

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Platycodon grandiflorum (PG) has been used in traditional medicines as well as food in East Asian countries such as Korea, Japan, and China [1]. It is effective in various diseases related to oxidative and inflammatory reactions, including respiratory diseases [2]. PG contains several nutrients such as polysaccharides, amino acids, vitamins, and minerals [3]. It also contains substantial amounts of phytochemicals beneficial to health, with saponins being the representative bioactive compounds. In particular, platycodin D is a component with excellent quantitative and qualitative physiological activity [4]. It has recently been reported that extraction of PG root with an additional fermentation process increases the proportion of its active ingredient [56]. In our previous study, we isolated a substance with increased saponin and platycodin D content by PG extraction by adding a hydrolysis process before fermentation. Exposure to this hydrolyzed and fermented Platycodon grandiflorum extract (HFPGE) in an in vitro system resulted in increased macrophage phagocytosis and cytokines such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, CXC motif chemokine ligand (CXCL) 10, granulocyte-colony stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), and monocyte chemoattractant protein (MCP)-1 [7]. Thus, we deduced that the HFPGE showed the potential to act as an effective immunostimulatory agent.
As the first line for host defense, the immune system protects the human body from various harmful substances such as infectious agents, allergens, and cancer cells. The immune system is composed of innate and adaptive immunity, and plays an important role in suppressing the occurrence of diseases [8]. Immunodeficiency results in susceptibility to infectious diseases and tumor development, while over activation of the immune system can lead to inflammatory and autoimmune diseases. Therefore, while strengthening the immune system to help maintain health, the immunity needs to be maintained and coordinated in harmony. While efforts to use functional food ingredients to develop immune enhancing agents are on the rise, there is increasing interest in plant ingredients with broader and greater therapeutic effects, relatively low toxicity, and immunomodulatory effects [9].
Cyclophosphamide (CPA) is used in chemotherapy or as an immunosuppressant because it causes a cytotoxic reaction by the formation of reactive metabolites that alkylate DNA and proteins and generate cross-links, and is fatal to proliferating organ tissues [101112]. Administration of CPA to experimental animals causes a rapid change in helper T (Th)1/Th2 bias, resulting in immunosuppression [13]. CPA inhibits the differentiation of T cells, and thus the amount of cytokines secreted from T cells is also reduced [1415]. Therefore, CPA is used experimentally to create immunosuppression models. It has been reported that PG extract has an anticancer effect by enhancing immunity in a CPA-induced immunosuppression model [5], and is effective in restoring the antibody secretion ability and the number of immune cells [13]. However, studies on the immune enhancing effect of PG and its mechanism are insufficient. Therefore, this study was undertaken to confirm the immune-enhancing effect by administering HGPGE to CPA-induced immunosuppressed BALB/c mice.
MATERIALS AND METHODS
Materials
The materials used in this study were purchased from the indicated suppliers: RPMI-1640, fetal bovine serum (FBS), penicillin/streptomycin, and phosphate buffered saline (PBS) from Welgene Inc. (Gyeongsan, Korea); CPA, concanavalin A (ConA), lipopolysaccharide (LPS), and red blood cell lysis buffer from Sigma-Aldrich Co. (St. Louis, MO, USA); enzyme-linked immunosorbent assay (ELISA) kits for immunoglobulin (Ig) A, IgG, IgM, IL-2, IL-4, IL-8, IL-12, interferon (IFN)-γ, TNF-α, and transforming growth factor (TGF)-β from R&D Systems (Minneapolis, MN, USA).
Preparation of HFPGE
HFPGE used in the experiment was manufactured by World Food Services Co. Ltd. (Gangneung, Korea) according to the methods described previously [7]. The HFPGE obtained contained 1.954 ± 0.085 mg (ginsenoside-Re equivalents)/g crude saponin and 1.170 ± 0.013 mg/g platycodin D.
Animals
All animal experiment reported herein were conducted according to the institutional guidelines for the care and use of laboratory animals. All protocols involving animal experiments were approved by the Institutional Animal Care and Use Committee of Hallym University (Hallym 2021-19).
