

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Systemic chronic inflammation is strongly linked to the development of diabetes, cardiovascular diseases, neurodegenerative diseases, and multiple types of cancers [1]. Inflammation-associated chronic diseases are estimated to account for approximately 63% of all-cause mortality worldwide [2]. Nutritional factors, such as the quantity of overall dietary intake and specific nutrients or phytochemicals, have been identified as causes or suppressors of chronic inflammation [2]. Many previous studies have reported that excessive dietary intake or the intake of saturated fatty acids can induce inflammation and result in metabolic stress [3]; however, phytochemicals, which provide color, flavor, smell, texture, and other sense-engaging properties to plants [4], exert health beneficial effects by reducing the incidence of inflammation-associated chronic diseases in humans and various animal models [5].
Curcumin, a phenolic acid isolated from the herb Curcuma longa [5], is a well-known phytochemical with anti-inflammatory effects [6]. It has been found to downregulate the production of diverse proinflammatory cytokines, including tumor necrosis factor (TNF)-α, monocyte chemotactic protein (MCP)-1, and interleukin (IL)-6, by suppressing nuclear factor kappa-B (NF-κB) activation [67]. The anti-inflammatory effects of curcumin contribute to the prevention of metabolic dysregulation [8]. In multiple clinical trials, curcumin supplementation has been found to improve metabolic parameters, including fasting glucose and insulin, serum triacylglycerol (TAG), high-density lipoprotein (HDL), and low-density lipoprotein (LDL) levels [91011].
On the other hand, curcumin has been reported to increase the expression of heat shock protein (HSP) 70 in various cell types, including carcinoma, leukemia, and epithelial cells [121314]. HSP70 is involved in the regulation of cellular responses to inflammatory conditions [151617]. As a ribosome-associated chaperone, HSP70 controls unfolded protein response and endoplasmic reticulum stress [15]. Moreover, as an inhibitor of NF-κB essential modulator, HSP70 suppresses NF-κB signaling activation by inhibiting the phosphorylation of inhibitory κB (IκB) and nuclear translocation of NF-κB, thereby downregulating the expression of proinflammatory cytokines [161718].
Although curcumin is one of the most investigated phytochemicals in terms of its anti-inflammatory effects and underlying mechanisms, whether curcumin upregulates the expression of HSP70 in mature adipocytes and whether the induction of HSP70 is crucial for regulating the anti-inflammatory response of mature adipocytes to curcumin remain unclear. Therefore, this study aimed to assess the role of HSP70 in regulating the effects of curcumin on NF-κB signaling and inflammation using fully differentiated 3T3-L1 adipocytes.
MATERIALS AND METHODS
Cell culture and treatment
3T3-L1 fibroblasts (American Type Culture Collection, Manassas, VA, USA) were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) containing 4,500 mg/L glucose and L-glutamine (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% bovine calf serum (Corning, Corning, NY, USA) and 1% antibiotic antimycotic solution at 37°C in the presence of 5% CO2. Two days after confluence, adipogenic differentiation was initiated using DMEM with 10% fetal bovine serum, 1.7 μM insulin, 1 μM dexamethasone, and 500 μM 3-isobutyl-1-methylxanthine (Sigma-Aldrich). After 2 days, the medium was changed to DMEM with 10% FBS and 1.7 μM insulin. After another 2 days, the medium was changed to DMEM with 10% FBS. When the cells had a spherical shape and were filled with lipid droplets, they were considered fully differentiated.
As previous studies have reported that curcumin concentrations ≤ 100 μM do not induce cell toxicity in adipocytes or adipose-derived mesenchymal stem cells [1920], fully differentiated 3T3-L1 adipocytes were pretreated with 0.1–100 μM curcumin (Cayman, Ann Arbor, MI, USA) dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich) for 24 h. The control was pretreated with DMSO. After curcumin or DMSO pretreatment, the cells were treated with 10 μg/mL lipopolysaccharide (LPS; Sigma-Aldrich) for 30 min or 3 h and/or 1 μM pifithrin-μ (PFT-μ, HSP70 inhibitor; Cayman) for 24 h.
