

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
The aging process is characterized by a cascade of cellular alterations and gradual deterioration in an organism’s metabolism. An increasing number of studies have highlighted the importance of exploring age-related diseases, including neurodegenerative and cardiovascular diseases [1,2]. Genome-wide gene expression analysis can provide insights into the molecular changes that underlie diseases, leading to a greater understanding of human biology [3]. Age-associated transcriptional alterations in cardiomyocytes isolated from 4- and 2-month old mice identified reduced activity of the mitochondrial electron transport system and cardiac-specific transcription regulators in older mice [4]. In addition, a recent study showed that transcriptional changes associated with aging were prominent in endothelial cells, and were directly related to functional changes in the aged brain [5].
Rats are larger in size, which exhibits advantages in the regard of handling and sampling, particularly in various surgical challenges. Indeed, genetic studies in rats have enabled genome-wide comparisons across species, and this could ultimately improve the transition of research to advance therapeutics for human diseases [6-8]. A RNA-seq BodyMap database showing organ-, age-, and sex-specific differential expression in 11 organs of rats ranging from juvenile to old age has been established [7,9]. Here, we use this valuable transcriptomic dataset [7,9] and our published computational pipeline, Sailfish-cir [10] to investigate the expression profile of ion channel genes across different organs/tissues including adrenal gland, brain, heart, kidney, liver, lung, muscle, spleen, thymus, testes, and uterus, and developmental stages (2-/6-/21-/104-week) in both sexes of Fischer 344 rats. Based on the knowledge of the diversity of ion channel subtypes and functions in entire tissues, we determined the extent to which ion channel genes showed a tissue-dependent pattern in the 11 organ/tissue types. The expression of several ion channel genes occurred in a tissue-specific manner; notably, the brain showed higher tissue-specific expression of ion channel genes than the other organs. We also found that the expression of ion channel genes in the thymus, including Aqp1, Clcn4, Hvcn1, Itpr1, Kcng2, Kcnj11, Kcnn3, and Trpm2, was positively correlated with age. We provide a comprehensive ion channel gene signature that occurs in tissue-specific and age-dependent manner.
METHODS
Processing of the rat BodyMap transcriptomic data
The rat BodyMap RNA-seq data were downloaded from the NCBI Gene Expression Omnibus (GEO) database [11] with the accession number of GSE53960 [7,9]. The quality of the sequencing data was assessed by RNA-SeQC [12] and the raw sequencing reads were filtered by removing adaptor sequences, contamination, and low-quality reads. We quantified the expression levels for all the protein-coding genes according to the Ensembl rat gene annotation [13] (release 91) using the Sailfish-cir framework [14] with default settings.
Tissue specificity
Based on the rat BodyMap transcriptomic data, we computed the tissue specificity of each rat protein-coding gene. We applied the “tau” method developed by Yanai et al. [15] to calculate the tissue specificity index (TSI): , where TSI stands for the tissue specificity index; N represents the number of tissue types; and ei denotes the mean expression of a given gene in tissue i normalized to its maximum expression across all the tissues. A gene with high TSI implies this gene shows a strong tissue-specific expression.
Statistical analyses
All the statistical analyses were performed using the R statistical platform. Principal component analysis (PCA) was applied to visualize the tissue specificity of ion channel gene expression using the function “dudi.pca” in the R package “ade4”. The function “heatmap.2” in the R package “gplots” was employed to generate hierarchical clustering for the tissue-specific ion channel genes. The R generic functions, e.g., “t.test”, “ks.test”, “wilcox.test”, and “cor.test”, were applied to conduct t-test, Kolmogorov–Smirnov test, Wilcoxon test, and Spearman’s rank correlation test, respectively.
