

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Numerous studies have shown that oxidative stress plays a causal role in initiating inflammation [12]. Inflammation can cause various diseases, such as metabolic disease [3]. Thus, the suppression of oxidative stress is a useful target for preventing inflammatory diseases.
Inflammation can result from oxidative damage to lipids [4]. As a major end product of lipid peroxidation, 4-hydroxy-2-nonenal (HNE) can stimulate the expression of cyclooxygenase-2 (COX-2), which is primarily responsible for inflammation [56]. Moreover, recent progress has identified the involvement of HNE in the nuclear factor-κB (NF-κB) signaling pathway [78], which is a master regulator in the inflammatory process [91011121314]. Therefore, HNE is a biomarker for oxidative stress and an important inducer of inflammation.
Dietary antioxidants can suppress oxidative stress to decrease the formation of HNE, leading to a better understanding of how the dietary antioxidant plays a role in preventing HNE-derived inflammation. Methionine (Met) is an essential sulfur-containing amino acid that can regulate oxidative stress and lipid metabolism [151617181920]. Wang et al. [17] reported that methionine could activate nuclear factor erythroid 2-related factor 2 (Nrf2)-antioxidant responsive element (ARE) pathway to increase intracellular antioxidant (glutathione, GSH) synthesis and stimulate the methionine sulfoxide reductase antioxidant system, resulting in an endogenous antioxidant response to suppress oxidative stress. On the other hand, the evidence on whether methionine can depress HNE-derived inflammation is lacking. In particular, the link between the regulation of the NF-κB signaling pathway and methionine intake is unclear.
Accordingly, this study examined the regulatory effects of methionine on HNE-derived inflammation. The following key questions were addressed: (1) whether there is a link between the anti-inflammatory action and the methionine consumption in growing rats, and (2) whether and how methionine can improve HNE-derived inflammation by modulating the NF-κB signaling pathway.
Go to : 
MATERIALS AND METHODS
Animal experiment
The animal experiment followed the same procedure as that reported elsewhere [17], and was approved by the Animal Experimental Ethics Committee of the Harbin Institute of Technology (IACUC-2020009). All animal studies used in the present work were performed according to the Guidelines of the Committee for the Experimental Animals of Harbin Institute of Technology (Harbin, China).
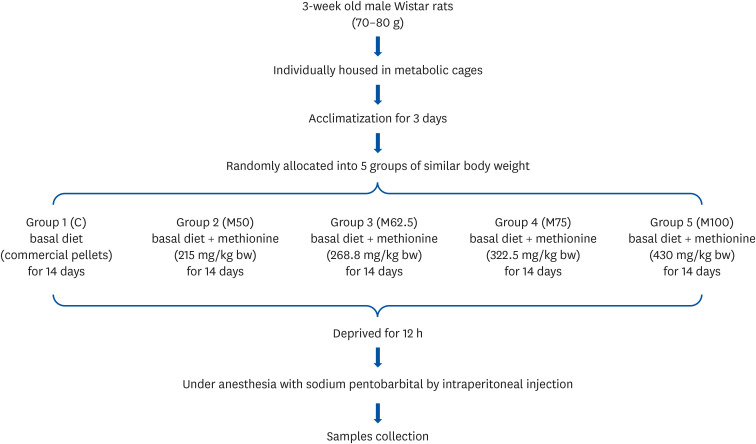
Male Wistar rats (three-week-old weighing 70–80 g) were purchased from the Vital River Laboratory (Beijing Vital River Laboratory Animal Technology Co. Ltd., Beijing, China) and housed in individual stainless steel metabolic cages, at a temperature of 22 ± 2°C and under a 12 h light/dark cycle (07:00–19:00 for light). The rats were allowed free access to commercial pellets (Beijing Vital River Laboratory Animal Technology Co. Ltd.) for three days. After acclimatization, the rats were allocated randomly into five groups with a similar body weight (bw). Each group consisted of six animals. All animals were fed commercial pellets ad libitum, which were served as the basal diets.
