

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Dopamine (DA) and serotonin (5HT) are representative neurotransmitters associated with various psychiatric disorders, such as schizophrenia, addiction, anxiety, and attention-deficit/hyperactivity disorder [1234]. DA neurons in the ventral tegmental area and 5HT neurons in the raphe nucleus innervate mesocorticolimbic areas, including the prefrontal cortex [5]. These neurotransmitters affect the function of target brain structures via cognitive and affective control [678]; hence, novel agents that modulate DA and 5HT transmission can be used as potential therapeutics for psychiatric disorders. Imbalance of DA and 5HT transmission due to altered expression and reuptake mechanisms via dopamine transporter (DAT) and serotonin transporter (SERT) has been implicated in various psychiatric disorders, such as the attention-deficit/hyperactivity disorder, autism spectrum disorder (ASD), major depressive disorder (MDD), substance use disorder, and schizophrenia [910].
Several species of brown seaweed exert various biological effects, such as antioxidative, memory-improving, and antidepressant effects, that are suggested to be useful for the treatment of neurological and psychiatric disorders [111213]. Brown seaweeds induce these effects by altering the formation of reactive species using mitogen-activated protein kinase, nuclear factor-kappa B, and phosphatidylinositol 3-kinase [11]. Ishige foliacea, a brown seaweed species, improves scopolamine-induced memory impairment via the upregulation of phosphorylated extracellular signal-regulated kinase and cyclic AMP-response element-binding protein expression levels [12]. Brown seaweed also contains substantial amounts of bioactive compounds, such as fucosterol, fucoidan, fucoxanthin, phlorotannins, and alginic acids [14]. Treatment with phlorotannins prevents hydrogen peroxide-induced increase in intracellular reactive oxygen species (ROS) levels, lipid peroxidation, and calcium release, which cause cell death in murine hippocampal HT22 cells [15]. Moreover, treatment with 6-hydroxydopamine causes cell apoptosis with accumulation of intracellular ROS and decrease in mitochondrial membrane potential in PC12 cells; fucoxanthin improves cell damage via the interaction between Kelch-like ECH-associated protein 1 and nuclear factor erythroid-2-related factor 2 [16].
Hizikia fusiformis (HF) is another brown seaweed species that is cultivated and used as a traditional cuisine and medicine, mainly in East Asian countries, such as South China and South Korea. HF exerts antioxidant, anticancer, and anti-inflammatory effects [17]. HF also induces neuroprotective effects by inhibiting inducible nitric oxide synthase expression and nitric oxide formation in BV2 microglial cell line derived from C57BL/6 mice [17]. Previous studies suggest that HF and its active compounds significantly affect the neuronal functions. However, it is unclear whether HF active compounds affect the DA and 5HT systems and their associated neural functions, which are the major targets in psychiatric disorders. Therefore, in this study, we investigated the effects of HF on the expression levels of molecules associated with DA and 5HT transmission in primary neuronal cell cultures from rodent cortex, including the prefrontal cortical region.
MATERIALS AND METHODS
Primary neuronal cell culture
All animal experiments were performed in accordance with the Research Ethics Policy of the Korean Association of Laboratory Animal Science and approved by the Institutional Animal Care and Use Committee of Daegu Catholic University (IACUC-2020-047). This study was performed using a minimal number of animals. Cerebral cortical neurons were obtained from CD-1 mice on embryonic day 14 (Orient-Bio, Inc., Seongnam, Korea) and cultured. After decapitation, the brains were removed from the skull and cortical neurons were dissected. Eight-well chamber slides were coated with 5.0 μg/mL poly-D-lysine (Sigma-Aldrich, St. Louis, MD, USA) for 24 h prior to harvesting cells. Cortical neurons (1.5 × 105 cells/well) were cultured on the coated slides with a neurobasal medium (Gibco, Rockville, MD, USA), consisting of 12% horse serum (Gibco), 0.6% glucose, and 2 mM L-glutamine in an incubator controlled at 37°C with 10% CO2. One day after incubation, the medium was replaced with a fresh neurobasal medium supplemented with 2% B-27 (Gibco).