RESULTS
A comprehensive repertoire of ion channel gene expression
The definition of all the rat ion channel genes was obtained from the International Union of Basic and Clinical Pharmacology/British Pharmacological Society Guide to PHARMACOLOGY [16]. In total, 141 voltage-gated ion channel genes, 75 Ligand-gated ion channel genes, and 50 other ion channel genes were collected. To explore the dynamic expression pattern of ion channel genes, we obtained the rat RNA-seq transcriptomic BodyMap [7,9] from the GEO database [11] (GEO accession: GSE53960). Using our published RNA-seq quantification tool, Sailfish-cir [14], we summarized the ion channel gene expression profiles from the rat BodyMap dataset, which consists of 320 rat samples from 11 organ/tissue types, including adrenal gland, brain, heart, kidney, liver, lung, muscle, spleen, thymus, testes, and uterus across four developmental stages, i.e., 2-, 6-, 21- and 104-week. For each ion channel gene, the transcripts per million (TPM) was calculated to measure the expression level. We next investigated the top 30 highly expressed ion channel genes among the different tissue types. The ion channel genes that are among the top lists in at least eight tissue types were deemed “housekeeping”. In total, we identified 12 housekeeping ion channel genes, including Aqp1, Chrna7, Clcn3, Gja1, Itpr1, Itpr2, Mip, Orai1, P2rx4, Pkd2, Tpcn1, and Trpm7 (Fig. 1). The “housekeeping” ion channel genes defined by more/less stringent criteria were demonstrated in Supplementary Fig. 1 and 2, respectively.
Ion channel genes show higher tissue specificity than the other genes
We systematically investigated the tissue-specific expression for the ion channel genes. To understand the extent to which the expression of ion channel genes shows a tissue-dependent pattern, PCA upon ion channel gene expression was applied for all the samples. We found that the samples of the same organ/tissue type tended to aggregate together according to the first and second principal components (Fig. 2A), suggesting that ion channel genes are expressed in a strong tissue-specific manner. Similar to the previous observation [8], the brain samples showed an extremely unique PCA pattern compared with the other organ/tissue types (Fig. 2A), which was further confirmed by the fact that the mean TPM values of ion channel genes were significantly higher in the brain compared with the other tissue types (Kolmogorov–Smirnov test: p < 