

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
There is the view that hypertension is associated with cognitive impairment and the view that no association is observed. Atrophy and loss of nerve cells and change in the cholinergic system were reported in both human [1234] and animal studies [567] of congenital hypertension, Recently, angiotensin (Ang)-(1-12) was discovered in the brain, and Ang-(1-12) was reported to have a blood pressure control function [8]. A decrease in blood pressure leads to the conversion of angiotensinogen in the blood to Ang I by renin, secreted by the kidneys. Ang I is further converted to Ang II by angiotensin-converting enzyme (ACE). Ang II is the key component in increasing blood pressure levels [9].
Most of the renin-angiotensin system (RAS) components are also found in the brain, and brain RAS is involved in cognitive function and behavioral regulation [10]. Changes in RAS components in the brain, especially Ang II, ACE activity, and Ang II receptors, were reported in Alzheimer’s disease (AD) in animal models [101112131415]. Ang II in the brain inhibited the secretion of acetylcholine from the amygdala and cortex and regulated neurotransmission via Ang II receptors [1116]. In the scopolamine (SCO)-treated rat model, a significant decrease in acetylcholine (ACh) levels and an increase in acetylcholinesterase (AChE) activity occurred, which was treated with angiotensin II receptor type 1 (AT1) blockers [16]. Clinical studies have also shown that blockade of chronic Ang II receptors improves memory in the elderly [17181920].
Thus, the evidence suggests that inhibition of RAS to improve cognitive function may be a novel strategy. However, most of the studies to date have reported on the effect of suppressing RAS by drugs and there is no research on cognitive improvement through RAS inhibition of food ingredients so far. Therefore, if we select ingredients with excellent antioxidant and anti-inflammatory properties from frequently consumed foods and reveal a new mechanism of cognitive improvement by inhibition of RAS, it will be possible to suggest a way to improve the quality of life of the elderly as well as academic development.
Curcumin is a spice derived from turmeric and is known to exhibit anti-inflammatory and antioxidant effects [21]. Curcumin has been shown to have a protective effect against hypertension and inflammatory diseases via the regulation of RAS components, like Ang II, ACE1, and AT1 protein levels [22232425]. Curcumin not only prevented oxidative stress in the brain by reducing the accumulation of beta-amyloid but also improved cognitive function [26272829]. Administration of curcumin to ICR mice reduced blood pressure by blocking RAS components in the brain and protecting the cholinergic system [30]. However, the relationship between curcumin and cognitive function mediated via brain RAS in the hypertensive rat has yet to be reported. Therefore, this study investigated whether the inhibition of RAS by curcumin had a positive effect on brain cognitive function in spontaneously hypertensive rat/Izm (SHR/Izm).
Go to : 
MATERIALS AND METHODS
Materials
Curcumin (Curcuma longa L., 65%) and scopolamine ((−)-scopolamine hydrobromide trihydrate), and tacrine (9-amino-1,2,3,4-tetrahydroacridine hydrochloride hydrate) were purchased from Sigma (Sigma Aldrich, St. Louise, MO, USA). Tween 80 (polyoxyethylene sorbitan monooleate) and 20X phosphate-buffered saline, which are solvents for curcumin and tacrine, were purchased from Daejung (Daejung Chemicals & Metals, Siheung, Korea) and Dongin (Dongin Biotech, Seongnam, Korea), respectively. Scopolamine is an antagonist of muscarinic choline receptors and was used to impair cognitive function in experimental animals. Tacrine is an acetylcholine hydrolase inhibitor that treats cognitive function. Curcumin and tacrine were dissolved in 10% Tween 80, and scopolamine was dissolved in sterile distilled water before use.