Western blot analysis
Proteins were extracted from 3T3-L1 adipocytes using RIPA lysis buffer containing a protease inhibitor cocktail (Sigma-Aldrich), phenylmethylsulfonyl fluoride, and Na3VO4. The protein concentration was determined using a bicinchoninic acid assay (Sigma-Aldrich). Protein lysates were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. After blocking, the membrane was incubated with rabbit anti-HSP70, rabbit anti-p-IκB, rabbit anti-IκB, and mouse anti-β-actin (Cell Signaling, Danvers, MA, USA). Following this, the membrane was incubated with horseradish peroxidase (HRP)-conjugated anti-mouse or anti-rabbit IgG (Cell Signaling) and imaged using Immobilon Western Chemiluminescent HRP Substrate (Millipore, Burlington, MA, USA). The band density was normalized relative to that of β-actin.
Quantitative real-time polymerase chain reaction (PCR)
Total RNA was extracted from 3T3-L1 cells using TRIzol (Ambion, Austin, TX, USA) and reverse transcribed to cDNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA). Quantitative real-time PCR was performed on a Bio-Rad iQ5 system (Bio-Rad, Hercules, CA, USA) using RT2 SYBR Green Fluor qPCR Mastermix (Qiagen, Valencia, CA, USA). The mRNA levels of Hspa1a, Nfkb1, Il6, Ptx3, Ccl2, and Tnfa were quantified and normalized relative to those of 18S rRNA. Fold changes in gene expression were calculated using the ΔΔCt method. Specific primer sequences used are shown in Table 1.
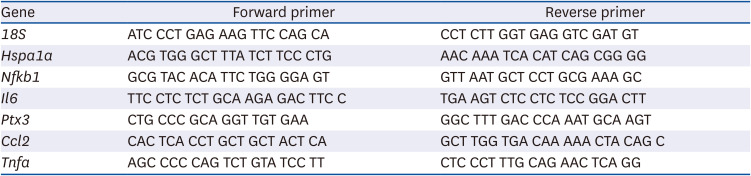
Table 1
Primer sequences used for quantitative real-time polymerase chain reaction
18S, 18S ribosomal RNA; Hspa1a, heat shock protein family A (heat shock protein 70) member 1A; Nfkb1, nuclear factor kappa B subunit 1; Il6, interleukin 6; Ptx3, pentraxin 3 (also known as tumor necrosis factor-inducible gene 14 protein); Ccl2, C-C motif chemokine ligand 2 (also known as monocyte chemoattractant protein-1); Tnfa, tumor necrosis factor α.
![]()
Statistical analysis
One-way or 2-way analysis of variance with Duncan’s post hoc test was performed to determine significant differences induced by treatment with curcumin, LPS, and/or PFT-μ. Data are presented as the means ± standard error of means and were analyzed using SAS 9.4 (SAS Institute Inc., Cary, NC, USA). Differences were considered statistically significant at P < 0.05.
RESULTS
Curcumin upregulates HSP70 expression
To determine whether curcumin induces HSPs, fully differentiated 3T3-L1 adipocytes were treated with 0.1–100 μM curcumin for 24 h. Curcumin upregulated the level of HSP70 in a dose-dependent manner, and its effects on HSP70 induction were significant at 100 μM (Fig. 1A). Although 75 μM curcumin did not have a significant effect on HSP70 induction, more consistent upregulation of HSP70 was noted on treatment with 75 μM curcumin than on treatment with 100 μM curcumin (data not shown). Therefore, 75 μM curcumin was used for treatment in subsequent experiments. On the other hand, curcumin did not affect HSP90 expression (Fig. 1B).