10–9; Fig. 2B). We further investigated the difference in tissue specificity between the ion channel genes and the other genes in rat transcriptome. We calculated TSI (see Methods for details) for both the ion channel genes and the other genes. We found that the TSI of ion channel genes was significantly higher than that of the other rat genes (Wilcoxon test: p < 10–10; Fig. 2C), which suggests that, compared with the other genes, ion channel genes are expressed in a more tissue-specific manner.
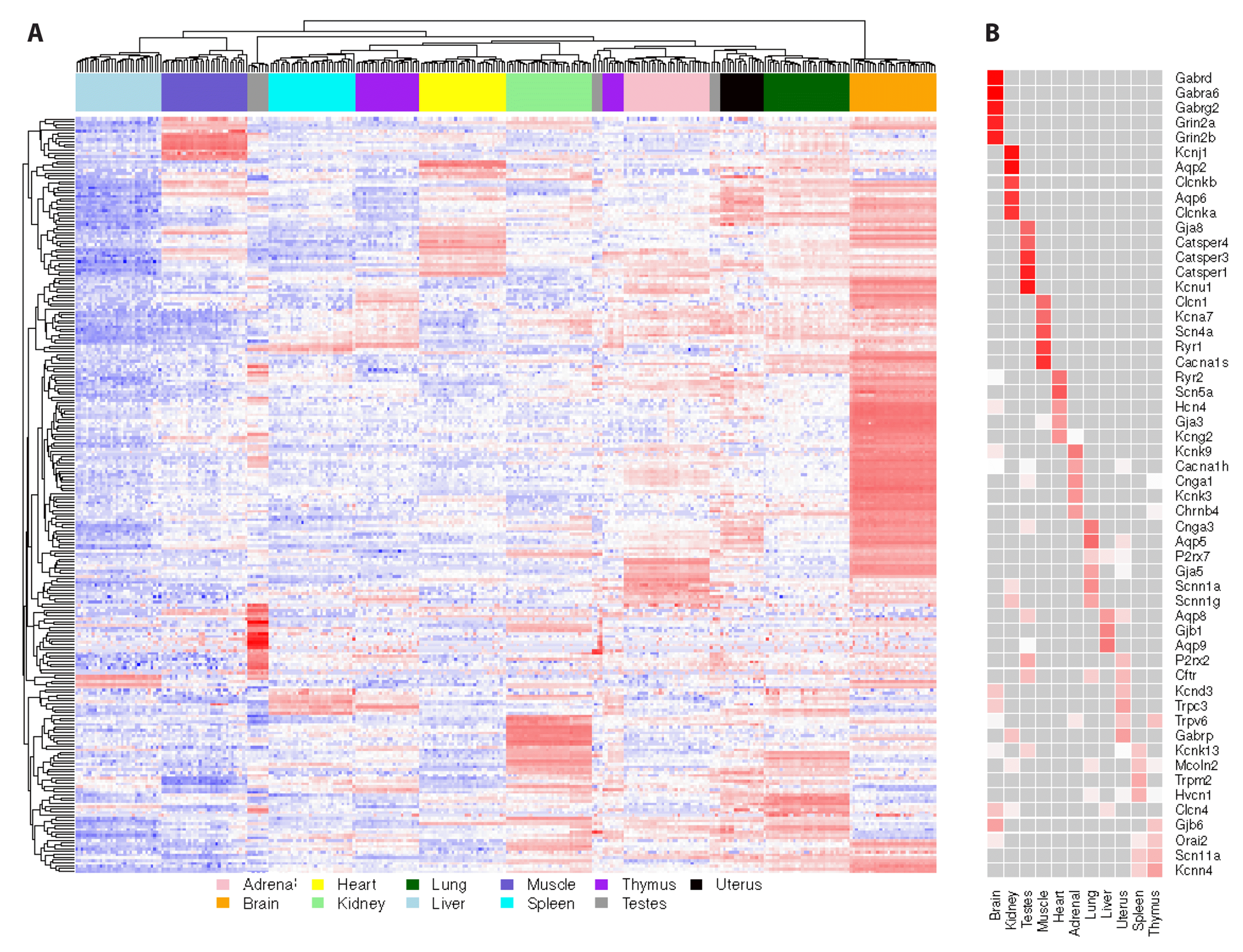
A map of tissue-specific ion channel genes
We further investigate whether the expression of the observed tissue-specific ion channel genes is relevant to the physiological function of the specific organ/tissue. A hierarchical clustering analysis was performed on the expression of all the rat ion channel genes across all the organ/tissue samples [8]. We observed that the samples from the same organ/tissue tended to be clustered into a single group except for thymus and testes. Also, each organ/tissue type presented several unique tissue-specific ion channel blocks (Fig. 3A). In addition, we found that the tissue types with similar physiological functions, such as thymus and spleen, tended to group together and share several common ion channel blocks, which suggests the common role of spleen and thymus in building the immune system (Fig. 3A). In contrast, the ion channel genes of brain samples showed a very distinct expression pattern compared with the other tissues (Fig. 3A), which largely mirrored the findings from Fig. 2A and B. To understand the functions of ion channels in different organs/tissues, we next looked into the top tissue-unique ion channel genes for each organ/tissue type. We found that the prioritized tissue-unique ion channels were related to the physiological function of each specific organ/tissue (Fig. 3B). For example, GABAA receptors and ionotropic glutamate receptors are dominant in brain, which are known to be critical for neural communication [17]; aquaporins are among the top kidney-specific ion channels, which facilitate renal water transportation through cell membrane [18]; Catsper channels are exclusively expressed in testes, which are actively involved in spermatogenesis [19] (Fig. 3B). Taken together, the above observations suggest that the tissue-specific ion channel genes play a critical role to perform their physiological functions in the individual organs/tissues.
