

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Graves’ disease (GD) is an autoimmune thyroid disorder that results in overproduction of thyroid hormones. GD-mediated thyrotoxicosis increases resting energy expenditure, body temperature, and heart rate [1]. Patients with GD usually exhibit accelerated whole-body catabolism due to increased oxygen consumption and thermogenesis, resulting in weight loss [2,3]. Moreover, an excess of thyroid hormones induces lipolysis and gluconeogenesis as well as a reduction of cholesterol levels [4-6]. More specifically, expression and activity of carnitine palmitoyl transferase 1, a key regulatory enzyme for fatty acid oxidation, are high in livers with hyperthyroidism [7]. Triiodothyronine (T3) is also involved in mitochondrial biogenesis and tricarboxylic acid cycle activity [8]. However, it is unclear which metabolites are altered upon methimazole treatment in patients with GD.
Metabolomics has the potential to identify diagnostic biomarkers in order to elucidate the mechanisms underlying diseases. Untargeted metabolomics provides a global profile of information through the simultaneous identification of as many metabolites as possible in specimens, which facilities better understanding of biochemical pathways associated with disease relapse and remission. Multiple studies have been conducted of metabolomic changes in patients with GD. Previous studies indicated that acylcarnitine and polyamine profiles differ between GD patients and the healthy population [9-12]. Hyperthyroidism had a significant impact on aminoacyl-transfer ribonucleic acid (tRNA) biosynthesis, purine and pyrimidine metabolism, and metabolism of several amino acids [12]. Moreover, remarkable changes of arginine and proline metabolic pathways are observed in adult patients with GD. Serum levels of phenylalanine and tyrosine are remarkably increased in pediatric, but not adult, patients with GD [13]. However, longitudinal analysis of changes in plasma metabolites before and after methimazole treatment has not been conducted.
In the present study, we investigated whether specific plasma metabolites were associated with recovery of euthyroidism in patients with GD using untargeted metabolomics analyzed by liquid chromatography (LC)-mass spectrometry (MS). We also aimed to identify remarkable excess thyroid hormone-mediated changes of key metabolites according to age. Furthermore, we studied whether restoration of euthyroidism by methimazole treatment altered systemic metabolic pathways in patients with GD.
METHODS
Study design and participants
Korean patients with newly diagnosed or relapsed GD who visited the Department of Internal Medicine, Chungnam National University Hospital in Daejeon between January 2019 and July 2020 were recruited. All patients were treated with methimazole for 12 weeks to achieve euthyroidism. Plasma samples were prepared from all 18 patients at the initial visit and 12 weeks later at a follow-up visit. We selected treatment period for 12 weeks. Because we previously published a paper about thyrotoxic myopathy in patient with GD [14]. In that paper, we found that skeletal muscle function and thyroid hormone levels were recovered in most GD patients treated with methimazole for 12 weeks.
Ethics statement
This study was approved by the Institutional Review Board of Chungnam National University Hospital (CNUH-2019-02-012). Each participant provided consent and documented by the Department of International Medicine of Chungnam National University Hospital in Korea.
Sample preparation for plasma metabolomics
Details have been described previously [14]. Briefly, for whole metabolite extraction, 10 µL of plasma was added to 240 µL of water and 250 µL of ice-cold methanol before being vortexed and centrifuged (14,000 ×g, 4°C, 15 minutes). The supernatant was collected in a 1.5 mL Eppendorf microtube, processed for the extraction of various types of compounds listed below, and used for LC-MS measurement. For water-soluble metabolites, including amino acids and nucleotides, 25 µL of the supernatant was diluted 3-fold with 0.1% formic acid. For acylcarnitines, 30 µL of the supernatant was added to 270 µL of ice-cold methanol, vortexed, sonicated, and centrifuged (14,000 ×g, 4°C, 15 minutes), and the supernatant was collected. For free fatty acids (FFAs), 25 µL of the supernatant was diluted 2-fold with ice-cold methanol. For bile acids, 50 µL of the supernatant was evaporated and dissolved with 25 µL of 20% methanol. For phospholipids, 5 µL of the supernatant was diluted 200-fold with 0.1% formic acid in 20% acetonitrile.