Animals and study design
The experimental animals were a 6-week-old male SHR/Izm with a history of essential hypertension, which was purchased from Daehan Biolink (Chungbuk, Korea) along with AIN-93G solid feed (Table 1). SHR/Izm is an experimental animal produced to have a hypertension gene by selectively crossing mice with hypertension among Wistar-Kyoto (WKY) rats, and it is known that hypertension usually appears from 5 to 6 weeks of age [31]. The laboratory environment was maintained at a constant temperature of 22–24°C and humidity of 55–65%. This animal study was approved by Dankook University Animal Ethics Committee (DKU-19-015) and conducted by institutional and national guides for the care and use of laboratory animals.
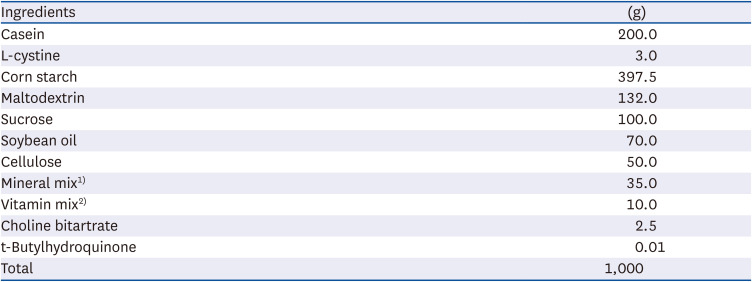
Table 1
Composition of AIN 93G diet
1)Mineral mix (g/kg): potassium phosphate 196 g, sodium chloride 74 g, potassium citrate 70.78 g, calcium carbonate 357 g, magnesium oxide 24.3 g, zinc carbonate 1.65 g, ferric citrate 6.06 g, cupric carbonate 0.31 g, manganous carbonate 0.63 g, potassium iodate 0.01 g, sodium selenate 0.0103 g, ammonium paramolybdate 0.008 g, sodium meta-silicate 1.54 g, lithium chloride 0.0174 g, chromium potassium sulfate 0.275 g, sodium fluoride 0.0635 g, boric acid 0.0815 g, nickel carbonate hydroxide 0.0318 g, ammonium meta-vanadate 0.0066 g, sucrose 220.7159 g, sodium selenate anhydrous 10.25 mg.
2)Vitamin Mix (g/kg): niacin 3.0 g, pyridoxine HCl 0.7 g, thiamine 0.6 g, folic acid 0.2 g, calcium pantothenate 1.6 g, biotin 0.02 g, vitamin E 15.0 g, vitamin B12 2.5 g, riboflavin 0.6 g, vitamin D3 0.2 g, vitamin A 0.8 g, vitamin K1 0.075 g, sucrose 974.705 g.
![]()
The SHR were freely fed with an AIN-93G diet and water during the experimental period (5 weeks). Following acclimation for one week, the rats were divided into 5 groups using a randomized block design: CON group (saline-treated, n = 8), SCO group (treated with scopolamine [1 mg/kg BW], n = 8), TAC group (positive control, treated with scopolamine [1 mg/kg BW] + tacrine [10 mg/kg BW], n = 8), CUR 100 group (treated with scopolamine [1 mg/kg BW] + curcumin [100 mg/kg], n = 8), and CUR 200 group (treated with scopolamine [1 mg/kg BW] + curcumin [200 mg/kg BW], n = 8). The detailed experimental treatment protocol for 4 weeks is as follows.
The treatments were administered orally (10% Tween 80, curcumin or tacrine) and intraperitoneally (in the case of scopolamine) for 4 weeks at a fixed time every day. During the first 2 weeks, curcumin was orally administered to the CUR100 and CUR200 groups (once a day). Tween 80 (vehicle) instead of curcumin was orally administered to the remaining 3 groups (CON, TAC, and SCO group). For the next 2 weeks, 30 minutes before the behavioral experiment, scopolamine was administered intraperitoneally to 4 groups (except the CON group) daily to reduce cognitive function. In addition, 10% Tween 80 was orally delivered to the CON and the SCO groups, tacrine to the TAC group, and curcumin (100 or 200 mg) orally administered to the CUR100 or CUR 200 groups every day 1 hour before the behavioral experiment, for the remaining 2 weeks. The concentrations of the curcumin administered to the SHR were based on a previous study. The concentration of the curcumin administered to the SHR was set in consideration of toxicity based on previous studies [252627282930]. The amount of curcumin (100 or 200 mg/kg BW) consumed by SHR in this study can be estimated to be about 15 mg and 30 mg according to the human equivalent dose calculation method based on an adult weighing 70 kg [26].