Developmental stage-dependent ion channel gene expression
We further investigated the alterations in ion channel gene expression in rat tissues at four different developmental stages (2-, 6-, 21-, and 104-week). PCA suggests that there is clearly a distinct pattern in ion channel gene expression across the four different developmental stages in all the tissue types (Fig. 4). To prioritize the ion channel genes that are longitudinally associated with age, Spearman’s rank correlation test was performed to measure the relationship between ion channel gene expression and rat age. The correlation coefficient (ρ) between expression and age and corresponding p-value were calculated for all the ion channel genes. In total, 23, 8, 19, 19, 8, 21, 22, 10, 74, 4, and 2 ion channel genes were found to be positively correlated with age, while 23, 42, 22, 26, 12, 33, 40, 13, 12, 19, and 12 ion channel genes were negatively correlated with age (Spearman’s rank correlation test: adjusted p < 0.05) in adrenal gland, brain, heart, kidney, liver, lung, muscle, spleen, thymus, testes, and uterus, respectively (Supplementary Table 1). More interestingly, thymus appeared to have more ion channel genes with large ρ values relative to the other tissues (t-test: p < 10–10; Fig. 5A), which suggests that the expression of most ion channel genes in thymus tends to monotonically increase with age (Kolmogorov–Smirnov test: p < 10-10; Fig. 5B). As an example, Fig. 5C demonstrated the top ion channel genes showing the strongest positive correlation between expression and age (ρ > 0.9) in thymus, including Aqp1, Clcn4, Hvcn1, Itpr1, Kcng2, Kcnj11, Kcnn3, and Trpm2, which potentially contribute to the development or age-related regression in thymus.
DISCUSSION
The role of ion channel impairment and/or mutation in the pathophysiology of various diseases has received increased attention. Changes in ionic flux maintain the resting potential, regulate intracellular calcium, and transmit electrical signals; therefore, altered ion channel gene expression causes cellular dysfunction, leading to various diseases [20,21]. The ion channel genes are ubiquitously expressed and selectively distributed in tissues, and their differential distribution contributes to their diverse cellular processes and physiological functions. For example, the expression of sodium/potassium/calcium exchanger 3 (NCKX3), copper-transporting ATPase 1 (ATP7A) and copper uptake protein 1 (CTR1) are the highest in the kidney, duodenum, and liver, respectively [22]. In addition, the CACNA1F gene encodes the Cav1.4 channel, which is preferentially expressed in the retina and is necessary for retinal neural transmission [23].
The age-related changes are complicated that multiple studies have reported altered cytokines, ROS, Ca2+ homeostasis, and inflammation play major role in pathologies of ion channels [24-28]. Indeed, altered splicing mRNA and under or over-expressed mRNA with aging have been reported and RNA instability, epigenetic changes, and telomere shortening, are accompanied by changes in regulation of gene expression [29-32]. Dynamic RNA modification in gene expression regulation would be associated with altered ion channel expression with aging. The age-related changes of ion channel expression or function have been identified in human and mouse [33,34]. For example, dysregulation of Ca2+ channels have been identified in neurodegenerative disease like an Alzheimer’s disease and lower expression of Ca2+-activated K+ channels have been seen with aging in human [35,36]. However, as far as we know, limited research has been done using transcriptomic approach to understand aging related ion channel expression in mouse or human.