Extract preparation and treatment
HF extract was prepared with 70% ethanol (EtOH) and the total filtrate was concentrated to dryness in vacuo at 70°C to obtain the EtOH extract [18]. HF extract at various concentrations (0.1, 1.0, and 10.0 μg/mL) was added to the culture cells in neurobasal medium with 0.2% dimethyl sulfoxide (DMSO) for 24 h before fluorescence immunostaining assays. In control preparations, vehicle (VEH) treatment (only neurobasal medium with 0.2% DMSO) was administered for the same duration.
Fluorescence immunostaining
In this study, we used primary antibodies for DA transmission: mouse monoclonal anti-tyrosine hydroxylase (TH; Abcam; catalog #ab129991), rabbit polyclonal anti-DAT (Santa Cruz; catalog #sc-14002), rabbit polyclonal anti-dopamine receptor D1 (DRD1; Santa Cruz; catalog #sc-14001), mouse monoclonal anti-DRD2 (Santa Cruz; catalog #sc-5303), mouse monoclonal anti-DRD3 (Santa Cruz; catalog #sc-136170), rabbit polyclonal anti-DRD4 (Elabscience; catalog #E-AB-31153), and rabbit polyclonal anti-DRD5 (Mybiosource; catalog #MBS2516950) antibodies; those for 5-HT transmission: rabbit polyclonal anti-5HT1A (Elabscience; catalog #E-AB-32950), rabbit polyclonal anti-5HT1B (Abcam; catalog #ab13896), anti-tryptophan hydroxylase (TPH; catalog #E-AB-16937), and mouse monoclonal anti-SERT (Santa Cruz; catalog #sc-33724) antibodies; and those for neuronal structures: mouse monoclonal anti-synaptophysin (SYP; Sigma-Aldrich; catalog #S5768), mouse monoclonal anti-microtubule-associated protein 2 (MAP2; Santa Cruz; catalog #sc-32791), and mouse monoclonal neurofilament-light chain (NF-L; Santa Cruz; catalog #sc-20012) antibodies.
The 24 h after HF extract treatment, the cells were fixed with 4% paraformaldehyde for 2 h and washed thrice with phosphate-buffered saline. Primary antibodies were prepared according to the manuals and used to treat cells for 24 h at 4°C. The 24 h after treatment, cells were incubated with Alexa Fluor 488 goat anti-mouse IgG (Life Technologies; catalog #A-11001) and Alexa Fluor 594 goat anti-rabbit (Life Technologies; catalog #A-11012) secondary antibodies and 1 µg/mL of 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich; catalog #10236270001) for counterstaining of the nuclei for 2 h at room temperature. The cultured cells were then mounted on glass slides with aqua-poly mounts (Polysciences Inc., Warrington, PA, USA). Staining images were captured using a fluorescence microscope (Leica DM2500; Leica Microsystems, Wetzlar, Germany).
Co-expression of DAPI and neuron-specific markers, such as MAP2 (dendritic marker [19]), SYP (synaptic marker [20]), and NF-L (axonal marker [21]), ensured that the cells analyzed for immunostaining were purified neurons (Supplementary Fig. 1).
Data and statistical analyses
Nine (or 18 for TH and TPH) images of approximately 72,230 μm2 (310 × 233 μm) area per image were captured at random culture sites for each treatment condition. Quantification of fluorescent signals in the images was conducted offline using the ImageJ software (version 1.52a; Schneider, CA, USA). Fluorescence intensity was calculated as the number of cells with fluorescent signals divided by the total number of cells stained with DAPI in each image.
Statistical analysis was conducted using JASP (version 0.16.3; JASP Team, The Netherlands). The effects of HF treatment were assessed via Bayesian analysis of variance (ANOVA) followed by a post-hoc pairwise Bayesian t-test. Bayes factor (BF10) between 1.0 and 3.0 indicates weak (anecdotal) evidence, BF10 between 3.0 and 10.0 indicates moderate evidence, and BF10 greater than 10.0 indicates strong evidence for a group difference.
RESULTS
Effects on the expression levels of molecules associated with DA transmission
We first examined the effects of HF extract on the expression levels of DA receptors (DRD1–5), DAT, and TH (Figs. 1 and 2).