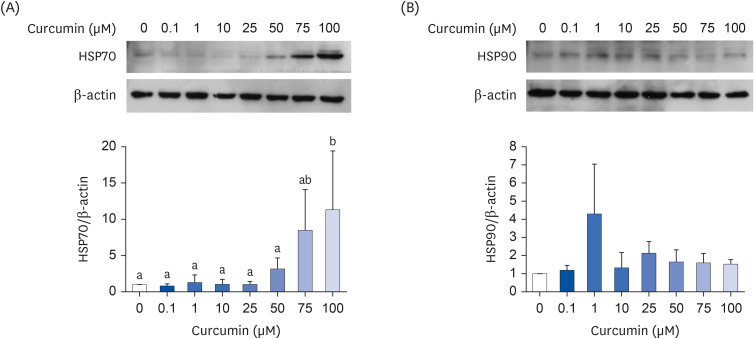
Fig. 1
Effects of curcumin on heat shock protein levels. (A) HSP70; (B) HSP90. Fully differentiated 3T3-L1 adipocytes were treated with curcumin dissolved in dimethyl sulfoxide for 24 h. The protein levels were determined by Western blot with normalization relative to β-actin. Data are presented as means ± standard error of mean (n = 3).
HSP, heat shock protein.
a,bDifferent superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different.
![]()
LPS induces inflammation and reverses curcumin-induced HSP70 expression
When 3T3-L1 adipocytes pretreated with 75 μM curcumin were treated with 10 μg/mL LPS for 30 min, the curcumin-induced upregulation of HSP70 expression decreased (Fig. 2A). LPS treatment increased the phosphorylation (P = 0.011; Fig. 2B) and degradation (P = 0.050; Fig. 2C) of IκB. Curcumin-pretreated cells tended to have a higher level of IκB (P = 0.079; Fig. 2C).
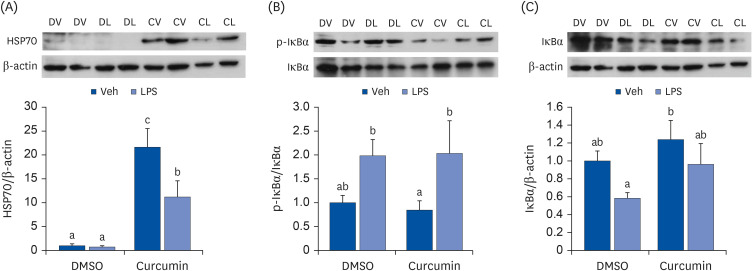
Fig. 2
Effects of curcumin and LPS on HSP70 and NF-κB signaling. (A) HSP70; (B) p-IκBα; (C) IκBα. Fully differentiated 3T3-L1 adipocytes were pretreated with 75 μM of curcumin dissolved in DMSO for 24 h and 10 μg/mL of LPS were treated for 30 min. The protein levels were determined by Western blot with normalization relative to β-actin. Data are presented as means ± standard error of mean (n = 6).
LPS, lipopolysaccharide; HSP, heat shock protein; DMSO, dimethyl sulfoxide; Veh, vehicle; DV, dimethyl sulfoxide with vehicle; DL, dimethyl sulfoxide with lipopolysaccharide; CV, curcumin with vehicle; CL, curcumin with lipopolysaccharide.
a,b,cDifferent superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different.
![]()
Curcumin attenuates LPS-induced inflammation
To determine the effects of curcumin and LPS on NF-κB target gene expression, 3T3-L1 adipocytes pretreated with 75 μM curcumin were treated with 10 μg/mL LPS for 3 h. As the nuclear translocation of NF-κB and transcription of NF-κB target genes take time after IκB phosphorylation and degradation, the cells were treated with LPS for longer in this experiment. Curcumin upregulated the mRNA expression of the HSP70-encoding gene Hspa1a; however, LPS did not suppress this upregulated gene expression. LPS tended to upregulate the mRNA expression of Nfkb1 (P = 0.058), and curcumin pretreatment suppressed the LPS-induced upregulation of NF-κB target genes, including Il6, Ptx3, and Ccl2, at the mRNA level (Fig. 3).