To our knowledge, we are among the first to carry out the comprehensive assessment of the transcriptional profile of ion channel genes in different organs/tissues and developmental stages. We found a stronger tissue-specific expression pattern of ion channel genes. In particular, five genes: including Gabrd, Gabra6, Gabrg2, Grin2a, and Grin2b, were enriched in the brain. GABAA receptors are a class of ligand-gated Cl- channels that mediate inhibitory neurotransmitters in the central nervous system. The GABAA receptor is expressed exclusively in the cerebellum, but also expressed in testis and CD4+ T cells [37,38]. Among the GABAA receptor subtypes, we found that Gabra6, Gabrd, and Gabrg2 were highly specific to the brain. The disruption of GABA transmission has been associated with different neurodegenerative diseases. Mutations in Gabra6, Gabrd, and Gabrg2 have been identified in patients susceptible to febrile seizures and genetic epilepsy [39-41]. In addition, Gabrd is a potential prognostic marker in cancer, as it shows lower expression in isocitrate dehydrogenase (IDH) wild-type tumors than in IDH mutant diffuse low-grade glioma [42], and upregulated expression in hepatocellular carcinoma and adenocarcinoma [43,44]. Both Grin2A and Grin2B, which encode the NMDA receptor subunits, cause variable neurodevelopmental phenotypes. Grin2A variants are predominantly associated with epilepsy, while Grin2B variants are commonly found in patients with developmental disorders [45,46].
Abnormal changes in ionic gradients and calcium homeostasis across the membrane can underlie age-related deterioration of cellular function. The age-related dysfunction of ion channels is known to impact organ failure, including the heart and brain [47-49]. For example, increases in NaV1.5, Navβ1, and CaV1.2, and decreases in Kv1.5 and HCN1 were seen in the sino-atrial node of the heart in old rats, and resulted in age-dependent impairment of cardiac function [2]. Age is also the strongest risk factor for neurodegenerative pathogenesis, and was associated with downregulated Cav3.1 T-type calcium channel and impaired calcium homeostasis in Alzheimer’s disease [50,51]. In addition, transcriptomic alterations that impact neurovascular function and neurodegenerative conditions were identified in endothelial cells of aged mouse brain compared with young adult mouse brain [5].
One of the progressive, age-related declines the immune system is the regression of the thymus, which is represented by a decrease in size and impaired function [52]. The thymus is the primary lymphoid organ where T cells develop and recognize antigens. Disrupted thymic architecture, with reduced medullary size and impaired epithelial cells, was observed in the aged thymus [53,54]. In addition, microarray analyses performed on thymus from young, middle-aged, and old mice identified age-associated transcriptional changes [55]. Notably, in our transcriptomic profile of ion channel genes, age-dependent changes in expression of ion channel genes, including Aqp1, Clcn4, Hvcn1, Itpr1, Kcng2, Kcnj11, Kcnn3, and Trpm2 were observed in the thymus. Potassium channels, Kcng2, Kcnj11, and Kcnn3, allow K+ flux and are essential for regulating cellular excitability, and the critical role of these ion channels in disease has been highlighted. For example, the age-associated elevated expression of Kcnn3, small-conductance calcium-activated potassium (SK3) channels in the hippocampus of aged mice resulted in learning and memory deficits [56,57]. The Kcnj11 gene and Kir6.2 subunit of the adenosine triphosphate-sensitive potassium (KATP) channel is considered a promising candidate for identifying type2 diabetes risk because Kcnj11 is crucial for insulin secretion [58]. Aquaporin-1 (AQT1) channels in the major intrinsic protein family mediate water movement across the membrane and are expressed in thymocytes, where they play a critical role in decreasing apoptotic volume [59]. Decreased expression of Itpr1, which encodes the inositol-triphophate receptor type, was identified in aged human skeletal muscle [60]. Although an age-dependent change in ion channel gene expression was observed in the thymus, there is still limited knowledge about ion channel function associated with aged thymus.
In summary, we have developed a comprehensive platform for studying the rat transcriptome that provides tissue-specific and age-dependent ion channel gene expression information. The expression of several ion channels is highly tissue-specific, supporting the idea that a therapeutic target for a specific ion channel should take into account a particular organ. Further studies are required to investigate and validate these findings.
SUPPLEMENTARY MATERIALS
Supplementary data including one table and two figures can be found with this article online at https://doi.org/10.4196/kjpp.2023.27.1.85.
 XML Download
XML Download