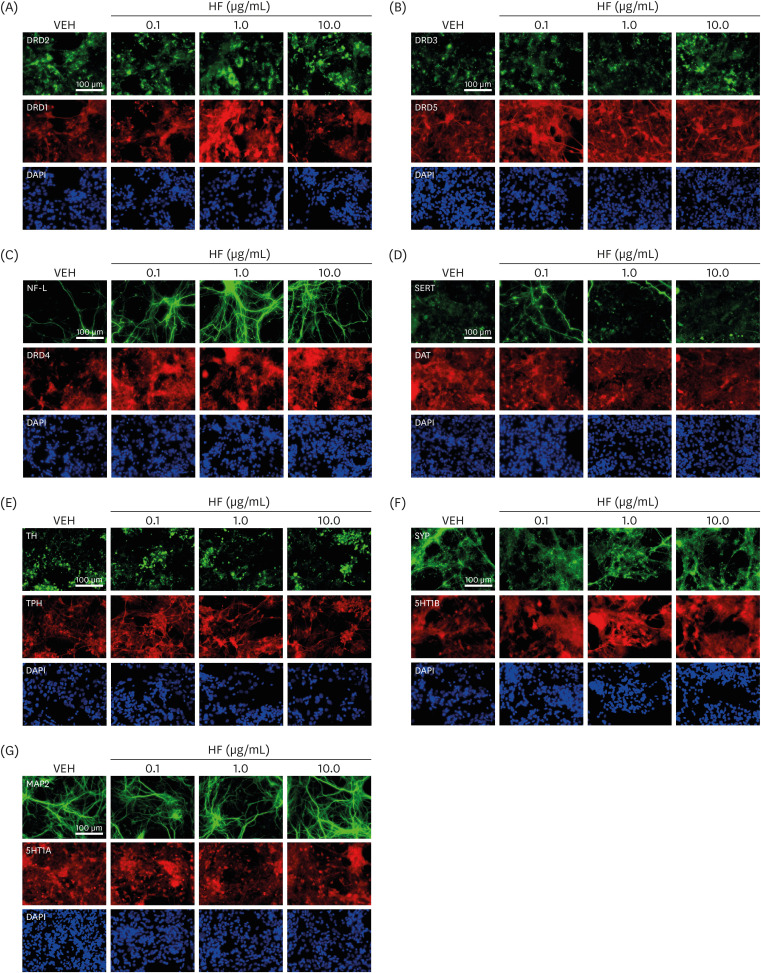
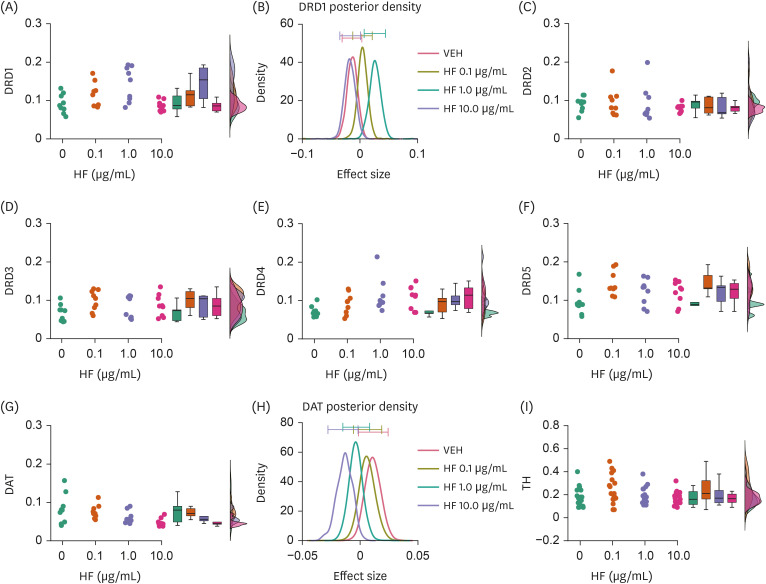
Fig. 1
Representative images of immunofluorescence staining. (A) Images of stained cells for DRD2 (green) and DRD1 (red). Blue color in the images represents fluorescence staining for cell nuclei with DAPI. (B-G) Similar images for (B) DRD3 (green) and DRD5 (red), (C) NF-L (green) and DRD4 (red), (D) SERT (green) and DAT (red), (E) TH (green) and TPH (red), (F) SYP (green) and 5HT1B (red), and (G) MAP2 (green) and 5HT1A (red).