Weight change, dietary intake, and dietary efficiency
The body weight of rats was measured every 2 days, and dietary intake was measured every 3 days at a specific time until the end of the study. The weight gain was calculated by dividing the difference between the weight before sacrifice and the initial weight by the breeding period. Dietary intake was calculated based on the difference between the supplied diet and the remaining diet, and the food efficiency ratio (FER) was calculated as follows: FER = Body weight gain during the experimental period/Food intake during the experimental period.
Blood pressure measurement
Blood pressure was measured three times with a blood pressure monitor (BP-2000 blood pressure analysis system, rat platform 4-channel, Visitech Systems, Apex, NC, USA) using the tail-cuff method. The experimental animal was placed in the holder, followed by insertion of the tail into the cuff. To ensure stability, it was acclimated at 37°C for 15 min and then measured 20 times for 30 min.
Sample preparation
After the final blood pressure measurement, the rats fasted for more than 12 h, and anesthesia was performed with avertin (2, 2, 2-tribromoethanol; Sigma Aldrich). Blood was drawn, and the heart, liver, kidney, epididymal fat, and brain tissue of the experimental animals were removed and weighed. The collected tissues and serum were stored at −70°C until the experiment.
Behavioral experiment
Y-maze test
Y-maze test was conducted to evaluate the effects of curcumin on spatial cognition and short-term memory [32]. The Y-maze is a black acrylic plate composed of three branches, each measuring 45 cm in length, 10 cm in width, and 35 cm in height. The angle between the three branches was set at 120°. The three different areas of the branches were designated as A, B, and C, respectively. The experimental animals were placed in one area and allowed to move freely for 8 minutes to observe and record their movement using Smart 3.0 (Harvard Apparatus, Holliston, MA, USA). The number of entries was recorded as the number of times the tail entered one of the branches completely, and actual alternation was awarded for each entry into the three different areas. The alternation behavior (%) represented by movement into a new branch rather than the previous branch was calculated using the following formula.
Passive avoidance test
A passive avoidance test was conducted to measure long-term memory [33], using the preference of the experimental animals for darker places. The instrument used for the passive avoidance test (Harvard Apparatus) consisted of a bright area with illumination (32 × 32 × 27 cm) and a dark area (18 × 12 × 12 cm), with the two areas separated by a middle door (8 × 8 cm); each area consisted of a wire mesh on the floor. After placing the rats in a bright area and letting them search freely for 10 s, the middle door was opened to allow entry into the dark area. Upon entry into the dark area, the middle door was closed, and an electric shock of 0.3 mA was generated through the wire mesh for 3 s. The time taken by the rat to enter the dark area from the initial location in the bright area was defined as a trial. The experiment was repeated 3 times. A retention test was performed 24 h after the end of the trial, and the time taken by the rat to enter the dark area after opening the middle door was defined as the latency time, limited to a maximum of 300 s.