Group 1 (C) was fed the basal diet without any oral administration and treated as the control. Groups 2 (M50), 3 (M62.5), 4 (M75), and 5 (M100) were fed the basal diet and administered L-methionine orally at 215.0 mg/kg bw, 268.8 mg/kg bw, 322.5 mg/kg bw, and 430.0 mg/kg bw, respectively, which corresponded to 50%, 62.5%, 75%, and 100% of the standard level according to AIN-93G [21]. All animals were orally administered M50, M62.5, M75, and M100 daily for 14 days (Fig. 1).
Sample collection
Sample collection followed the same protocol as in previous studies [1718222324]. Briefly, at the end of the feeding period (14 days), the rats were fasted for 12 h before sample collection. Under anesthesia with sodium pentobarbital (5 mg/100 g bw) via intraperitoneal injection, all animals were sacrificed without pain. Blood was withdrawn from the abdominal vein into a heparinized syringe, cooled immediately on ice, and separated by centrifugation at 12,000 × g for 5 min. The plasma was frozen at −20°C until analysis. After blood collection, the liver was excised immediately, rinsed in saline, and weighed after being blotted on a filter paper. The liver was cut into several portions, freeze-clamped quickly in liquid nitrogen, and stored at −80°C until analysis.
Measurement of hepatic HNE content
The hepatic content of HNE after 14 days of feeding was determined using enzyme-linked immunosorbent assay (ELISA) commercial kits (Shanghai Enzyme-linked Biotechnology, Shanghai, China).
Quantitative real-time polymerase chain reaction (PCR)
Quantitative real-time PCR was performed according to previous studies [1718222324]. Briefly, the total RNA was extracted from a rat liver after 14 days of feeding using a TRIzol reagent kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. cDNA was reverse transcribed from 1 μg of the total RNA using a PrimeScript™ 1st strand cDNA Synthesis Kit (Takara Bio. Inc., Otsu, Shiga, Japan). For quantitative real-time PCR, the cDNAs were analyzed using an ABI 7500 sequence detection system (Applied Biosystems, Foster City, CA, USA) with SYBR Green (Takara Bio. Inc.). The RNA purity was determined from the A260/A280 ratio (C, 2.00; M50, 2.01; M62.5, 2.00; M75, 1.99; M100, 2.01). The result was normalized to the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA level. Table 1 lists the primer sequences used in this study. In this study, group C was a control group. Thus, the relative mRNA level in group C was set to 1.00. The mRNA levels of groups M50, M62.5, M75, and M100 were the ratio of groups M50, M62.5, M75, and M100 to group C, respectively.
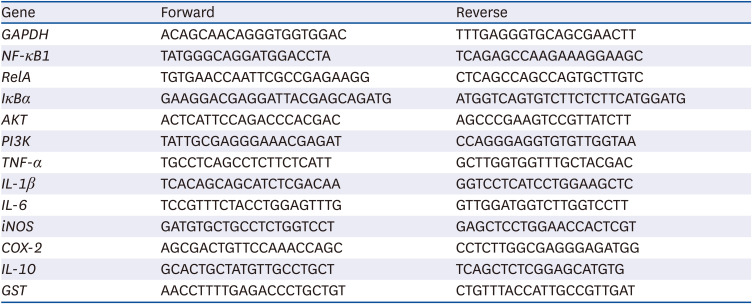
Table 1
Sequences of the primers used for quantitative real-time polymerase chain reaction
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NF-κB1, nuclear factor-κB1; RelA, reticuloendotheliosis viral oncogene homolog A; IκBα, inhibitory κBα; AKT, protein kinase B; PI3K, phosphoinositide 3 kinase; TNF-α, tumor necrotic factor alpha; IL-1β, interleukin-1β; IL-6, interleukin-6; iNOS, inducible nitric oxide synthase; COX-2, cyclooxygenase-2; IL-10, interleukin-10; GST, glutathione S-transferase.