Male, 5-weeks-old BALB/c mice were purchased from Dooyeol Biotech Co. Ltd. (Seoul, Korea). The mice were maintained at the animal research facility of Hallym University in specific pathogen free, controlled conditions at 23 ± 3°C temperature, 50 ± 10% relative humidity, and 12-h light/dark cycle. The mice had free access to a commercial rodent diet (Cargil Agri Purina, Inc., Seongnam, Korea) and tap water.
Experimental design and treatment
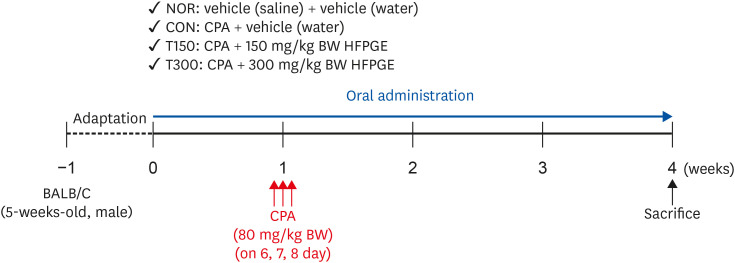
After 1 week acclimation, the BALB/c mice were randomly divided to 4 groups (n = 10 per group) as follows: 1) normal control group (NOR), 2) control group (CON), 3) 150 mg/kg body weight (BW)/day HFPGE-treated group (T150), and 4) 300 mg/kg BW/day HFPGE-treated group (T300). The experimental period for all animals was 4 weeks. The mice in the T150 and T300 groups were orally administered HFPGE dissolved in sterile water once daily for 4 weeks. The mice in the NOR and CON groups were orally administered an equal volume of sterile water as a vehicle. To induce immunosuppression, mice in the CON, T150, and T300 groups were intraperitoneally injected with CPA at a dose of 80 mg/kg BW/day on day 6, 7, and 8, respectively. Simultaneously, mice in the NOR group were intraperitoneally injected with the same volume of physiological saline as a vehicle. After 4 weeks of administering HFPGE, the mice were anesthetized with tribromoethanol diluted in tertiary amyl alcohol, after which blood was collected from the orbital vein. The mice were euthanized by cervical dislocation and the spleen was excised immediately, rinsed, and processed for isolation of splenocytes (Fig. 1).
Fig. 1
Experimental design to examine the effect of HFPGE on immune-enhancing activity in CPA-treated BALB/c mice.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; CPA, cyclophosphamide; NOR, normal control group (injected with vehicle + treated with vehicle); CON, control group (injected with CPA + treated with vehicle); T150, 150 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 150 mg of HFPGE/kg BW/day); T300, 300 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 300 mg of HFPGE/kg BW/day); BW, body weight.
Determination of Igs and cytokines in serum
Serum was obtained from the blood by centrifugation at 1,000 ×g at 4°C for 20 min. The levels of IgA, IgG, IgM, IL-8, IL-12, TNF-α, and TGF-β were measured in the sera using the relevant ELISA kits, according to the manufacturer’s instructions.
Splenocytes isolation
The harvested spleen was chopped in RPMI-1640, passed through a 40 µm nylon cell strainer, and centrifuged at 200 ×g for 10 min. Erythrocytes in the cell mixture were lysed with red blood cell lysis buffer, and the remaining cells were subsequently centrifuged at 200 ×g for 10 min followed by washing in PBS. The resultant splenocytes were resuspended in RPMI-1640 supplemented with 100 mL/L fetal bovine serum, 100,000 U/L penicillin, and 100 mg/L streptomycin, and counted using the Luna-II™ Automated Cell Counter (Logos Biosystems Inc., Anyang, Korea).
Determination of splenocytes proliferation
Fresh splenocytes were plated at a density of 1 × 105 cells/well in a 24-well plate. After 24 h, cells were stimulated with T cell mitogen (ConA) at 5 µg/mL and B cell mitogen (LPS) at 100 µg/mL, and incubated for 48 h. Cell proliferation was measured using a CellTiter 96® Aqueous One Solution Cell Proliferation Assay kit (Promega, Madison, WI, USA), according to the manufacturer’s instructions.