LC-MS measurements
Plasma samples were analyzed using LC-MS on a LCMS-8060 instrument (Shimadzu, Kyoto, Japan), essentially as described previously [14]. To measure wide varieties of water-soluble metabolites, the prepared sample was separated on a Discovery HS-F5-3 column (150×2.1 mm, 3 μm particle size; Sigma-Aldrich, St. Louis, MO, USA) with mobile phases consisting of solvent A (0.1% formic acid) and solvent B (0.1% formic acid in acetonitrile). The column oven temperature was 40°C. The gradient elution program was as follows with a flow rate of 0.25 mL/min: 0–2 minutes, 0% B; 2–5 minutes, 0%–25% B; 5–11 minutes, 25%–35% B; 11–15 minutes, 35%–95% B; 15–25 minutes, 95% B; and 25.1–30 minutes, 0% B. The parameters for the heated electrospray ionization (ESI) source in negative/positive ion mode under multiple reaction monitoring (MRM) were as follows: drying gas flow rate, 10 L/min; nebulizer gas flow rate, 3 L/min; heating gas flow rate, 10 L/min; interface temperature, 300°C; desolvation line (DL) temperature, 250°C; heat block temperature, 400°C; and collision-induced dissociation (CID) gas, 270 kPa. For acylcarnitines, the sample was separated on a Luna HILIC column 200A (150×2.0 mm, 3 μm; Phenomenex, Torrance, CA, USA) with mobile phases consisting of solvent A (10 mM ammonium formate) and solvent B (acetonitrile/10 mM ammonium formate [9/1, v/v]). The column oven temperature was 40°C. The gradient elution program was as follows with a flow rate of 0.3 mL/min: 0–2.5 minutes, 100% B; 2.5–4 minutes, 100%–50% B; 4–7.5 minutes, 50%–5% B; 7.5–10 minutes, 5% B; and 10.1–12.5 minutes, 100% B. Acylcarnitine profiles were detected in positive ESI mode under precursor ion scan for m/z 85.5 by changing collision energy according to the lengths of fatty acids as follows: –20 for shortchain acylcarnitines (C0–C8); –35 for middle-chain acylcarnitines (C9–C12); and –45 for long-chain acylcarnitines (C12–C18). For FFAs, the sample was separated on a ACQUITY BEH Amide column (150×2.1 mm, 1.7 μm; Waters, Milford, MA, USA) with mobile phases consisting of solvent A (10 mM ammonium formate in 90% acetonitrile) and solvent B (0.1% formic acid in acetonitrile). The column oven temperature was 40°C. The gradient elution program was as follows with a flow rate of 0.4 mL/min: 0–5 minutes, 0% B; 5.1–7.5 minutes, 100% B; and 7.6–11 minutes, 0% B. FFA profiles were detected in negative ESI mode under selected ion monitoring. For bile acids, the sample was separated on a ACQUITY BEH Amide column (150×2.1 mm, 1.7 μm; Waters) with mobile phases consisting of solvent A (0.1% formic acid) and solvent B (acetonitrile). The column oven temperature was 50°C. The gradient elution program was as follows with a flow rate of 0.3 mL/min: 0–2 minutes, 20% B; 2–10 minutes, 80% B; 10–12 minutes, 80% B; and 12.1–15 minutes, 20% B. Bile acid profiles were detected in negative ESI mode under MRM. For phospholipids, the prepared sample was separated on a Kinetex C8 column (150×2.1 mm, 2.6 μm particle size; Phenomenex) with mobile phases consisting of solvent A (20 mM ammonium formate) and solvent B (acetonitrile/isopropanol [1:1, v/v]). The column oven temperature was 45°C. The gradient elution program was as follows with a flow rate of 0.3 mL/min: 0–1 minutes, 20% B; 1–2 minutes, 40% B; 2–25 minutes, 92.5% B; 25.1–35 minutes, 100% B; and 35.1–38 minutes, 20% B. Phospholipid profiles were detected in positive ESI mode under MRM.
Metabolomics data processing and analysis
Data processing was performed using the LabSolutions LC-MS software program (Shimadzu).
Biochemical measurements
Peripheral blood was collected in heparin-coated tubes. Plasma levels of thyroid-stimulating hormone (TSH), T3, free thyroxine (T4), and TSH-binding inhibitor immunoglobulin (TBII) were measured by standard methods on an automated analyzer (Cobas 6000, Roche Diagnostics GmbH, Mannheim, Germany).
Principal component analysis and metabolic pathway analysis of metabolomics data
MetaboAnalyst was used for pathway analysis and performs metabolite set enrichment analysis, which includes a set of human and mammalian metabolites and a set of chemical grade metabolites. This module accepts a list of compound names, a list of compound names with concentrations, or a table of concentrations. The analysis is based on multiple libraries containing a set of approximately 9,000 biologically significant metabolites collected primarily from human studies, including more than 1,500 chemical classes. A portion of the data utilized for analysis could be analyzed because it was matched to the MetaboAnalyst database. The R program’s FactoMineR function was used to perform principal component analysis (PCA).
Statistical analysis
The analysis method was selected after proving the normalcy of the data in accordance with the distribution of each variable. Using the details of statistical tests included in the plot of ggstatsplot, an extension of ggplot2, violin plots could be created. The Student’s t test (two-sided) was used to compare variables between groups (before and after treatment) after assessing the assumption of normality. The Wilcoxon signed-rank test was used to compare paired variables between groups. Statistical analyses were performed using R software version 4.1.0 (R Foundation for Statistical Computing, Vienna, Austria). P<0.05 was considered significant.