![]()
Western blotting analysis
Western blotting analysis followed the same protocol reported elsewhere [1718222324]. Briefly, with equal amounts of protein, the samples of the total, cytoplasmic, and nuclear proteins were denatured by boiling for 5 min and separated by 10% SDS-PAGE. The gels were transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, MA, USA). After blocking with 5% fat-free milk in TBS at room temperature for 1 h, the membranes were incubated overnight at 4°C with the primary antibodies for the following: inhibitory κBα (IκBα, Proteintech, Wuhan, China), NF-κB1 (p105/p50, Proteintech), reticuloendotheliosis viral oncogene homolog A (RelA/p65, Proteintech), COX-2 (Proteintech), interleukin-1β (IL-1β, Proteintech), interleukin-6 (IL-6, Proteintech), inducible nitric oxide synthase (iNOS, Proteintech), tumor necrotic factor alpha (TNF-α, Proteintech), phosphoinositide 3 kinase (PI3K, Proteintech), protein kinase B (AKT, Proteintech), interleukin-10 (IL-10, Santa Cruz Biotechnology, Santa Cruz, CA, USA), glutathione S-transferase α1 (GSTA1, Proteintech), H1.2 (Proteintech), GAPDH (Proteintech), and β-actin (Cell Signaling, Danvers, MA, USA). The membranes were then washed three times with TBST (TBS with 0.1% Tween-20) and incubated at room temperature for 2 h with a secondary antibody (Santa Cruz Biotechnology). The protein bands were visualized using the ECL reagent (Beyotime). The amount of protein was quantified using QuantityOne software (Bio-Rad, Hercules, CA, USA). In this study, group C is a control group; thus, the relative protein expression in group C was set to 1.00. The protein expression was calculated as the ratio of groups M50, M62.5, M75, and M100 to group C, respectively.
Statistical analysis
Data are expressed as the mean ± SEM. In this study, the statistical analyses and correlation analyses were performed using the PC software package SPSS (version 23.0; SPSS Inc., Chicago, IL, USA). Statistically significant differences among the groups were analyzed via one-way analysis of variance (ANOVA) and the least significant difference test. Statistical significance was set at P < 0.05.
Go to :
RESULTS
Body weights
With similar initial body weight (C, 73.39 ± 2.60 g; M50, 73.66 ± 2.13 g; M62.5, 73.60 ± 2.02 g; M75, 73.43 ± 1.81 g; M100, 73.47 ± 1.80 g; P > 0.05), after 14 days of feeding, no significant differences of final body weight (C, 167.25 ± 4.75 g; M50, 168.78 ± 3.78 g; M62.5, 169.02 ± 2.70 g; M75, 169.03 ± 7.51 g; M100, 171.24 ± 2.52 g; P > 0.05), and body weight (C, 93.86/14 days ± 3.79 g; M50, 95.12/14 days ± 3.12 g; M62.5, 95.42 ± 4.78 g; M75, 95.60 ± 4.16 g; M100, 97.77 ± 3.48 g; P > 0.05) were found among the experimental groups.
Hepatic HNE contents
After oral administration for 14 days, the hepatic contents of HNE decreased with increasing L-methionine levels. As shown in Fig. 2, compared to C, groups M50, M62.5, M75 and M100 showed 12.92% (P < 0.05), 14.92% (P < 0.05), 17.75 (P < 0.05), and 20.23% (P < 0.05) less hepatic HNE accumulation in growing rats, respectively, showing a good dose-dependent relationship. The significant negative correlation between L-methionine intake and hepatic HNE content (r = −0.8171, P < 0.05) suggested that L-methionine could suppress lipid peroxidation.
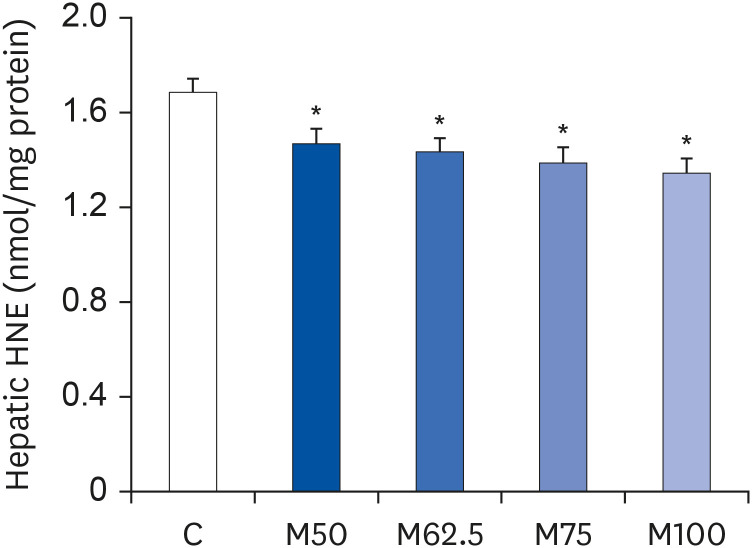 | Fig. 2Hepatic contents of HNE in growing rats. Hepatic contents of HNE in growing rats after the oral administration of L-methionine for 14 days. The values are expressed as the means ± SEM (n = 6).HNE, 4-hydroxy-2-nonenal; C, control; M50, basal diet and administered L-methionine orally at 215.0 mg/kg bw; M62.5, basal diet and administered L-methionine orally at 268.8 mg/kg bw; M75, basal diet and administered L-methionine orally at 322.5 mg/kg bw; M100, basal diet and administered L-methionine orally at 430.0 mg/kg bw.