Determination of cytokines in splenocytes
Fresh splenocytes were plated at a density of 2.5 × 105 cells/well in a 48-well plate. After 24 h, cells were stimulated with ConA at 5 µg/mL and LPS at 100 µg/mL, and incubated for further 48 h. The 48 h-conditioned media were collected and the levels of IL-2, IL-4, IL-8, IL-12, IFN-γ, TNF-α, and TGF-β were measured using the relevant ELISA kits, according to the manufacturer’s instructions.
Statistical analysis
All data are expressed as the mean ± SEM (n = 10). All statistical analyses were performed using the Statistical Analysis System for Windows version 9.4 (SAS Institute, Cary, NC, USA). Significant differences between groups were analyzed by Analysis of variance followed by Duncan’s comparison test (P < 0.05).
RESULTS
HFGPE administration does not affect the BW gain in CPA-induced immunosuppressed BALB/c mice
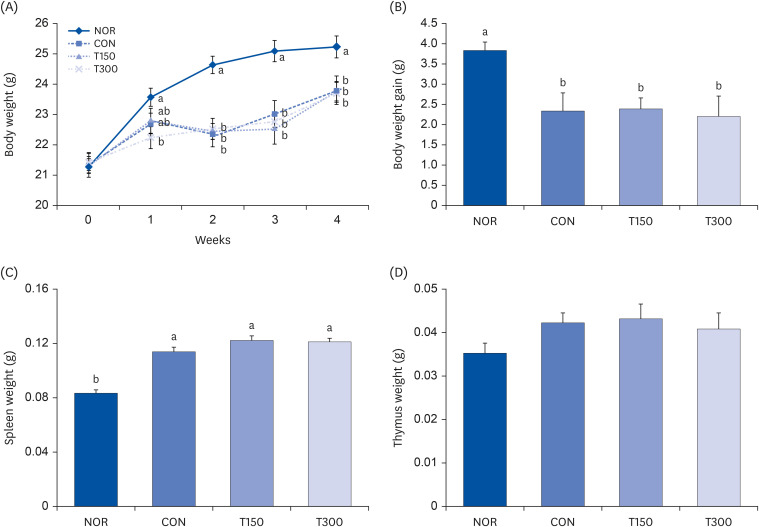
In this experiment, the effect of HFPGE treatment on BW gain was investigated in CPA-induced immunosuppressed BALB/c mice. After completion of the 4-week experimental period, the BW gain of the CON group was significantly reduced compared to the NOR group. There was no significant difference in BW gain among the groups with CPA-induced immunosuppression (Fig. 2A and B). The weight of the spleen was significantly increased by CPA treatment compared to NOR animals, which were not overcome by HFPGE treatment (Fig. 2C). No significant difference was obtained in the thymus weight between all experimental groups (Fig. 2D).
Fig. 2
Effect of HFPGE on BW and spleen and thymus weight in CPA-treated BALB/c mice.
BALB/c mice were administered HFPGE (150 or 300 mg/kg BW/day) for 4 weeks. To induce immunosuppression, the mice were intraperitoneally injected with CPA (80 mg/kg BW/day) on day 6, 7, and 8. The BW was measured, and the BW gain was calculated. After 4-weeks HFPGE administration, all mice were sacrificed, and the spleen and thymus were resected from mice. (A) BW changes; (B) BW gain; (C) Spleen weight; (D) Thymus weight. Values are expressed as the mean ± SEM (n = 10). Means without a common letter differ significantly at P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; CPA, cyclophosphamide; BW, body weight; NOR, normal control group (injected with vehicle + treated with vehicle); CON, control group (injected with CPA + treated with vehicle); T150, 150 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 150 mg of HFPGE/kg BW/day); T300, 300 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 300 mg of HFPGE/kg BW/day).