RESULTS
Characteristics of the study participants
The demographic and clinical characteristics of 18 patients with GD (male 33%, female 67%) at the initial visit are showed in Table 1. Twelve weeks after methimazole treatment, the thyroid hormone levels of all patients were within the normal range and were significantly different from those before treatment (Fig. 1A). The mean free T4, T3, TSH, and TBII levels of GD patients were similar in males and females (Fig. 1B). The age of the study participants was 22 to 64 years. Among all the measured metabolites, only nine plasma metabolites, including uracil, linolenic acid (C18:3), lysophosphatidylcholine (20:5), and cysteine, had positive correlations (P<0.05) with age in drug-naïve patients with GD. On the other hand, only palmitic acid (C16:0) had a negative correlation with age (Fig. 2).
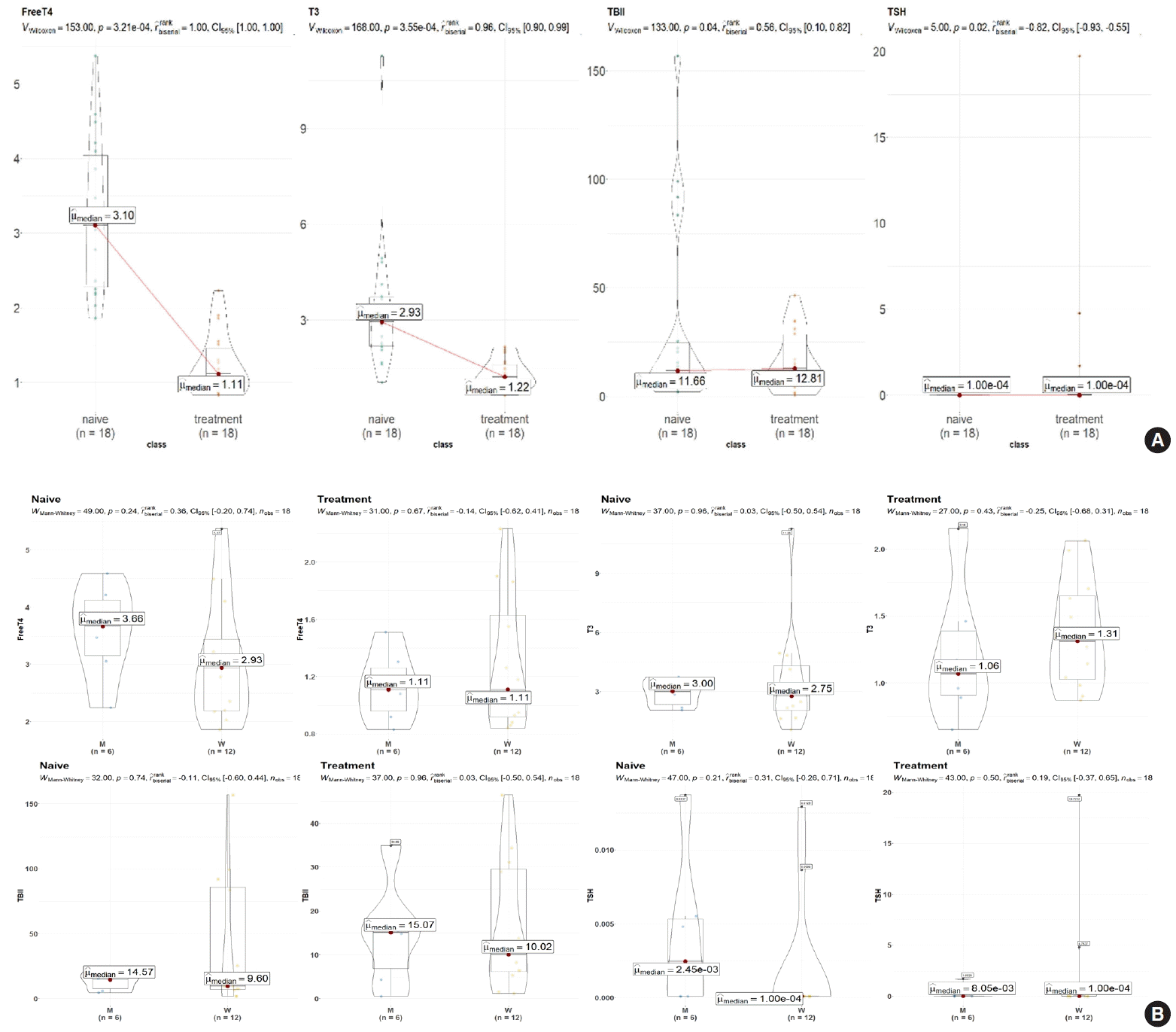
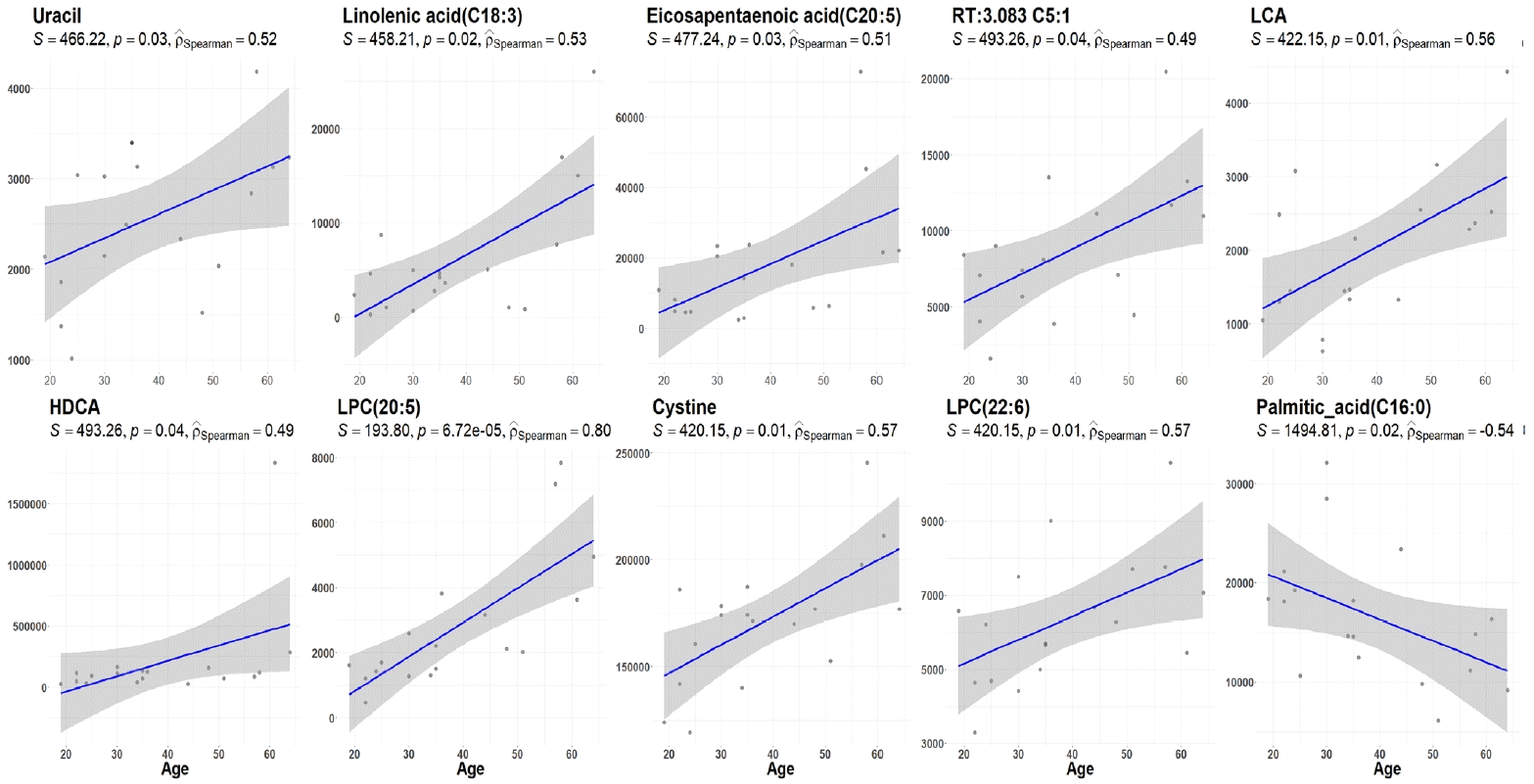
Fig. 1.
Serum levels of thyroid hormones and thyroid-stimulating hormone (TSH)-binding inhibitor immunoglobulin (TBII) in patients with Graves’ disease at weeks 0 and 12 after commencement of methimazole treatment. (A) Changes of thyroid hormone levels before and after treatment of Graves’ disease with methimazole. (B) Levels of thyroid hormones, TBII, and TSH in male and female patients with Graves’ disease. Parametric (paired Student’s t test) and nonparametric (Wilcoxon signed-rank test) tests were used for statistical analysis. T4, thyroxine; T3, triiodothyronine; CI, confidence interval.
![]()
Fig. 2.
Plasma levels of metabolites that show remarkable age-related increases or decreases in methimazole treatment-naïve patients with Graves’ disease. Aging-related metabolite changes in Graves’ disease patients were evaluated by Spearman’s correlation analysis. RT, retention time; LCA, lithocholic acid; HDCA, hyodeoxycholic acid; LPC, lysophosphatidylcholine.
![]()
Table 1.