*P < 0.05, in comparison with C.
|
Protein expression and mRNA level of COX-2
After 14 days feeding, the protein expression and mRNA level of COX-2, which was primarily responsible for inflammation, were decreased by L-methionine feeding. Compared to C, M62.5 (P < 0.05), M75 (P < 0.05) and M100 (P < 0.05) produced dramatic decreases in the hepatic protein expression of COX-2 (Fig. 3A), whereas no significant difference in the COX-2 protein level was observed between M50 and C (P > 0.05, Fig. 3A). The mRNA levels of COX-2 were reduced markedly in groups M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05) and M100 (P < 0.05) (Fig. 3B), showing a good dose-dependent manner in COX-2’s mRNA level. The results showed a significantly positive correlation between the hepatic HNE content and COX-2 expression (r = 0.8916, P < 0.05), but a significantly negative correlation between L-methionine intake and COX-2 expression (r = −0.8117, P < 0.05). Overall, these findings suggested that L-methionine suppresses HNE-induced inflammation.
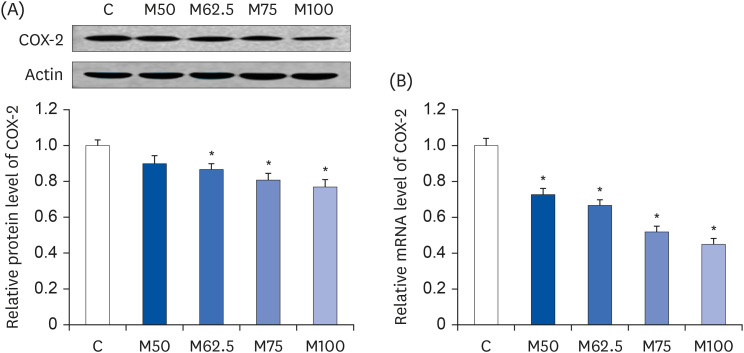 | Fig. 3Hepatic protein expression and mRNA level of COX-2 after the oral administration of L-methionine for 14 days. (A) Hepatic protein expression of COX-2. (B) Hepatic mRNA level of COX-2. The values are expressed as the means ± SEM (n = 6).COX-2, cyclooxygenase-2; C, control; M50, basal diet and administered L-methionine orally at 215.0 mg/kg bw; M62.5, basal diet and administered L-methionine orally at 268.8 mg/kg bw; M75, basal diet and administered L-methionine orally at 322.5 mg/kg bw; M100, basal diet and administered L-methionine orally at 430.0 mg/kg bw.
*P < 0.05, in comparison with C.
|
Protein expressions and mRNA levels of inflammatory mediators
After oral administration for 14 days, L-methionine regulated the protein and mRNA levels of the inflammatory mediators. As summarized in Fig. 4, the mRNA levels of IL-1β (P < 0.05, Fig. 4A), IL-6 (P < 0.05, Fig. 4B), iNOS (P < 0.05, Fig. 4C) and TNF-α (P < 0.05, Fig. 4D), which are major inflammatory mediators, were decreased markedly by L-methionine. Compared to group C, the protein expression of IL-1β (P < 0.05, Fig. 4A), IL-6 (P < 0.05, Fig. 4B), iNOS (P < 0.05, Fig. 4C), and TNF-α (P < 0.05, Fig. 4D) were depressed significantly by M62.5, M75 and M100, whereas M50 did not induce significant increases in the protein expressions of IL-1β (P > 0.05, Fig. 4A), IL-6 (P > 0.05, Fig. 4B), iNOS (P > 0.05, Fig. 4C), and TNF-α (P > 0.05, Fig. 4D) compared to C.