HFGPE administration increases the serum Ig levels in CPA-induced immunosuppressed BALB/c mice
In order to confirm the effect of HFPGE administration on the immune activity of mice, the levels of Ig, an immune-related marker in the sera, were analyzed. IgA was significantly decreased by CPA treatment, but increased significantly in a dose-dependent manner by HFPGE administration. Serum IgA concentration increased by 3.4% in the T150 group and 4.6% in the T300 group compared to NOR group. IgG was decreased by CPA treatment and increased significantly compared to the CON group subsequent to HFPGE administration. However, even at 300 mg/kg BW HFPGE, the level of IgG did not recover to the NOR group. IgM tended to decrease by CPA treatment, and was increased to the same level as NOR group by HFPGE treatment. IgM concentration was not statistically significant between NOR and CON, but was statistically significant between CON and HFPGE administration groups, with no dose-dependent difference between the T150 and T300 groups (Table 1). Taken together, these results indicate that in the CPA-induced immunosuppressed model, exposure to HFPGE effectively increases Ig secretion, with some differences depending on the xlink:type of Ig.
Table 1
Effect of HFPGE administration on serum Ig levels in CPA-treated BALB/c mice
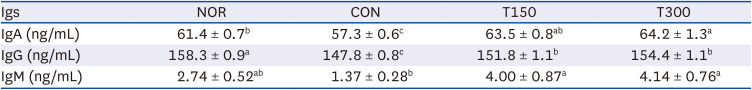
Values are expressed as the mean ± SEM (n = 10). Means without a common letter differ significantly at P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; Ig, immunoglobulin; CPA, cyclophosphamide; NOR, normal control group (injected with vehicle + treated with vehicle); CON, control group (injected with CPA + treated with vehicle); T150, 150 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 150 mg of HFPGE/kg BW/day); T300, 300 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 300 mg of HFPGE/kg BW/day); BW, body weight.
HFGPE administration increases the serum cytokine levels in CPA-induced immunosuppressed BALB/c mice
To investigate the effect of HFPGE administration on adaptive immunity, the serum concentrations of IL-12 and TNF-α (cytokines acting on cell-mediated immunity) and IL-8 and TGF-β (cytokines acting on humoral immunity) were analyzed. A decrease was obtained in all cytokine levels after CPA treatment, but levels were increased subsequent to HFPGE administration. Except for IL-8, the other three cytokines showed significantly higher values than the NOR group after HFPGE administration. Compared to the CON group, the T300 group increased IL-12, TNF-α, IL-8, and TGF-β by 255.9%, 124.5%, 15.4%, and 87.0%, respectively (Table 2). These results suggest that exposure to HFPGE increases the secretion of cytokines reduced by CPA treatment, and has the potential to contribute to the enhancement of cytokine secretion related to cell-mediated immunity in particular.
Table 2
Effect of HFPGE administration on serum cytokine levels in CPA-treated BALB/c mice
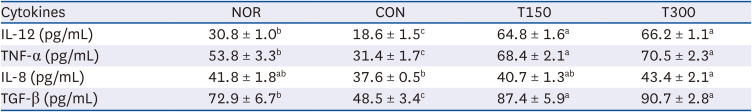
Values are expressed as the mean ± SEM (n = 10). Means without a common letter differ significantly at P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; CPA, cyclophosphamide; NOR, normal control group (injected with vehicle + treated with vehicle); CON, control group (injected with CPA + treated with vehicle); T150, 150 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 150 mg of HFPGE/kg BW/day); T300, 300 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 300 mg of HFPGE/kg BW/day); IL, interleukin; TNF, tumor necrosis factor; TGF, transforming growth factor; BW, body weight.