RAS and cholinergic system
The concentration of Ang II in brain tissue was assayed using the Rat Ang II ELISA Kit from Cusabio (Cusabio, MD, USA). The concentration of acetylcholine in brain tissue was measured with an Acetylcholine Elisa kit from MyBioSource (MyBioSource, San Diego, CA, USA). For the study of RAS, mRNA expressions of ACE and AT1 were determined. For the study of the cholinergic system, the mRNA expression of muscarinic m1 acetylcholine receptor (m1AChR), and muscarinic m2 acetylcholine receptor (m2AChR) was determined. The experimental method related to mRNA expression was described in detail in a previous study [30]. In brief, following the addition of Tris reagent to the homogenized brain tissue, the sample mRNA was extracted using chloroform, isopropanol, and 75% ethanol. A SuperScript kit (Invitrogen, CA, USA) was used to reverse-transcribe the extracted mRNA into cDNA. Oligo DT, 2X reaction mixture (10X RT buffer, 0.1M DTT, 10 mM NTP), superscript II reverse transcriptase, and RNase H was sequentially all added and incubated to synthesize cDNA. The following materials were added: prepared cDNA sample, SYBP Green Master Mix (Applied Biosystems, Foster City, CA, USA), nuclease-free water, and forward/reverse primers consisting of ACE (5′-GCGCTTGACCCTGGATTGC-3′ and 5′-GGCTCGACTTGGTGGTAGAG-3′), AT1 (5′-CTCTGCCACATTCCCTGAGTTA-3′ and 5′-TGGGGCAGTCATCTTGGATTC-3′), m1AChR (5′-CATTGGCTGCTACAAACACCC-3′ and 5′-GCTAACTGCACGGCCATCTA-3′), and m2AChR (5′-CTGGCCTTTGGGACCTGTAG-3′and 5′-GCCATTTTTGTGGTCCGCTT-3′). All assays were independently repeated three times.
Statistical analysis
All data were expressed as the mean and standard error using the SPSS program (IBM SPSS Statistics 23.0; IBM Corp., Armonk, NY, USA), and the significance among groups was determined at the P < 0.05 level using Duncan’s multiple range test after ANOVA.
Go to :
RESULTS
Body and tissue weights and food intake
The initial and final body weights of rats did not vary significantly between different groups (Table 2). No significant differences were found in dietary intake or FER between the experimental groups. Tacrine and curcumin intake did not affect the body weight or dietary intake of rats. The weights of liver, kidney, epididymal fat, and brain did not differ significantly among the experimental groups, suggesting that tacrine and curcumin intake did not affect the tissue weight (Table 3).
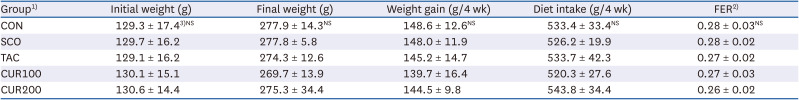
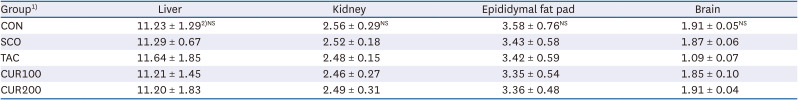
Table 2
Body weight, weight gain, food intake and FER of experimental rats
FER, food efficiency ratio.
1)CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg).
2)FER = Body weight gain for 4 weeks/Food intake for 4 weeks.
3)Mean ± SE.
![]()
Table 3
Organ weight of experimental rats (g)
1)CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg).
2)Mean ± SE.
![]()
Blood pressure
There was no significant difference in systolic and diastolic blood pressure of SHR among the groups during days 0 and 15 of oral curcumin administration (Fig. 1). However, on day 29, both the systolic and diastolic blood pressure were significantly lower in the curcumin intake group (CUR 200) than in the other groups (P < 0.05).