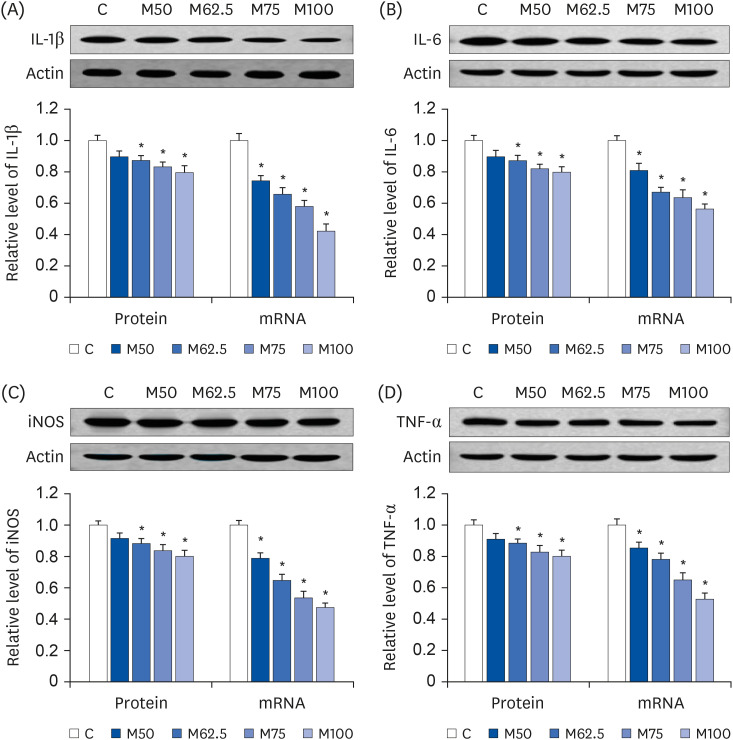 | Fig. 4Hepatic protein expression and mRNA levels of inflammatory mediators after the oral administration of L-methionine for 14 days. (A) Hepatic protein expression and mRNA levels of IL-1β. (B) Hepatic protein expressions and mRNA levels of IL-6. (C) Hepatic protein expressions and mRNA levels of iNOS. (D) Hepatic protein expressions and mRNA levels of TNF-α. Values are the means ± SEM (n = 6).IL-1β, interleukin-1β; IL-6, interleukin-6; iNOS, inducible nitric oxide synthase; TNF-α, tumor necrotic factor-alpha; C, control; M50, basal diet and administered L-methionine orally at 215.0 mg/kg bw; M62.5, basal diet and administered L-methionine orally at 268.8 mg/kg bw; M75, basal diet and administered L-methionine orally at 322.5 mg/kg bw; M100, basal diet and administered L-methionine orally at 430.0 mg/kg bw.
*P < 0.05, compared with C.
|
In this study, there were significantly positive correlations between the hepatic HNE contents and the inflammatory mediator expression of TNF-α (r = 0.8697, P < 0.05), of IL-1β (r = 0.8338, P < 0.05), of IL-6 (r = 0.8988, P < 0.05), and of iNOS (r = 0.8157, P < 0.05), confirming that HNE is an important inducer of inflammation. In contrast, the results showed significantly negative correlations between L-methionine intake and the expression of TNF-α (r = −0.8187, P < 0.05), IL-1β (r = −0.7646, P < 0.05), IL-6 (r = −0.8279, P < 0.05), and iNOS (r = −0.7566, P < 0.05), respectively. These results suggest that L-methionine could prevent HNE-derived inflammation, which is dependent on the L-methionine availability.
Protein expression and mRNA levels of NF-κB
After oral administration for 14 days, the pronounced decreases in the mRNA levels of NF-κB1 (P < 0.05, Fig. 5A) and RelA (P < 0.05, Fig. 5B), the major master regulators in the inflammatory process, were induced by L-methionine. Compared to group C, the hepatic protein levels of NF-κB1 (Fig. 5A) in groups M62.5, M75, and M100 were reduced markedly by 13.23% (P < 0.05), 18.18% (P < 0.05), and 20.63% (P < 0.05), respectively after 14 days feedings, whereas M50 showed only a 10.76% decrease (P > 0.05) in the protein expression of NF-κB1 with respect to C. As shown in Fig. 5B, dramatic decreases in RelA protein expressions were found in M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05), showing that the protein expressions of RelA were inhibited significantly by increasing the L-methionine intake. The results showed significant negative correlations between the L-methionine intake and expressions of NF-κB1 (r = −0.8027, P < 0.05) and RelA (r = −0.8187, P < 0.05).