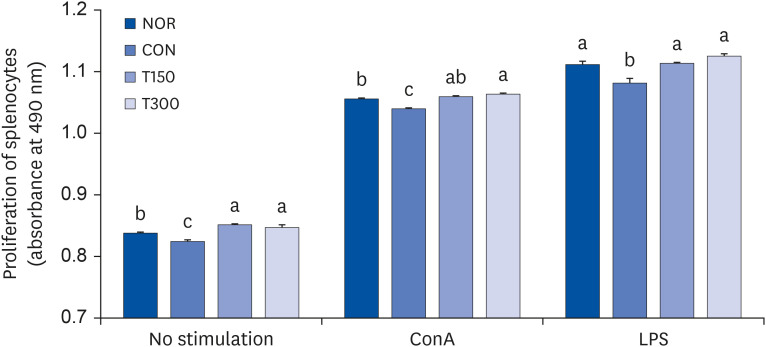
HFGPE administration promotes the proliferation of splenocytes in CPA-induced immunosuppressed BALB/c mice
To investigate the effect of HFPGE administration on splenocyte proliferation, splenocytes isolated from the spleen of BALB/c mice were cultured for 48 h. The proliferation index of splenocytes obtained from the CON group was 0.825 ± 0.002, which was significantly decreased compared to 0.837 ± 0.002 of the NOR group. In the HFPGE-administered group, the splenocyte proliferation index increased significantly in both the T150 and T300 groups as compared to the CON group (Fig. 3). To investigate the effect of HFPGE administration on the proliferation of T cells in splenocytes, the proliferation index was investigated after activating cells with ConA. The proliferation index of splenocytes in the NOR group treated with ConA was determined to be 1.055 ± 0.002, which was significantly increased compared to the NOR group cells not stimulated with the mitogen. Values were significantly decreased in the CON group, and increased significantly in the T150 and T300 groups as compared to the CON group. Similar to stimulation with ConA, the proliferation index was significantly increased when the splenocytes were stimulated with LPS, and the proliferation index decrease after exposure to CPA was significantly increased by the administration of HFPGE (Fig. 3). These results indicate that exposure to HFPGE increases the immune capacity of lymphocytes through proliferation of splenocytes.
Fig. 3
Effect of HFPGE on proliferation of splenocytes from CPA-treated BALB/c mice.
Splenocytes were plated at a density of 1 × 105 cells/well in a 24-well plate. After 24 h, cells were stimulated with 5 µg/mL ConA and 100 µg/mL LPS, and incubated for 48 h. Cell proliferation was measured using a CellTiter 96® AQueous One Solution Cell Proliferation Assay kit. Values are expressed as the mean ± SEM (n = 10). Means without a common letter differ significantly at P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; CPA, cyclophosphamide; ConA, concanavalin A; LPS, lipopolysaccharide; NOR, normal control group (injected with vehicle + treated with vehicle); CON, control group (injected with CPA + treated with vehicle); T150, 150 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 150 mg of HFPGE/kg BW/day); T300, 300 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 300 mg of HFPGE/kg BW/day); BW, body weight.
HFPGE administration increases cytokine production in mitogen-stimulated splenocytes obtained from CPA-induced immunosuppressed BALB/c mice
To determine whether HFPGE administration affects the cytokine production in splenocytes, splenocytes isolated from the CPA-treated BALB/c mice administered with HFPGE were stimulated with mitogen (5 µg/mL ConA). The concentrations of IL-2, IL-12, IFN-γ, and TNF-α were measured in the collected conditioned media after 48 h incubation. Compared to the NOR group, levels of all four cytokines were decreased in the CON group and showed a tendency to increase in a dose-dependent manner by the HFPGE administration. In particular, levels of IL-2, IL-12, IFN-γ, and TNF-α obtained in the T300 group were increased by 132.5%, 133.3%, 30.5%, and 127.3%, respectively, as compared to the CON group (Table 3). Similarly, the levels of IL-4, IL-8, and TGF-β were measured in the collected conditioned media after stimulation of splenocytes with 100 µg/mL LPS. All three cytokines were significantly decreased in the CON group compared to the NOR group, and increased in the HFPGE-administered group. However, unlike the cell-mediated immunity-related cytokines measured after stimulation with ConA, there was no dose-dependent increase by HFPGE. Compared to the CON group, levels of IL-4, IL-8, and TGF-β obtained in the T300 group were 31.1%, 20.9%, and 33.3%, respectively, which was lower than the cytokine increase in the ConA stimulated HFPGE-administered group (Table 4). These results indicate that stimulation with a mitogen enhances both cell-mediated immunity and humoral immunity after exposure to HFPGE, and was particularly effective for cell-mediated immunity.