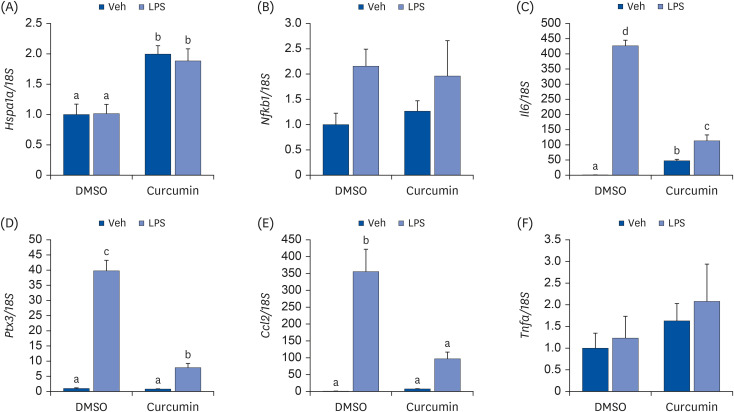
Fig. 3
Effects of curcumin and LPS on pro-inflammatory cytokine expression. (A) Hspa1a; (B) Nfkb1; (C) Il6; (D) Ptx3; (E) Ccl2; (F) Tnfa. Fully differentiated 3T3-L1 adipocytes were pretreated with 75 μM of curcumin dissolved in DMSO for 24 h and 10 μg/mL of LPS were treated for 3 h. The mRNA levels were determined by quantitative real-time polymerase chain reaction with normalization relative to 18S rRNA. Data are presented as means ± standard error of mean (n = 3).
LPS, lipopolysaccharide; DMSO, dimethyl sulfoxide; Veh, vehicle; Hspa1a, heat shock protein family A (heat shock protein 70) member 1A; Nfkb1, nuclear factor kappa B subunit 1; Il6, interleukin 6; Ptx3, pentraxin 3 (also known as tumor necrosis factor-inducible gene 14 protein); Ccl2, C-C motif chemokine ligand 2 (also known as monocyte chemoattractant protein-1); Tnfa, tumor necrosis factor α.
a,b,c,dDifferent superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test.
![]()
HSP70 inhibition tends to exacerbate LPS-induced inflammation
To determine whether HSP70 induction is crucial for the anti-inflammatory effects of curcumin in 3T3-L1 adipocytes, cells were co-treated with 1 μM of the HSP70 inhibitor PFT-μ and 10 μg/mL LPS after pretreatment with 75 μM curcumin. The cells co-treated with PFT-μ and LPS tended to have higher mRNA expression levels of Il6, Ptx3, Ccl2, and Tnfa than those treated with LPS only. However, no effect of PFT-μ was noted in the curcumin-pretreated cells (Fig. 4).
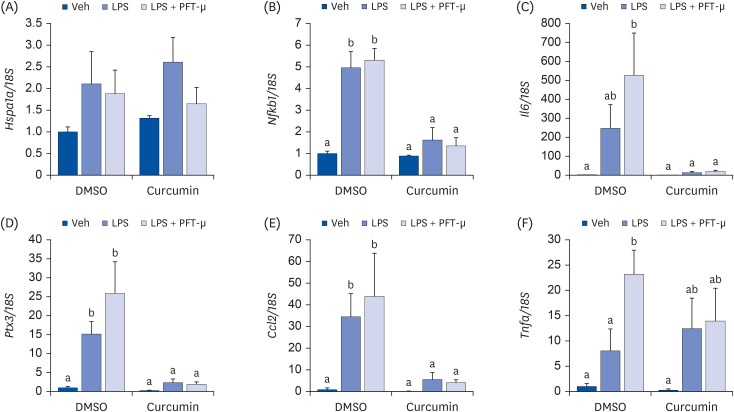
Fig. 4
Effects of HSP70 inhibition on pro-inflammatory cytokine expression in 3T3-L1 cells pretreated with curcumin. (A) Hspa1a; (B) Nfkb1; (C) Il6; (D) Ptx3; (E) Ccl2; (F) Tnfa. Fully differentiated 3T3-L1 adipocytes were pretreated with 75 μM of curcumin dissolved in DMSO for 24 h, and 1 μM of PFT-μ for 24 h and 10 μg/mL of LPS for 3 h were treated. The mRNA levels were determined by quantitative real-time polymerase chain reaction with normalization relative to 18S rRNA. Data are presented as means ± standard error of mean (n = 3).