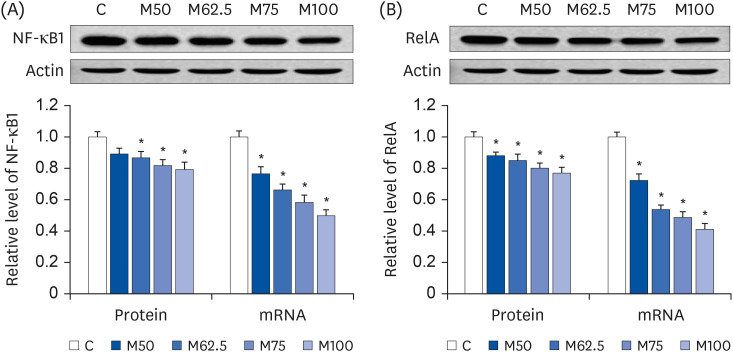 | Fig. 5Hepatic protein expressions and mRNA levels of NF-κB1 and RelA after the oral administration of L-methionine for 14 days. (A) Hepatic protein expressions and mRNA levels of NF-κB1. (B) Hepatic protein expressions and mRNA levels of RelA. The values are expressed as the means ± SEM (n = 6).NF-κB1, nuclear factor-κB1; RelA, reticuloendotheliosis viral oncogene homolog A; C, control; M50, basal diet and administered L-methionine orally at 215.0 mg/kg bw; M62.5, basal diet and administered L-methionine orally at 268.8 mg/kg bw; M75, basal diet and administered L-methionine orally at 322.5 mg/kg bw; M100, basal diet and administered L-methionine orally at 430.0 mg/kg bw.
*P < 0.05, compared with C.
|
Activation of NF-κB
This study examined the effects of L-methionine on the NF-κB signaling pathway after 14 days of feeding. With the intake of L-methionine, the protein expression (Fig. 6A) and mRNA level (Fig. 6B) of IκBα, which was an inhibitor of NF-κB, was increased gradually by L-methionine administration in growing rats. Compared to group C, both protein expression (Fig. 6A) and mRNA level (Fig. 6B) of IκBα were increased markedly by M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05), showing a significant positive correlation between the L-methionine intake and IκBα expression (r = 0.8315, P < 0.05).
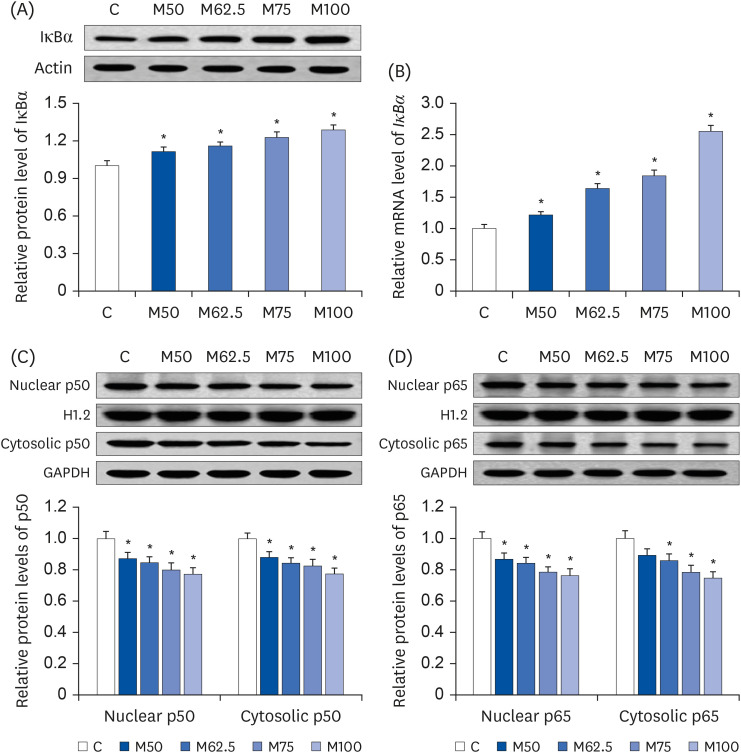 | Fig. 6Effects of L-methionine on nuclear factor-κB activation after the oral administration for 14 days. (A) Hepatic protein expressions of IκBα. (B) Hepatic mRNA levels of IκBα. (C) Nuclear and cytosolic protein contents of p50. (D) Nuclear and cytosolic protein contents of p65. The values are expressed as the means ± SEM (n = 6).IκBα, inhibitory κBα; C, control; M50, basal diet and administered L-methionine orally at 215.0 mg/kg bw; M62.5, basal diet and administered L-methionine orally at 268.8 mg/kg bw; M75, basal diet and administered L-methionine orally at 322.5 mg/kg bw; M100, basal diet and administered L-methionine orally at 430.0 mg/kg bw.