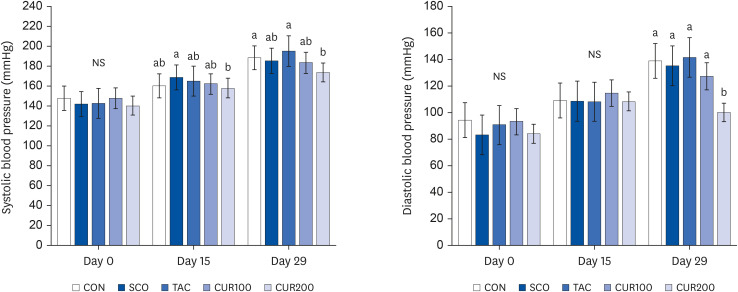 | Fig. 1Effect of curcumin on systolic (left) and diastolic blood pressure (right) in spontaneously hypertensive rat. The data showed mean systolic and diastolic blood pressure measured on days 0, 15, and 29 of the experiment. Different letters above each bar indicate significant differences among groups as determined by Duncan’s multiple range test (P < 0.05).CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg); NS, not significant.
|
Cognitive function
The results of the Y-maze test are shown in Fig. 2. The SCO group exhibited the lowest alteration behavior due to cognitive impairment caused by scopolamine (1 mg/kg, P < 0.05, Fig. 2 [left]). The alteration behavior of the TAC group treated with tacrine, which is cognitive therapy, was significantly higher than in the SCO group at 15.84 ± 3.86% (P < 0.05), The change in the alteration behavior of the curcumin intake group was significantly higher than in the SCO group (P < 0.05), but there was no significant difference based on curcumin intake. The number of visits to the three branches, defined as a total number of arm changes, was measured to compare the behavioral patterns and movement states of the experimental animals. There was no significant difference in the total number of arm changes among groups (Fig. 2 [right]).
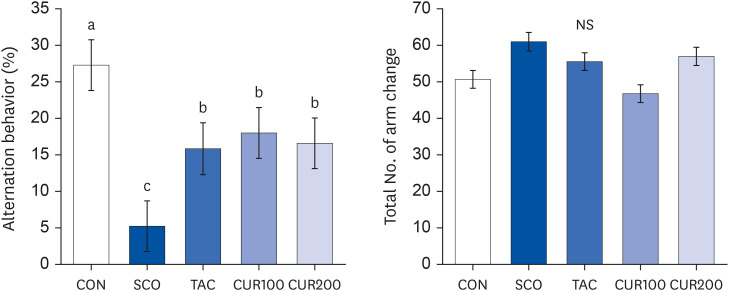 | Fig. 2Effect of curcumin on alternation behavior (left) and the total number of arm entries (right) of Y-maze test in spontaneously hypertensive rat. Alternation behavior (%) (right) was calculated as follows: Alternation behavior (%) = total number of alternations/(total number of arm entries – 2) × 100. Total number of arm entries (left) under the same experimental condition. Different letters above each bar indicate significant differences among groups as determined by Duncan’s multiple range test (P < 0.05).CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg); NS, not significant.
|
The passive avoidance test determines the habit of an experimental animal seeking a dark place. The results of the passive avoidance test are shown in Fig. 3. Compared with the CON group, the latency time, which is the time taken to enter the dark space following electric shock, was significantly lower in the SCO group (P < 0.05). The CUR100 and CUR200 groups showed significantly higher latency times than the SCO group (P < 0.05), but no significant difference was detected according to the dose of intake.
 | Fig. 3Effect of curcumin on latency time of passive avoidance test in spontaneously hypertensive rat. The data represent mean latency time(s). Total number of arm entries under the same experimental condition. Different letters above each bar indicate significant differences among groups as determined by Duncan’s multiple range test (P < 0.05).CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg).
|
RAS
The Ang II concentration in the brain of the SCO group was significantly higher than in the CON group, whereas it was significantly lower than in TAC and CUR200 groups, compared to the SCO group (Fig. 4 [left], P < 0.05). The mRNA expression of AT1 in brain tissue was significantly higher in the SCO group and significantly lower in the CUR200 group compared with the CON group (P < 0.05) (Fig. 4 [right]). However, there was no significant difference in mRNA expression of AT1 according to the curcumin concentration. The mRNA expression of ACE was significantly higher in the TAC and CUR200 group than in the CON group by approximately 52% and 65%, respectively (P < 0.05) (Fig. 5). The mRNA expression of ACE in the CUR200 group was significantly lower than that in the SCO group (P < 0.05).