HSP, heat shock protein; LPS, lipopolysaccharide; DMSO, dimethyl sulfoxide; Veh, vehicle; PFT-μ, pifithrin-μ; Hspa1a, heat shock protein family A (heat shock protein 70) member 1A; Nfkb1, nuclear factor kappa B subunit 1; Il6, interleukin 6; Ptx3, pentraxin 3 (also known as tumor necrosis factor-inducible gene 14 protein); Ccl2, C-C motif chemokine ligand 2 (also known as monocyte chemoattractant protein-1); Tnfa, tumor necrosis factor α.
a,bDifferent superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different.
![]()
As it is possible that the effects of curcumin were too strong and masked the effects of PFT-μ on HSP70 inhibition, a similar experiment was conducted using a lower concentration of curcumin (7.5 μM). Although the difference was not statistically significant, in the cells pretreated with 7.5 μM curcumin, PFT-μ and LPS co-treatment resulted in slightly higher mRNA expression levels of Il6 and Ptx3 than LPS treatment only (Fig. 5).

Fig. 5
Effects of HSP70 inhibition on pro-inflammatory cytokine expression in 3T3-L1 cells pretreated with lower concentration of curcumin. (A) Hspa1a; (B) Nfkb1; (C) Il6; (D) Ptx3; (E) Ccl2; (F) Tnfa. Fully differentiated 3T3-L1 adipocytes were pretreated with 7.5 μM of curcumin dissolved in DMSO for 24 h, and 1 μM of PFT-μ for 24 h and 10 μg/mL of LPS for 3 h were treated. The mRNA levels were determined by quantitative real-time polymerase chain reaction with normalization relative to 18S rRNA. Data are presented as means ± standard error of mean (n = 3).
HSP, heat shock protein; LPS, lipopolysaccharide; DMSO, dimethyl sulfoxide; Veh, vehicle; PFT-μ, pifithrin-μ; Hspa1a, heat shock protein family A (heat shock protein 70) member 1A; Nfkb1, nuclear factor kappa B subunit 1; Il6, interleukin 6; Ptx3, pentraxin 3 (also known as tumor necrosis factor-inducible gene 14 protein); Ccl2, C-C motif chemokine ligand 2 (also known as monocyte chemoattractant protein-1); Tnfa, tumor necrosis factor α.
a,b,cDifferent superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different.
![]()
DISCUSSION
In this study, curcumin was found to suppress the expression of LPS-induced proinflammatory cytokines in fully differentiated 3T3-L1 adipocytes. The anti-inflammatory effects of curcumin have been reported in various experimental models and humans [72122232425]. In 3T3-L1 murine adipocytes, curcumin suppressed the TNF-α-induced upregulation of Il1b, Il6, and Tnfa mRNA expression [7]. In diet-induced or genetically obese mice, curcumin reduced macrophage infiltration in the white adipose tissue and NF-κB activity in the liver [2122]. In an atherosclerotic mouse model, curcumin administration alleviated atherosclerotic lesions; lowered plasma cholesterol, TAG, and LDL levels; and increased the HDL level [23]. In prediabetic or type 2 diabetic patients, curcumin intervention lowered fasting blood glucose, glycated hemoglobin, serum nonesterified fatty acid, and serum TAG levels and improved pancreatic β-cell function [2425]. These data indicate that the anti-inflammatory effects of curcumin can lower the risk of atherosclerosis and insulin resistance.
Curcumin specifically upregulated HSP70 expression at both mRNA and protein levels in 3T3-L1 adipocytes. The expression level of HSP70 was found to be negatively correlated with the degree of inflammation [26], and HSP70 induction by drugs ameliorated symptoms of inflammatory diseases in animal models [2728]. Patients with steatohepatitis and fibrosis had lower levels of HSP70 and HSF1, a transcription factor of HSP70, in the visceral adipose tissue and liver than those with steatosis [26]. Geranylgeranylacetone, an HSP70-inducing drug, could recover the HSP70 level and HSF1 induction and improve the glucose clearance rate in diabetic monkeys [27]. In addition, 17-allylamino-demethoxygeldanamycin, another HSP70-inducing drug, increased neuronal survival and reduced brain edema in rats with brain injury [28].