Clinical and Biochemical Characteristics of the Study Subjects at the Initial Visit
![]()
Differential metabolomics analysis before and after 12 weeks of methimazole treatment in GD patients
Two hundred metabolites were measured in plasma samples of all participants with GD before and after methimazole treatment (Supplemental Table S1). PCA score plots showed separate clustering before and after 12 weeks of methimazole treatment in patients with GD (Fig. 3A). Creatine, stearic acid (C18:0), and palmitic acid (C16:0) were enriched in plasma of methimazole-naïve patients with GD. On the other hand, many LPC family members, 2-ketoglutaric acid, and nervonic acid (C24:1) were enriched after methimazole treatment (Fig. 3B).
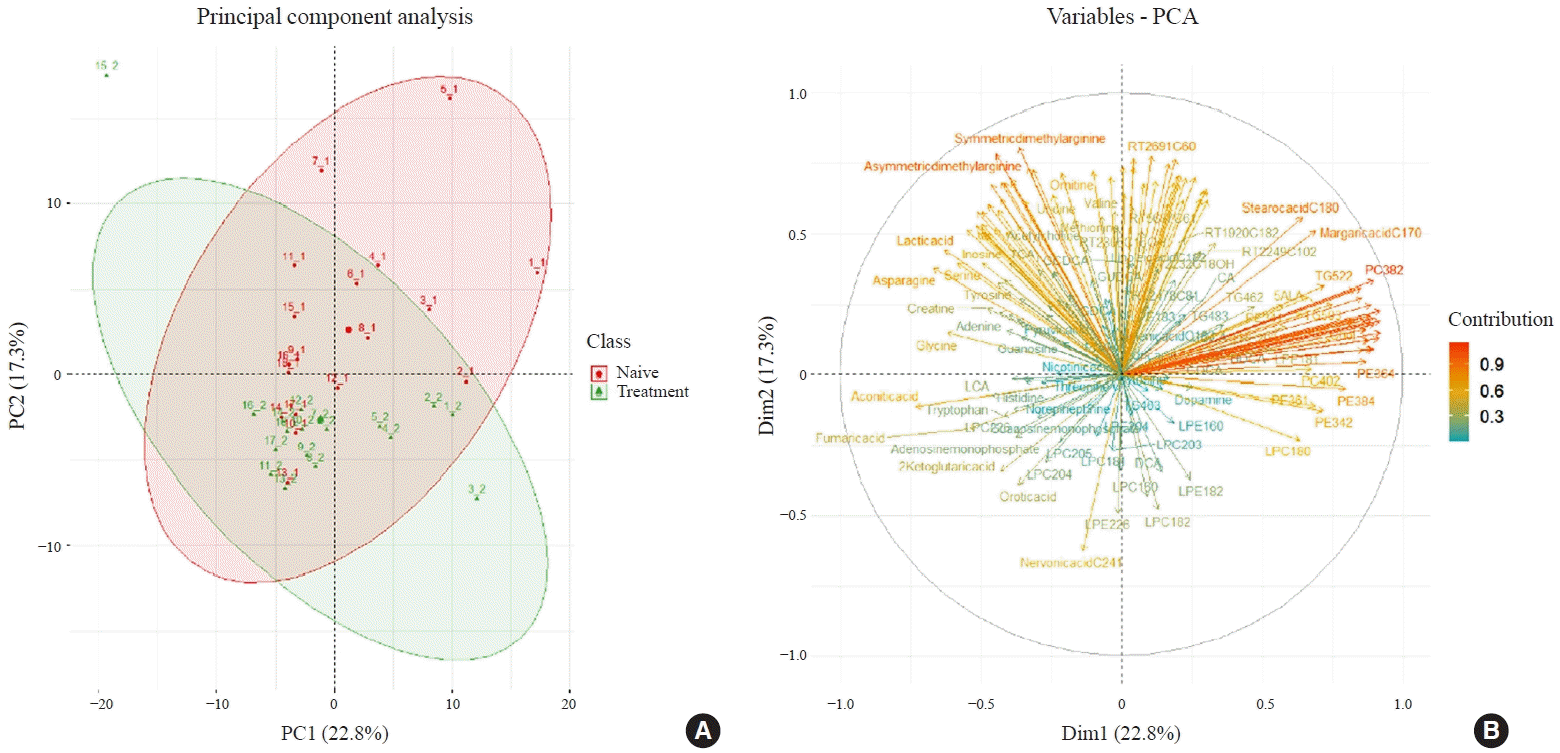
Fig. 3.
Principal component analysis (PCA) of metabolite data from patients with Graves’ disease at weeks 0 and 12 after commencement of methimazole treatment. (A) PCA of metabolite data from patients with Graves’ disease at weeks 0 and 12. The numbers in parentheses show the contribution rates. The red dots indicate week 0 (n=18) and the green dots indicate week 12 (n=18). (B) The loadings of individual metabolites on the principal components shown in (A).