HF, Hizikia fusiformis extract treatment; VEH, vehicle treatment (control); DRD1-5, dopamine D1–5 receptors; DAT, dopamine transporter; DAPI, 4′,6-diamidino-2-phenylindole; NF-L, neurofilament light chain; SERT, serotonin transporter; TH, tyrosine hydroxylase; TPH, tryptophan hydroxylase; SYP, synaptophysin; 5HT1A and 1B, serotonin 1A and 1B receptors; MAP2, microtubule-associated protein 2.
![]()
Fig. 2
Effects of HF extract on the expression levels of molecules associated with DA transmission. (A, B) Raincloud plot illustrating DRD1 expression levels with different doses of HF extract (A) and a graph showing averaged posterior probability distributions (effect size) and 95% confidence intervals (bars on the tops) of it (B). (C-F) Raincloud plots similar to (A), but showing DRD2 (C), DRD3 (D), DRD4 (E), and DRD5 (F) receptor expression levels. (G, H) Raincloud and averaged posterior probability distribution graph similar to (A) and (B), but showing DAT expression. (I) Raincloud plot showing TH expression.
DRD1–5, dopamine D1–5 receptors; HF, Hizikia fusiformis extract treatment; VEH, vehicle treatment (control); DAT, dopamine transporter; TH, tyrosine hydroxylase.
![]()
HF extract affected the expression levels of DRD1 (Bayesian ANOVA, BF10 = 19.987, error% = 0.003; Fig. 2A and B) but not those of other DA receptors (DRD2, BF10 = 0.200, error% = 5.68e-4; DRD3, BF10 = 0.907, error% = 3.16e-5; DRD4, BF10 = 2.356, error% = 9.74e-7; DRD5, BF10 = 2.295, error% = 1.18e-6; Fig. 2C-F). Post-hoc pair-wise Bayesian Student’s t-test revealed that the effects of HF extract on DRD1 expression were dose-dependent, with DRD1 expression being higher at 1.0 μg/mL (BF10 = 5.668, error% = 2.68e-5; Fig. 2A; Table 1), but not at 0.1 or 10.0 μg/mL, than that with VEH treatment (BF10 = 1.024, error% = 0.003 at 0.1 μg/mL; BF10 = 0.461, error% = 0.001 at 10.0 μg/mL; Fig. 2A and B; Table 1). In addition, DAT expression was also found to be lower in cells treated with HF extract at 10.0 μg/mL, but not other doses, compared to those in cells treated with VEH (BF10 = 3.704, error% = 5.20e-7; pair-wise comparisons, BF10 = 0.444, error% = 0.001 at 0.1 μg/mL; BF10 = 0.906, error% = 0.003 at 1.0 μg/mL; BF10 = 3.168, error% = 0.009 at 10.0 μg/mL; Fig. 2G and H; Table 1). No difference was observed in TH expression levels between the HF extract and VEH treatment groups (Fig. 2I).
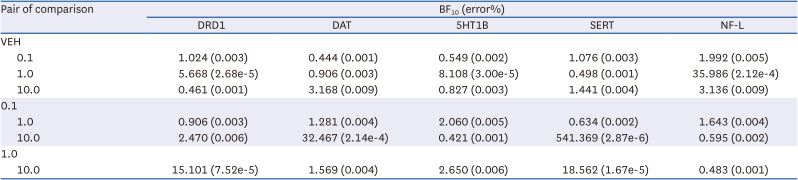
Table 1
A summary of BF10 with post-hoc pair wise comparisons
BF10, Bayes factors; DRD1, dopamine receptor D1; DAT, dopamine transporter; 5HT1B, serotonin 1B receptors; SERT, serotonin transporter; NF-L, neurofilament-light chain.
![]()
These results suggest that HF extract may selectively upregulate D1 receptor expression, along with the potential downregulation of DAT expression.
Effects on the expression levels of molecules associated with 5HT transmission
To determine whether the effects of the HF extract were limited to the DA system, we further investigated the effects of HF extract on the expression levels of molecules associated with 5HT transmission, such as 5HT1A and 5HT1B receptors, SERT, and TPH (Figs. 1 and 3).
