylchrysin antagonized renal fibrosis and abnormal retinal neovascularization in db/db mice, in spite of no blood glucose-lowering effect. It can be assumed that using diacetylchrysin as an adjuvant to conventional antidiabetic therapies is a potentially effective approach to manage diabetic complications.
Although cutting-edge investigations have identified several molecular mechanisms related to the development of DN and DR, the impact of how diabetes injures the kidneys and retina remains unclear, and therapeutic options are limited for these threatening diseases. Inhibition of the sodium-glucose co-transporter (SGLT) 2 has surfaced as a promising antidiabetic strategy, showing renoprotective effects in type 2 diabetic kidneys [39]. The SGLT proteins located in the renal proximal tubules are responsible for glucose reabsorption into the systemic circulation. In addition, incretin-based agents, including glucagon-like peptide 1 receptor agonists and dipeptidyl peptidase 4 inhibitors, effectively lower the blood glucose level and reduce the risk of developing or worsening chronic kidney disease through inhibiting inflammation, oxidative stress, fibrosis, and apoptosis [40]. Severe dysfunction of the glomerular capillary barrier to circulating proteins produces protein overload onto tubular epithelial cells that extend the injury to the tubulointerstitium [41]. The current study revealed that diacetylchrysin inhibited glomerular fibrosis and albuminuria, indicating that this compound well-operated glomerular capillary barrier and tubular function in diabetic kidneys. Conversely, the inhibition of ocular VEGF has emerged as a promising treatment modality for proliferative DR [2142]. However, the intravitreal use of anti-VEGF agents is severely limited due to risk factors for crunch development in proliferative DR [43]. In this study, diacetylchrysin acted as a natural risk-free anti-VEGF agent, inhibiting abnormal capillary formation and neovascularization in the diabetic retina. Similarly, natural compounds of curcumin, the chalcone isoliquiritigenin, the flavonoid deguelin, the isoflavone genistein, and the flavone apigenin have shown promising anti-angiogenic effects in DR and choroidal neovascularization [44]. These natural products may be as efficacious and affordable lead compounds with minimal side effects.
Our previous studies showed that oral administration of chrysin exhibited renoprotective and retinoprotective effects in diabetic mice [2324]. Nevertheless, the bioavailability of chrysin is very low due to the low solubility in water and under physiological conditions [45]. The current study, therefore, attempted to increase the absorption of chrysin and thus augment its functionalization on biological activity and in vivo therapeutic efficacy. Comparative pharmacological efficacy was evaluated using synthetic diacetylchrysin with higher solubility than chrysin. Unlike chrysin [23], diacetylchrysin showed no glucose-lowering effects. However, this synthetic compound reduced urinary albumin excretion in diabetic mice, indicating that diacetylchrysin displayed pro-health effects in the kidneys and the retina. A previous investigation reported that 5,7-diacetylchrysin displays the most potent antitumor activity among a series of 5,7-disubstituted chrysin, 7-monosubstituted chrysin, 5-monosubstituted chrysin derivatives synthesized by alkylation, acetylation, benzoylation, and carboxymethylation [46].
In summary, the current study determined that the highly water-soluble diacetylchrysin antagonized diabetes-associated renal fibrosis and abnormal retinal microvascular abnormalities. Oral administration of diacetylchrysin blocked the formation of collagen fibers in mouse kidneys by reducing the collagen IV formation and enhancing its degradation. Moreover, diacetylchrysin attenuated glomerulosclerosis and tubular interstitial fibrosis leading to renal injury. Furthermore, the administration of diacetylchrysin elevated the levels of VE-cadherin and ZO-1, which are downregulated in the diabetic retina. In addition, this compound inhibited the hyperglycemia-mediated induction of VEGF involved in retinal neovascularization, ultimately blocking the formation of acellular capillaries and the appearance of leaky vessels in retinal vascular beds of diabetic mice. Therefore, diacetylchrysin is a therapeutic compound, antagonizing both renal and retinal damages in animal models of diabetic complications. However, to validate the beneficial effects of diacetylchrysin in DN and DR, clinical studies are warranted to elucidate the role of diacetylchrysin against diabetic complications.
Go to :
 XML Download
XML Download