![]()
The levels of only 76 plasma metabolites significantly differed before and after methimazole treatment. The levels of 61 metabolites were increased in drug-naïve patients with GD and the levels of 15 metabolites were increased after methimazole treatment. The levels of fatty acids including palmitic acid (C16:0), stearic acid (C18:0), carnitine, and oleic acid (C18:1), and amino acids including dimethylglycine, cysteine, and valine were elevated in patients with GD at the initial visit, but they were remarkably decreased upon methimazole treatment (Fig. 4). By contrast, the levels of nervonic acid (C24:1), LPC family members including LPC (16:0), (18:0), and (18:1), glycine, and creatinine were significantly increased upon methimazole treatment (Fig. 4 and Supplemental Fig. S1).
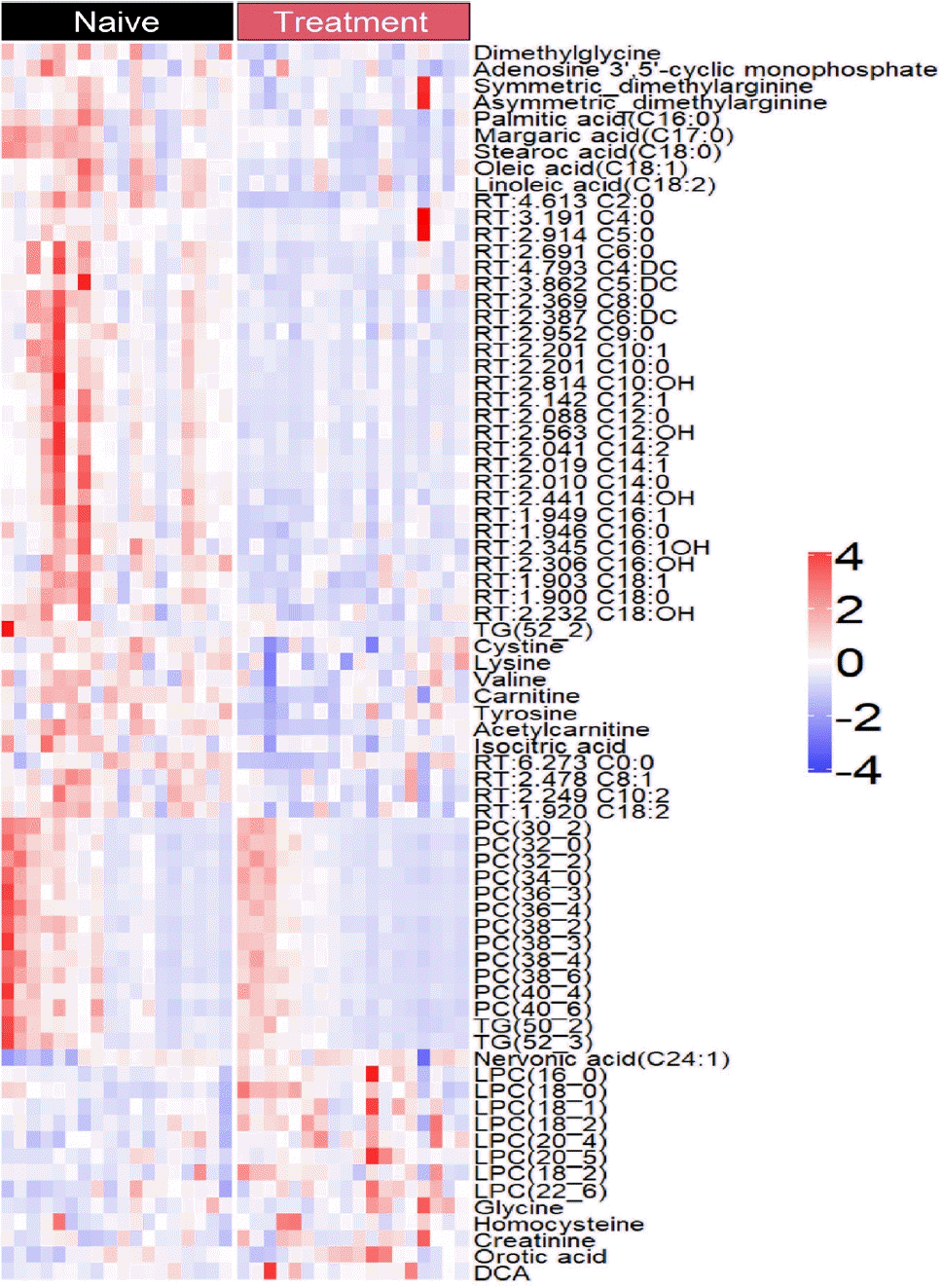
Fig. 4.
Plasma levels of metabolites before and after 12 weeks of methimazole treatment in patients with Graves’ disease. Visualization of significantly differentiated plasma metabolomic data (200 metabolites) between methimazole treatment with Graves’ disease in a heatmap. RT, retention time; TG, triglyceride; PC, phosphatidylcholine; LPC, lysophosphatidylcholine; DCA, dichloroacetic acid.