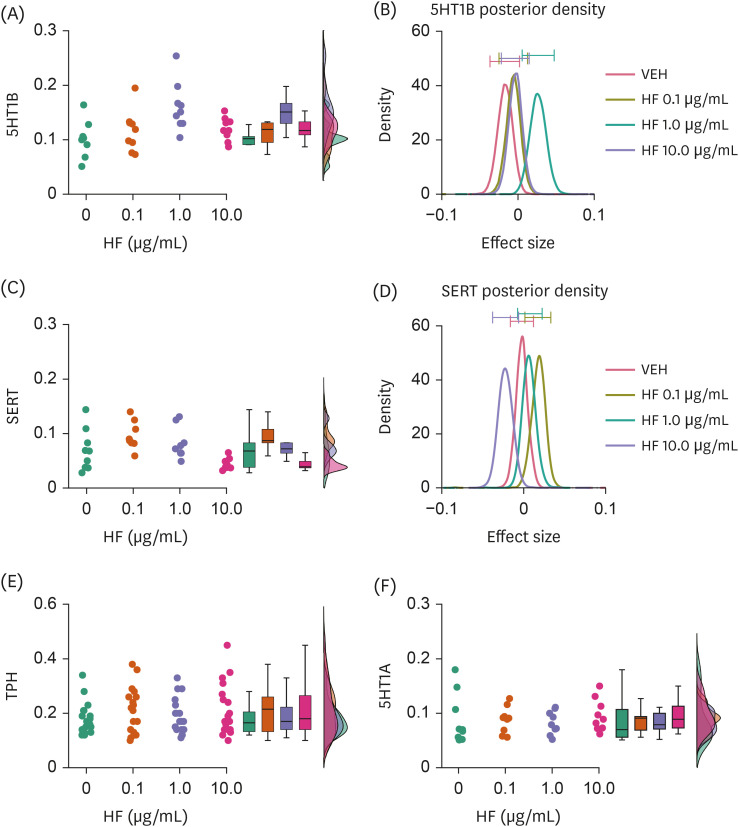
Fig. 3
Effects of HF extract on the expression levels of molecules associated with 5HT transmission. (A, B) Raincloud plot showing 5HT1B expression with different doses of HF extract (A) and a graph showing averaged posterior probability distributions (effect size) and 95% confidence intervals (bars on the tops) of it (B). (C, D) Raincloud and averaged posterior probability distribution graphs similar to (A) and (B), but showing SERT expression. (E, F) Raincloud plots similar to (A, C), but showing TPH (E) and 5HT1A receptor (F) expression levels.
5HT1A and 1B, serotonin 1A and 1B receptors; HF, Hizikia fusiformis extract treatment; VEH, vehicle treatment (control); SERT, serotonin transporter; TPH, tryptophan hydroxylase.
![]()
Interestingly, 5HT1B expression appeared to be higher in cells treated with HF at 1.0 μg/mL, but not other doses, compared to those in cells treated with VEH (BF10 = 6.538, error% = 0.008; pair-wise comparisons, BF10 = 0.549, error% = 0.002 at 0.1 μg/mL; BF10 = 8.108, error% = 3.00e-5 at 1.0 μg/mL; BF10 = 0.827, error% = 0.003 at 10.0 μg/mL; Fig. 3A and B; Table 1). Strong evidence of group differences was also found for SERT expression (BF10 = 15.758, error% = 0.003; Fig. 3C and D). However, pair-wise comparisons yielded only anecdotal evidence of differences between HF extract and VEH treatments at any dose (BF10 = 1.076, error% = 0.003 at 0.1 μg/mL; BF10 = 0.498, error% = 0.001 at 1.0 μg/mL; BF10 = 1.441, error% = 0.004 at 10 μg/mL; Fig. 3C and D; Table 1), suggesting that the source of group difference was primarily nested between different doses of HF extract treatments. No differences were observed in TPH and 5HT1A receptor expression levels between the HF extract and VEH treatment groups (Fig. 3E and F).
These results suggest that the HF extract may selectively upregulate 5HT1B receptor expression, which may be associated with altered SERT expression.
Effects on the expression levels of molecules associated with neuronal structures
Both DA and 5HT play important roles in neuronal growth during embryonic development [22]. Thus, we investigated the expression levels of molecules associated with neuronal structure formation, such as NF-L, SYP, and MAP2 (Figs. 1 and 4).