Table 3
Effect of HFPGE on cytokine production in ConA-stimulated splenocytes from CPA-treated BALB/c mice
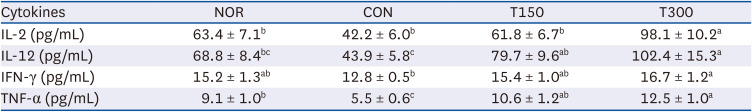
Values are expressed as the mean ± SEM (n = 10). Means without a common letter differ significantly at P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; ConA, concanavalin A; CPA, cyclophosphamide; NOR, normal control group (injected with vehicle + treated with vehicle); CON, control group (injected with CPA + treated with vehicle); T150, 150 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 150 mg of HFPGE/kg BW/day); T300, 300 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 300 mg of HFPGE/kg BW/day); IL, interleukin; IFN, interferon; TNF, tumor necrosis factor; BW, body weight.
Table 4
Effect of HFPGE on cytokine production in LPS-stimulated splenocytes from CPA-treated BALB/c mice

Values are expressed as the mean ± SEM (n = 10). Means without a common letter differ significantly at P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; LPS, lipopolysaccharide; CPA, cyclophosphamide; NOR, normal control group (injected with vehicle + treated with vehicle); CON, control group (injected with CPA + treated with vehicle); T150, 150 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 150 mg of HFPGE/kg BW/day); T300, 300 mg/kg BW/day HFPGE-treated group (injected with CPA + treated with 300 mg of HFPGE/kg BW/day); IL, interleukin; TGF, transforming growth factor; BW, body weight.
DISCUSSION
PG is a perennial vegetable belonging to the family Bellaceae. The roots are well known for their soothing, antipyretic, analgesic, and anti-inflammatory effects, and thus have traditionally been used for food or medicinal purposes [14]. It has been reported that an additional fermentation process during PG extraction enhances the pharmacological effects [616]. By adding a hydrolysis process, we obtained an extract with high total flavonoids, crude saponin, and platycodin contents as compared to fermented PGE. The HFPGE thus obtained showed excellent immunostimulatory effect in an in vitro study [7]. In the current study, we further confirmed the immune enhancing effect of HFPGE in BALB/c mice in an in vivo system.
In this study, the effect of HFPGE administration on the serum Igs levels of immunosuppressed mice was investigated. IgG is the most common xlink:type of Ig in blood and accounts for about 75% of the serum antibodies [17]. As a major component of human immunity, IgG binds to and neutralizes pathogens, activates the complement system, binds to the toxin and neutralizes it, and plays an important role in antibody-dependent cell-mediated cytotoxicity (ADCC) [18]. The IgA antibody plays an important role in the immune function of the mucous membrane. In blood, IgA interacts with Fc receptors to initiate inflammatory reactions, regulates ADCC, and plays a role in the degranulation of eosinophils, basophils, monocytes, and macrophages, phagocytosis by neutrophils, and respiratory burst activity [19]. IgM is the largest antibody, accounting for 5–10% of all antibodies. It is the first antibody secreted when exposed to an antigen [9]. Spleen is the major site of specific IgM production [20]. In our study, the concentrations of serum IgG, IgA, and IgM were decreased by 6.63%, 6.67%, and 50%, respectively, in the CON group as compared to the NOR group. This was increased by HFPGE administration by varying extents depending on the xlink:type of Ig; IgG was increased in both T150 and T300 groups as compared to the levels obtained in the CON group, but was significantly lower than the NOR group levels; IgA increased to the same level or higher in the HFPGE treatment groups than in the NOR group. However, there was no significant difference in the levels of IgM between T150 and T300; compared to the NOR group, maximum decrease of IgM was obtained in the CON group, which significantly increased and recovered to the NOR group level by both HFPGE administrations.