![]()
Pathways that differ between GD patients with thyrotoxicosis and euthyroidism
We observed changes of plasma metabolites before and after methimazole treatment in GD patients. To identify changes in metabolic pathways, we investigated which metabolic pathways differed between the initial visit and 12-week follow-up visit using pathway enrichment analysis. The aminoacyl-tRNA biosynthesis pathway was most significantly altered (Fig. 5). Moreover, various amino acid metabolic pathways, including valine, leucine, and isoleucine biosynthesis and glycine, serine, and threonine metabolism, differed before and after methimazole treatment in patients with GD (Fig. 5). In addition, phenylalanine, tyrosine, and tryptophan biosynthesis- and cysteine and methionine metabolism-related metabolite sets significantly differed between the initial visit and 12-week follow-up visit (Fig. 4B).
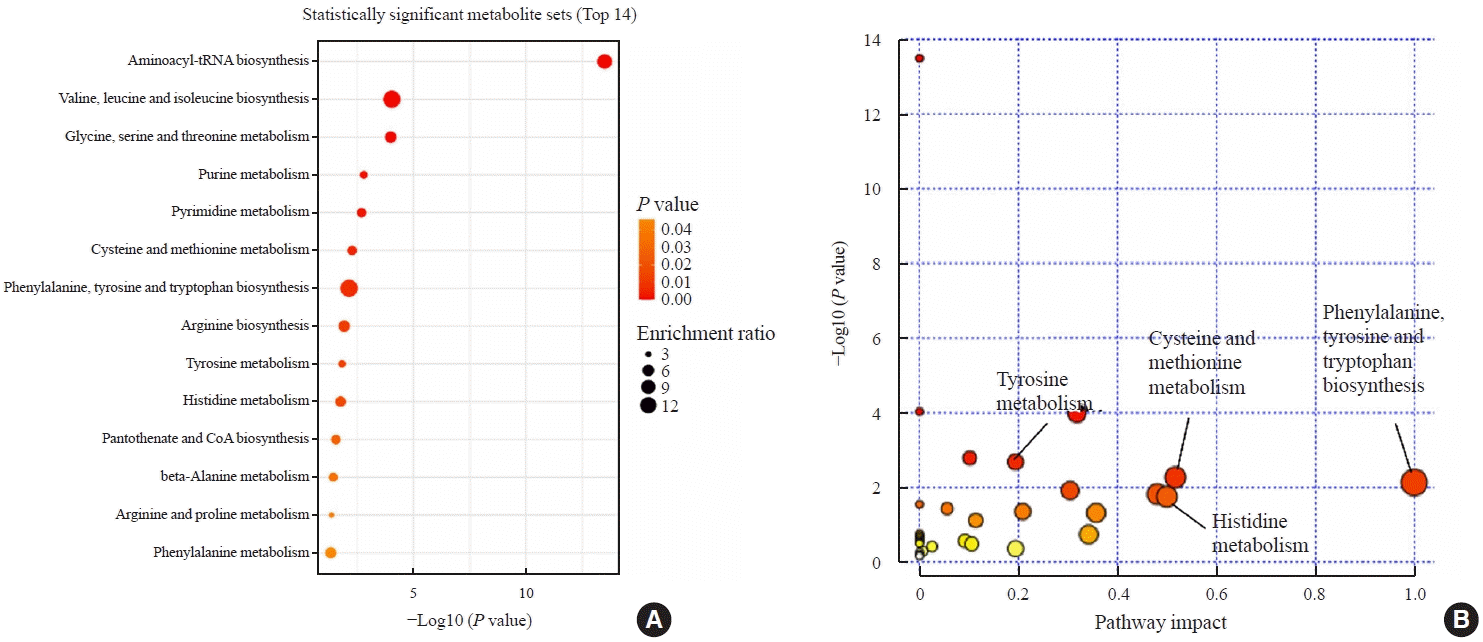
Fig. 5.
Pathway analysis of plasma metabolite profiles before methimazole treatment in patients with Graves’ disease. (A) Metabolite set enrichment analysis based on the altered metabolites. (B) Serum metabolites based on pathway analysis before treatment in patients with Graves’ disease. CoA, coenzyme A.
![]()
DISCUSSION
This study provides an overview of alterations of plasma metabolites in GD patients according to age and 12 weeks of methimazole treatment. The levels of 200 plasma metabolites, including amino acids, FFAs, bile acids, and lipids, were measured in patients with GD before and after methimazole treatment. The levels of 76 metabolites significantly differed before and after treatment. Of these, the levels of 61 (80.2%) metabolites were increased in drug-naïve patients with hyperthyroidism and the levels of 15 metabolites (19.8%) were higher in the recovery phase after methimazole treatment. An excess of thyroid hormones induced significant changes of numerous amino acid biosynthesis pathways, especially the aminoacyl-tRNA biosynthesis pathway, which were stabilized upon recovery of euthyroidism. We also found that metabolites varied according to age in GD patients (Fig. 2). A previous report showed that many metabolites including arginine, carnitine, and β-hydroxybutyrate are altered by aging [15], but we found that some metabolites including uracil, LPC family members, and palmitic acid were only changed when thyroid hormones were in excess. A further study is warranted to determine not only the biological meaning of the metabolite changes but also to reveal the roles of each enriched metabolite in GD patients.