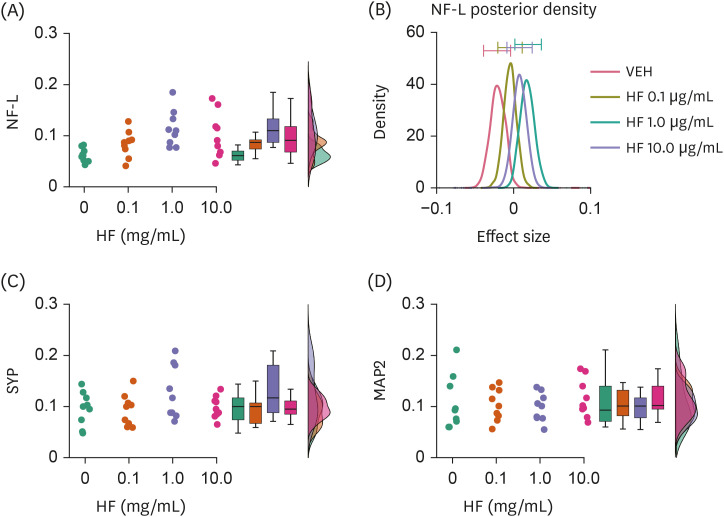
Fig. 4
Effects of HF extract on the expression levels of molecules associated with neural structure formation. (A, B) Raincloud plot showing NF-L expression levels with different doses of HF extract (A) and a graph showing averaged posterior probability distributions (effect size) and 95% confidence intervals (bars on the tops) of it (B). (C, D) Raincloud plots similar to (A), but showing SYP (C) and MAP2 (D) expression levels.
NF-L, neurofilament light chain; HF, Hizikia fusiformis extract treatment; VEH, vehicle treatment (control); SYP, synaptophysin; MAP2, microtubule-associated protein 2.
![]()
HF extract at 1.0 μg/mL, but not other doses, resulted in higher expression of NF-L in cells than VEH (BF10 = 6.053, error% = 0.009; post-hoc comparison, BF10 = 1.992, error% = 0.005 at 0.1 μg/mL; BF10 = 35.986, error% = 2.12E-4 at 1.0 μg/mL; BF10 = 3.136, error% = 0.009 at 10 μg/mL; Fig. 4A and B; Table 1). No differences were observed in SYP and MAP2 expression levels between HF extract and VEH treatment groups (Fig. 4C and D).
These results suggest that the HF extract may affect neuronal structure formation to a limited extent.
DISCUSSION
In this study, we found that the HF extract could affect the expression levels of molecules associated with DA and 5HT transmission and neuronal structural formation in cortical neurons, at least in vtiro, although the effects appear to be relatively limited to specific aspects. In particular, DRD1 and 5HT1B expression levels were upregulated by the HF extract at a moderate dose (1.0 μg/mL), but not at low (0.1 μg/mL) and high (10.0 μg/mL) doses, resulting in inverted U-shape dose-response curves. In contrast, DAT and SERT expression levels were downregulated by the HF extract only at the highest dose. These results can be reconciled by a compensatory mechanism. Therefore, a moderate dose of the HF extract can upregulate D1 and 5HT1B receptor expression levels under a relatively normal range of DA and 5HT release. However, when the dose of HF extract is increased, DAT and SERT expression levels are downregulated, increasing the release of DA and 5HT that downregulate D1 and 5HT1B receptor expression levels, respectively. Consequently, D1 and 5HT1B receptor expression levels return to the normal range after treatment with a high dose of the HF extract.
HF contains various active compounds, including fucosterol, fucoidan, fucoxanthin, phlorotannins, and alginic acids, which exhibit considerable anti-diabetic and anti-inflammatory effects [171823]. Fucosterol [2425], fucoidan [26], and fucoxanthin and its metabolites, fucoxanthinol and amarouciaxanthin [27], have been reported to permeate the blood–brain barrier, thereby affecting the central nervous system. These compounds can be metabolized in the body; in vivo effects of fucosterol in attenuating immobility in the forced swim test have been demonstrated in rodents [28], suggesting that at least in the metabolism of murine species, sufficient amounts of these bioactive compounds can reach the brain to affect its function.
Fucoidan has been shown to prevent the loss of TH-positive DA neurons [29]. Fucoxanthin can function as a selective DA D3/D4 receptor agonist by interacting with H-bonds at Ser196 and Thr115 sites of the D3 receptor as well as at Ser196 and Asp115 sites of the D4 receptor [30]. In contrast, fucosterol increases 5-HT release and the levels of its metabolite, 5-hydroxyindoleacetic acid, in the rat brain [28]. Fucoidan has also been demonstrated to yield in vivo effects similar to those of ondansetron, a serotonin 5HT3 receptor antagonist [31]. Moreover, phlorotannins have recently been identified as monoamine oxidase inhibitors and DA receptor agonists [32]. Collectively, the effects of HF extract on DA and 5HT systems in the current study are not surprising, as HF includes several active compounds that can potently modulate DA and 5HT transmission. However, our study has a major limitation. As the HF extract contains various active compounds, the specific compounds contributing to the observed effects as well as the mechanisms underlying these effects remain unknown. We aim to address this limitation in our future study and examine the in vivo effects of the HF extract on DA and 5HT systems.