Although LPS treatment for 30 min downregulated the protein expression level of HSP70, LPS treatment for 3 h did not change the mRNA expression level of Hspa1a. As HSP70 protein levels were found to be positively correlated with Hspa1a mRNA levels in multiple studies that assessed HSP70 expression at both protein and mRNA levels [2930], this inconsistency may be attributed to the difference in treatment times. LPS treatment for more than 60 min was found to upregulate HSP70 expression in various in vitro models [313233]. LPS activated HSF1 by phosphorylation within 5 min of treatment; this effect persisted for 30 min and returned to baseline by 60 min [33].
To determine the role of HSP70 inhibition in the anti-inflammatory response of 3T3-L1 adipocytes to curcumin, cells were co-treated with PFT-μ and LPS after curcumin pretreatment. PFT-μ is known to inhibit HSP70 by disrupting the association between HSP70 and its substrate proteins [34]. To determine the optimal concentration, curcumin-pretreated 3T3-L1 adipocytes were treated with 0.1–200 μM PFT-μ (Supplementary Fig. 1). Compared with other concentrations, 1 μM PFT-μ slightly upregulated the mRNA expression of Hspa1a (albeit not significantly so) and significantly augmented the LPS-induced mRNA upregulation of Ccl2. Therefore, 1 μM PFT-μ was used for HSP70 inhibition in subsequent experiments.
When 3T3-L1 adipocytes were not pretreated with curcumin or were pretreated with a very low concentration of curcumin (7.5 μM), treatment with 1 μM PFT-μ tended to augment the LPS-induced upregulation of proinflammatory cytokine-encoding genes, including Il6, Ptx3, and Ccl2; however, it did not alter the mRNA expression of Hspa1a. Previous studies have reported inconsistent data about the effects of PFT-μ on HSP70 expression [3536]. The expression of HSP70 was upregulated by 40 μM PFT-μ [35] but not by 8 μM PFT-μ [36]; however, both concentrations of PFT-μ significantly increased cell apoptosis, which is mediated by HSP70 inhibition. These results suggest that a low concentration of PFT-μ is enough to inhibit HSP70.
Although HSP70 inhibition by PFT-μ did not significantly reverse the anti-inflammatory effects of curcumin (75 μM) in this study, previous studies have reported that the genetic suppression of Hspa1a or Hsf1 could reduce the anti-inflammatory effects of other phytochemicals. Celastrol, a triterpenoid quinone methide present in thunder god vine root extracts, could upregulate the expression level of HSP70 in a time- and dose-dependent manner [37], while Hspa1a siRNA could reverse the celastrol-induced downregulation of NF-κB p65 DNA-binding activity and IL-6 secretion in human retinal pigment epithelial cells [38]. Zerumbone, a sesquiterpene present in subtropical ginger, also induced HSP70 expression in a dose-dependent manner [39], and Hsf1 siRNA reversed the zerumbone-induced downregulation of Il1b and Nos2 mRNA expression in RAW 264.7 macrophages [40]. These data indicate that HSP70 induction is crucial for the anti-inflammatory effects of celastrol and zerumbone.
A limitation of this study is that the effect of PFT-μ on the disruption of HSP70 binding to its substrate proteins was not determined. Moreover, to reach a strong conclusion, it would be helpful to assess the expression of the markers at both mRNA and protein levels, to increase the number of replicates of the experiments, or to include a positive control, such as HSP70-inducing drugs. However, despite these limitations, this study provides some novel evidence regarding the necessity of HSP70 for the anti-inflammatory effects of curcumin in mature adipocytes.
In summary, curcumin upregulated the expression of HSP70 and attenuated the LPS-induced upregulation of proinflammatory cytokine-encoding genes in fully differentiated 3T3-L1 adipocytes; however, the inhibition of HSP70 by PFT-μ did not significantly curb the anti-inflammatory effects of curcumin. This implies that HSP70 induction, involved in the regulation of anti-inflammatory responses, may not be the main mechanism by which curcumin exerts its anti-inflammatory effects. However, based on the results of previous studies [3840], it is still possible that the genetic suppression of HSP70 expression could reduce the effects of curcumin. Therefore, additional studies with Hspa1a knockdown or knockout are warranted to clarify the role of HSP70 in the regulation of the anti-inflammatory effects of curcumin.
 XML Download
XML Download