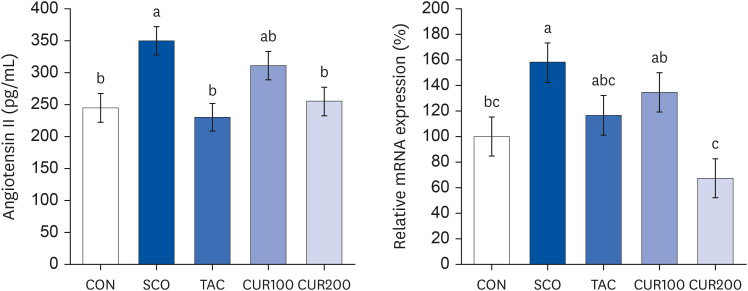 | Fig. 4Effect of curcumin on Angiotensin II content (left) and on the mRNA expression of angiotensin II receptor (right) in brain. Angiotensin II content was determined by using an assay kit (left). The total RNA was isolated and real-time polymerase chain reaction was performed (right) and GADPH level was used as the loading control Different letters above each bar indicate significant differences among groups as determined by Duncan’s multiple range test (P < 0.05).CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg).
|
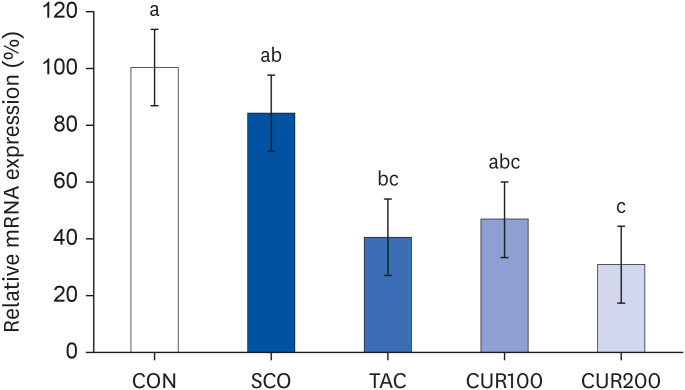 | Fig. 5Effect of curcumin on the mRNA expression of angiotensin-converting enzyme in the brain. Total RNA was isolated and real-time polymerase chain reaction was performed. GADPH levels were used as the loading control. Different letters above each bar indicate significant differences among groups as determined by Duncan’s multiple range test (P < 0.05).CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg).
|
Cholinergic system
The content of ACh was significantly lower in the SCO group by about 56% compared with the CON group (P < 0.05) (Fig. 6). The TAC group showed significantly higher levels of ACh than the SCO group by about 80% (P < 0.05). The CUR200 group showed a significant increase in the content of ACh compared with the SCO group, without curcumin dose-dependently (P < 0.05). The expression of m1AChR and m2AChR in brain tissue is shown in Fig. 5. The expression of m1AChR mRNA was significantly lower in the SCO group than in the control group, and the mRNA expression of m1AChR was significantly increased by treatment of both tacrine and curcumin (Fig. 7 [left]). In the case of curcumin treatments, the level of m1AChR expression increased in a concentration-dependent manner. The results of mRNA expression of m2AChR also appeared like the result of m1AChR expression (Fig. 7 [right]). The level of m2AChR mRNA in CUR100 or CUR200 was significantly increased compared with the SOC group. In the CUR 200 group, the mRNA expression of m2AChR was increased similarly to the positive control group, the TAC group.