During the 4-week experimental period, the weight gain in the CON group was significantly reduced to 2.4 g compared to 3.9 g in the NOR group. This weight loss showed no recovery even in the HFPGE-administered groups: T150 (2.5 g) and T300 (2.3 g) (Fig. 2B). Other studies have reported similar observations that the treatment of CPA induces immunosuppression and causes weight loss as a side effect [2122]. However, unlike other papers [16] which reported that the administration of PG extract, especially an extract with a high platycodin D content, improved this weight loss problem, in our study, HFPGE administration could not recover the CPA-induced weight loss side effect. This may be due to differences in the age, experimental period, CPA dose and treatment time, and experimental diet between studies. The HFPGE doses used in this study (150 and 300 mg/kg BW) are within the 100–500 mg/kg BW levels used as extract concentrations in many other studies. Considering that even a lower dose of PGE (150 mg/kg BW) than in our study (300 mg/kg BW) had an effect on weight improvement, there is a possibility that weight loss may be a side effect caused by administering a high concentration of HFPGE. However, there was no difference in the dietary intake and behavior of the animals between all the experimental groups and the control group, thereby indicating that there is no problem in the stability of the concentration of HFPGE used in this study (data not shown). In the future, a follow-up study is needed to determine the cause.
When considering PGE extraction by the general method, an immunostimulatory effect was shown after administration of 500 mg/kg BW [23] However, even at a low dose of 75 mg/kg BW administration of fermented PGE, the Ig concentration was significantly higher than that of immunosuppressed animals. It was reported that fermented PGE had a greater immune-enhancing effect than the general PGE because of the high content of platycodin D present [16]. Comparing this with our study, although we administered a higher dose, the effect of increasing Ig levels in the blood was clear. In the previous study, fermented PGE had a greater immunostimulatory effect compared to normal PGE, with significant increase in the Ig concentration compared to immunosuppressed animals even with a low dose of 75 mg/kg BW. It was reported to be due to the high content of platycodin D [16]. In our HFPGE, not only fermentation but also a hydrolysis process was added, resulting in increased content of active ingredients such as platycodin D, thereby increasing the immune-enhancing effect.
CPA used as an immunosuppressant in this study is a drug used for the purpose of chemotherapy and suppression of the immune system. It is widely used for the treatment of various cancer patients, organ transplant patients, or for immunosuppression of autoimmune diseases [2425]. The CPA mechanism of action is mainly achieved by reducing the number of T cells, transmitting a signal to target cells, and suppressing the secretion of cytokines that affect the regulation of immune responses [26]. However, CPA treatment has many side effects such as nephrotoxicity, cardiotoxicity, and neurotoxicity [24]. Therefore, there is active research on traditional medicinal plants that can reduce these side effects. These plants usually have characteristics that are involved in T cell proliferation and activation [27]. CD4+ Th cells are roughly classified into functionally different subsets (Th1 and Th2) according to their secretory cytokines and immunological roles. In this study, we measured the serum concentrations of IL-12 and TNF-α (Th1 cytokines) and IL-8 and TGF-β (Th2 cytokines). All four cytokines were significantly decreased in the CON group compared to the NOR group, and increased in the HFPGE-treated groups. In the case of IL-8, only the NOR level was recovered by HFPGE administration. However, IL-12 was significantly increased in the HFPGE-treated group at a level more than twice the concentration of the NOR group. TNF-α and TGF-β were also significantly higher than the NOR group, by 1.3 and 1.2 times, respectively (Table 2). In other words, oral administration of HFPGE effectively improves both cell-mediated immunity and humoral immunity, and was more effective in improving cell-mediated immunity than humoral immunity.