Several studies have investigated the role of thyroid hormones in systemic lipid metabolism. Thyroid hormones regulate lipogenesis and lipolysis [16]. Overproduction of thyroid hormones stimulates systemic lipolysis, elevating the circulating FFA level [17]. The current study shows that overproduction of thyroid hormones increases the levels of fatty acids of different lengths, including acetylcarnitine and acylcarnitine, which are involved in the conversion of fat into energy [18]. Therefore, systemic lipolysis and mitochondrial β-oxidation are stimulated by overproduction of thyroid hormones and decreased by methimazole treatment. Another publication indicated that patients with hyperthyroidism have an increased serum concentration of FFAs and treatment with antithyroid drugs revealed a direct relationship between thyroid activity and the FFA concentration [19]. Our data are consistent with an inverse relationship between thyroid activity and the percentage levels of palmitic acid in FFAs. In a cross-sectional study, Zhou et al. [20] examined the relationship between serum palmitic acid and thyroid function in the United States population using the database of the National Health and Nutrition Examination Survey from 2011 to 2012. They identified a significant negative association between serum palmitic acid and free T4 [20]. In our study, methimazole-naïve patients had an elevated serum level of palmitic acid and this was decreased after methimazole treatment. This discrepancy is thought to originate from a difference in study design. Alterations in serum thyroid hormones are associated with aging [21,22]. In the European population, Hoogendoorn et al. [23] reported that the serum free T4 concentrations increases with age in individuals older than 60 years. These data seem to be consistent with our finding that the serum palmitic acid decreased with age and methimazole treatment in Korean patients. However, another study from Western Australia found no significant change in the free T4 level with aging [21].
Amino acids are organic compounds involved in protein synthesis and protein-protein interactions, which are essential for the composition of muscle and other tissues [24,25]. Branched-chain amino acids (BCAAs), valine, leucine, and isoleucine, are essential amino acids and serve as substrates for inhibition of proteolysis, protein synthesis, glucose metabolism, and energy production [26,27]. In the present study, various amino acid metabolic pathways differed before and after 12 weeks of methimazole treatment in patients with GD. Levels of valine, leucine, and arginine have significant positive correlations with serum levels of T4 and free T4 [28]. Consistently, valine and leucine biosynthesis was reduced after 12 weeks of treatment in the current study. BCAAs, valine, leucine, and isoleucine have been well studied in the context of muscle protein recovery [29]. The elevation of BCAAs in GD patients is thought to be due to their release from skeletal muscle, which is the initial catabolism site of BCAAs due to high expression of BCAA aminotransferase, the first enzyme in the pathway of BCAA catabolism [27,30]. GD patients exhibit muscle loss and weakness; therefore, the elevation of BCAAs via valine, leucine, and isoleucine biosynthesis is a compensatory effect. Thus, to overcome muscle myopathy in GD patients, there is a compensatory effect to improve muscle function through elevation of BCAA biosynthesis. Glycine and serine were reported to inhibit TSH secretion in rats by enhancing the activity of the periventricular hypothalamus and pituitary gland [31]. The elevated level of glycine after methimazole treatment in our study may reflect the function of glycine in the transition from hyperthyroidism to euthyroidism.
The main strength of this study is that it acquired serial data from LC-MS-based plasma metabolomics to make findings that may have clinical value for understanding the pathophysiology of GD. However, the present study has several limitations. First and most importantly, as an observational study, it cannot determine a causal association between variables. Second, the study population was exclusively South Korean and therefore the results may not be applicable to other ethnicities with GD. Third, a variety of confounding factors could not be considered in multivariate analyses due to the small sample size. Fourth, the follow-up period of 12 weeks was too short to fully assess euthyroid-induced recovery of metabolic networks. Longer serial follow-up may provide additional information about the long-term systemic metabolic effects of hyperthyroidism. Lastly, the lack of dynamic information precluded interpretation of metabolomics data in the context of metabolic fluxes. It is difficult to determine whether our findings are the result of increased flux from a synthesizing enzyme, decreased flux toward a consuming enzyme, or alteration in transport of a metabolite into or out of cells or tissues.
In conclusion, this study reports comprehensive metabolite changes that occur during the transition from hyperthyroidism to euthyroidism, and provides new insights into systemic energy homeostasis and metabolic processes, which are beneficial for understanding the clinical features and management of patients with GD.
 XML Download
XML Download