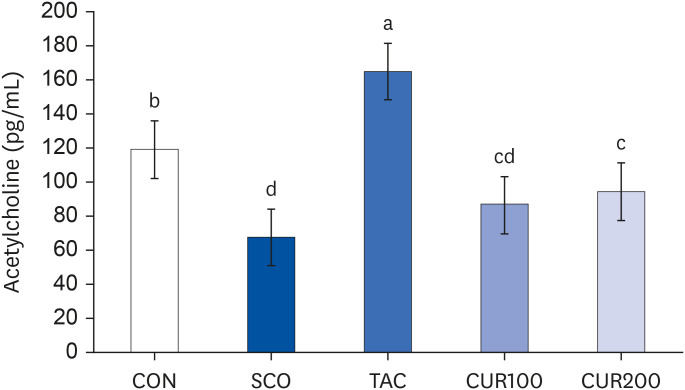 | Fig. 6Effect of curcumin on acetylcholine content in brain.Different letters above each bar indicate significant differences among groups as determined by Duncan’s multiple range test (P < 0.05).
CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg).
|
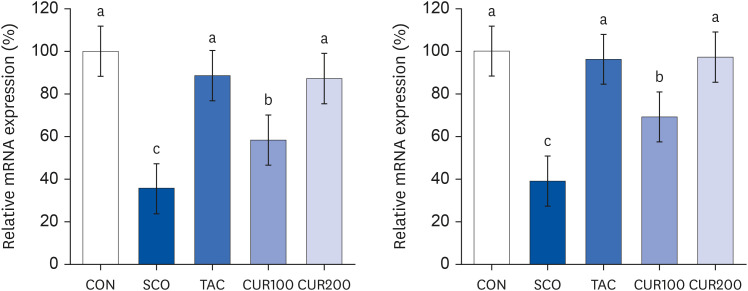 | Fig. 7Effect of curcumin on the mRNA expression of muscarinic acetylcholine receptor type I (left) and muscarinic acetylcholine receptor type II (right) in the brain. Total RNA was isolated and real-time polymerase chain reaction was performed. GADPH levels served as the loading control. Different letters above each bar indicate significant differences among groups as determined by Duncan’s multiple range test (P < 0.05).CON, control; SCO, scopolamine (1 mg/kg); TAC, scopolamine (1 mg/kg) + tacrine (10 mg/kg); CUR100, scopolamine (1 mg/kg) + curcumin (100 mg/kg); CUR200, scopolamine (1 mg/kg) + curcumin (200 mg/kg).
|
Go to :
DISCUSSION
The purpose of this study was to investigate whether curcumin intake improves cognitive function by regulating brain RAS and cholinergic system in the SHR/Izm. The SHR/Izm is characterized by hypertension onset usually from 5–6 weeks of age [31]. SHR is also characterized by loss of nerve cells and impaired cholinergic neurotransmission and is therefore used to assess hypertensive brain damage and treatment [3435].
In this study, cognitive decline was induced by injecting scopolamine (SCO) into a 6-week-old SHR. SCO is a cholinergic muscarinic receptor antagonist. Several studies demonstrated that AD in animals induced by SCO facilitated the study of the mechanism of the cholinergic nervous system [171819]. SCO-induced brain RAS alteration was reversed by cholinesterase inhibitors physostigmine and tacrine [1127]. Thus, in this study, SHR were treated with scopolamine to induce cognitive impairment and then treated with either tacrine, as a positive control, or curcumin.
After 4 weeks of curcumin intake, especially CUR200, the blood pressure was significantly decreased in SHR. In this study, the weight change or dietary intake between the control and the curcumin groups was not significantly different, suggesting that the reduction of blood pressure in the curcumin-treated group is thought to be the curcumin intake.
The Y-maze test was conducted to determine the short-term memory of SHR and evaluated the transient movement pattern of the experimental animals in the three branches [32]. The Y-maze test showed no significant difference in the total number of arms changes in all groups, suggesting that the changes in spontaneous alternation behavior were not due to locomotor deficits, but due to cognitive dysfunction induced by scopolamine. Previous studies also reported that intraperitoneal treatment with scopolamine resulted in a decrease in cognitive function and thus altered behavior [3637]. The alteration behavior (%) in the TAC group or the curcumin intake groups (CUR100 or CUR200) was significantly higher than in the SCO group (P < 0.05), which suggested that scopolamine-induced cognitive impairment was improved by curcumin treatment.