The spleen is an important secondary immune organ. It is also involved in iron metabolism and erythrocyte homeostasis, and serves as a main filter for blood-borne pathogens and antigens. It has been shown that the spleen can also function as a secondary lymphoid organ. The spleen contains immune cells such as T cells, B cells, dendritic cells, and macrophages [28]. Hematopoietic stem and progenitor cells (HSPCs) exist in the bone marrow, but some of them enter the circulation under normal conditions [29]. When conditions are right, circulating HSPCs generate lineage descendant cells outside the bone marrow [30]. This process, called extramedullary hematopoiesis, occurs mainly in the developing embryo’s liver, but can also occur in adult tissues, including the spleen. In certain disease conditions, splenic HSPCs expand greatly and generate the progeny locally [28]. As such, in the spleen, various immune stimuli occur between antigen presenting cells (macrophage, dendritic cells) and lymphocytes (T cells, B cells) that detect the antigen, leading to the production of various antibodies and activation of T cells. In this study, splenocytes were isolated from the spleen of BALB/c mice raised on an HFPGE treatment, and the effect of HFPGE on proliferation was investigated. In splenocytes not stimulated with mitogen, the proliferation index was decreased in CPA-treated animals, but HFPGE administration significantly increased the index, more than the NOR group. Similar results were obtained in splenocytes stimulated with ConA or LPS (Fig. 3). Therefore, we determined that exposure to HFPGE significantly increases the proliferation of both T cells and B cells.
Cytokines are produced by various immune cells and play a major role in immune regulation by influencing the activation, growth, and differentiation of immune cells. Activation of splenocytes results in the secretion of various cytokines to regulate and induce immune responses. Therefore, measurement of the induced secretion of these cytokines is a very important indicator for determining the immune activity of splenocytes [28]. Cytokines are classified according to several criteria. Representative cytokines secreted by Th1 cells include IL-1β, IL-2, IL-12, TNF-α, and IFN-γ, which enhance the cell-mediated immune response. Representative cytokines secreted by Th2 cells include IL-4, IL-5, IL-8, IL-10, and TGF-β, which enhance the humoral immune response [31]. In this study, to determine the effect of HFPGE on the concentration of cytokines secreted by splenocytes, the splenocytes were stimulated with mitogen and cultured, following which the concentrations of cytokines were measured in the conditioned media. We determined that cytokines secreted by ConA-stimulated splenocytes (IL-2, IL-12, IFN-γ, and TNF-α) and cytokines secreted by LPS-stimulated splenocytes (IL-4, IL-8, and TGF-β), both were significantly reduced in the CON group compared to NOR group. However, in the T150 and T300 groups, the cytokine concentrations were increased to a level that was statistically equal to or higher than that of the NOR group after stimulation with the two mitogens. In particular, there was a dramatic increase in cytokines secreted by ConA-stimulated splenocytes following HFPGE administration (Tables 3 and 4).
According to a meta-analysis conducted by Jung et al. [32], PGE is administered at 6 mg/day to 8 g/day in clinical trials, and the dose varies greatly depending on the extraction method of PGE and the target disease. In this study, 150 and 300 mg/kg BW/day administered to mice correspond to 0.75 and 1.5 g/day, respectively, when applied to the human. This is considered a moderate dose compared to other studies. Investigating the bioavailability and the physiological concentration of HFPGE used in this study is very important for future application to humans. There has been no report on the bioavailability and physiological concentration of PGE so far; hence, there is a need for future studies on the bioavailability and physiological concentration of HFPGE.
In conclusion, our results confirm the immune-enhancing effect of HFPGE in a CPA-induced immunosuppressed animal model. HFPGE administration resulted in increased levels of serum Igs (IgG, IgA, and IgM), serum cytokines (IL-12, TNF-α, IL-8, and TGF-β), proliferation index of splenocytes, and cytokine secretion in mitogen-stimulated splenocytes. These results indicate that HFPGE stimulates the innate and adaptive immunity in immunosuppressed conditions, and increases cytokines, Igs, and immune cell proliferation, thereby enhancing the immune response. Therefore, we propose that HFPGE has the potential to be used as functional foods and medicine for immune recovery in various situations with reduced immunity. However, there was little dose-dependent effect between 150 and 300 mg/kg BW HFPGE. Moreover, decreases in the BWs due to CPA toxicity were not completely recovered by HFPGE treatment. In the future, more studies are needed to find the appropriate treatment concentration of HFPGE and to determine the CPA treatment required to optimize the experimental animal model to measure immune-enhancing effects.
 XML Download
XML Download