Many psychiatric disorders have been reported to show DA and 5HT deficits, with alterations in DA and 5HT reuptake via DAT and SERT, respectively [910]. Positron emission tomography imaging studies in subjects have reported higher limbic and striatal DAT availability in subjects with schizophrenia [33] and cortical DAT availability in subjects with ASD [34], but lower striatal DAT availability in subjects with MDD [35]. Given that the HF extract downregulates DAT expression, it may be used as a therapeutic agent for psychiatric conditions, such as schizophrenia and ASD, but not MDD. Low SERT availability has been reported in patients with MDD [10]. Moreover, 5HT1B agonists have been suggested to have antidepressant effects [37]; therefore, the HF extract may exert effects antagonistic to those of antidepressants.
Our study has demonstrated that the HF extract upregulates the expression of NF-L, which contributes to the structural stability of neurons and is used as an axonal marker of neuronal damage [38], which suggests that the HF extract can impact neuronal growth during neurodevelopment. HF extract effects on molecular expression associated with neuronal structure formation may be achieved directly or indirectly via alterations in DA and 5HT transmission, as DA and 5HT are associated with neuronal development, including neurite outgrowth. For instance, D1 agonists promote the transition of cerebral cortical neurons under development from the G1 to S phase via the downregulation of cyclin D and P27 levels and upregulation of Raf-1 phosphorylation levels in cerebral cortical neurons [38]. Moreover, D1 receptor stimulation in cerebral cortical neurons induces MAP2 phosphorylation via protein kinase A activation [39]. Similarly, 5HT1B receptor agonist also induce the elongation of neurites in thalamic neurons [40].
Accumulating evidence suggests that neurofilaments interact with various receptor subunits to regulate the surface expression of receptors. Short hairpin RNAs targeting DRD1 induce axon degeneration by decreasing NF-L expression in the corpus callosum [41]. A direct interaction occurs between NF-L and the NMDA receptor subunit, NR1 [42], and NF-M, but not NF-L, appears to regulate the surface expression of DRD1 [43]. A recent study reported that the cerebrospinal fluid NF-L levels were positively correlated with striatal DAT expression levels [44]. These findings are somewhat inconsistent with the current findings that the DRD1 and NF-L levels are upregulated and DAT levels are downregulated after HF extract treatment, suggesting that the effects of the HF extract on DA and NF-L expression levels may be mediated via independent molecular processes.
Our study also revealed the upregulation of 5HT1B receptor expression levels and downregulation of SERT expression levels. Notably, 5HT1B receptors are abundant on presynaptic terminals [45] and work as autoreceptors to reduce 5HT synthesis and release upon stimulation [46]. Given that SERT also plays a critical role in the regulation of 5HT release via re-uptake of 5HT molecules, there may be an inverse relationship between autoreceptor and reuptake mechanisms to maintain 5HT release. In contrast, one study has demonstrated that genetic manipulations to increase or decrease 5HT1B receptor expression levels enhance or attenuate SERT functions, respectively [47]. Upregulation of the 5HT1B receptor expression levels and downregulation of SERT expression levels with HF extract treatment in this study are also somewhat inconsistent with the previous report; therefore, further investigations are necessary to determine the specific effects of the HF extract on the 5HT system.
In conclusion, HF extract selectively upregulated DA and 5HT receptor expression levels and downregulated DAT and SERT expression levels in cerebral cortical neurons. HF extract also affected some aspects of neuronal growth, either directly or indirectly, through the modulation of DA and 5HT transmission. These results suggest that HF is a potential modulator of DA and 5HT systems that can be applied for the therapeutic treatment of psychiatric disorders, such as drug addiction [48] and MDD [3649], which have been suggested to be associated with the altered expression levels of D1 and 5HT1B receptors, DAT, and SERT.
 XML Download
XML Download