Latency time (s), the time to enter the dark zone after receiving an electric shock, 24 h after the end of learning through electrical stimulation was measured in the passive avoidance test [33]. In this study, administration of curcumin significantly improved the latency time in SCO-induced hypertensive rats, suggesting that the decreased latency time, due to intraperitoneal administration of SCO, was improved by curcumin intake, which was effective in improving long-term memory. Previous studies have reported similar results, despite differences in the behavioral test method or the amount of curcumin administered to animals, ranging from 20 to 300 mg/kg [262728293038394041].
In this study, we investigated whether the effect of curcumin on cognitive function in SHR was related to brain RAS and the cholinergic system. Curcumin intake in SHR decreased Ang II content as well as the mRNA expression of ACE and AT1 in the brain tissue. In addition, compared with the SCO group, the administration of curcumin increased ACh concentration as well as the mRNA levels of m1AChR and m2AChR.
Increased Ang II, a major component of RAS, in the brain has been found to decrease the release of ACh [14]. ACh deficiency has been reported in patients with Alzheimer’s disease manifesting impaired learning and memory [15]. Ang II is activated when Ang II binds with its receptor: AT1 or AT2. Ang II exerted behavioral effects mainly via AT1 in memory-impaired mice [2223], scopolamine-induced memory impairment in rodents [1117], or renin/angiotensinogen transgenic mice [131628], Thus blockers of AT1 have been used as antihypertensive and anti-AD drugs in humans with hypertension [1819204243]. Our previous study also reported that among phytochemicals, curcumin significantly inhibited ACE activity, and the mRNA expressions of ACE and AChE in ICR rats induced hypertension [30].
A limitation of this study was that we did not measure plasma or brain curcumin concentrations after oral administration of curcumin to rats. Thus, we do not know how much curcumin administered was absorbed into the body of the mice. Although the bioavailability of curcumin is low, it has been reported that it crosses the blood-brain barrier and reaches the brain due to its lipophilicity [44].
In this study, the administration of curcumin (100 and 200 mg/kg BW) was effective in lowering blood pressure and improving cognitive function, and in particular, Cur 200 was more effective, which suggests that curcumin was administered to mice was delivered to the brain. Concentrations of curcumin used in this study (100–200 mg/kg BW) correspond to the daily intake of 15 mg to 30 mg of curcumin in humans, depending on the human equivalent dose calculated based on an adult body weight of 70 kg [26]. The amount of curcumin investigated in clinical trials varies from 10–1,000 mg/day, and it is known that up to 700 mg/day of curcumin intake is not harmful to the human body [44]. Therefore, additional studies on various curcumin concentrations that can have the maximum effect on hypertension and cognitive function are needed.
To the best of our knowledge, the mechanism of how curcumin simultaneously improves blood pressure and cognitive function in hypertensive rats has not yet been reported. This study showed that curcumin improved cognitive function through the Y-maze test and passive avoidance test, and this improvement was mediated through brain RAS inhibition and brain cholinergic system enhancement. Therefore, the results of this study suggest the possibility that the intake of phytochemical foods such as curcumin, rather than medicinal products, can lower blood pressure and improve cognitive function in hypertensive animals. Furthermore, this study might be provided basic data for future research that is helpful for the quality of life and health of the elderly in a situation where the number of dementia patients is increasing. In conclusion, curcumin treatment of rats with congenital hypertension improved cognitive function via inhibition of RAS (by decreasing Ang II and AT1 receptor expression) and via protection of the cholinergic system (by increasing acetylcholine and acetylcholine receptors expression).
Go to :
 XML Download
XML Download