*P < 0.05, in comparison with C.
|
As a result, the dramatic decreases in the nuclear proportion of p50 (Fig. 6C) and p65 (Fig. 6D) were produced by M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05), inducing the significant inhibition of nuclear translocation of p50 and p65 after L-methionine feeding. In addition, the protein expressions of cytosolic p50 (Fig. 6C) were reduced significantly by M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05). Compared to C, the cytosolic p65 protein contents (Fig. 6D) were increased significantly in groups M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05), whereas group M50 did not produce a significant increase in the cytosolic p65 protein content (P > 0.05) compared to group C.
Significant positive correlations were observed between the hepatic HNE content and the nuclear expression of p50 (r = 0.9069, P < 0.05) and p65 (r = 0.9198, P < 0.05). In contrast, significant negative correlations were observed between the L-methionine intake and nuclear contents of p50 (r = −0.9178, P < 0.05) and p65 (r = −0.9285, P < 0.05). Overall, these results suggested that L-methionine can depress HNE-derived NF-κB activation.
Protein expression and mRNA levels of PI3K and AKT
The expression of PI3K and AKT, which could regulate the NF-kB signaling pathway, was investigated to determine if L-methionine could depress the activation of NF-κB. Compared to group C, the mRNA levels of PI3K (Fig. 7A) and AKT (Fig. 7B) were markedly reduced in groups M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05), respectively. The protein expression of PI3K and AKT was also attenuated by L-methionine intake. The dramatic depressions on PI3K protein expression (Fig. 7A) were found in M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05). As shown in Fig. 7B, the protein expression of AKT was reduced significantly by M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05), whereas no significant difference was found between M50 and C (P > 0.05). Overall, L-methionine feeding can decrease the gene and protein expression of PI3K and AKT, which are dependent on L-methionine availability.
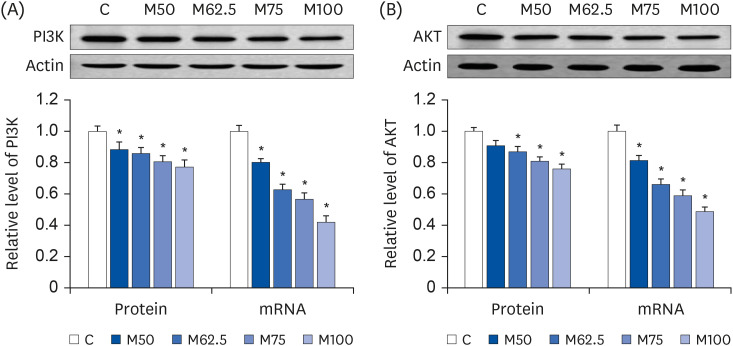 | Fig. 7Hepatic protein expressions and mRNA levels of PI3K and AKT after the oral administration of L-methionine for 14 days. (A) Hepatic protein expressions and mRNA levels of PI3K. (B) Hepatic protein expressions and mRNA levels of AKT. The values are expressed as the means ± SEM (n = 6).PI3K, phosphoinositide 3 kinase; AKT, protein kinase B; C, control; M50, basal diet and administered L-methionine orally at 215.0 mg/kg bw; M62.5, basal diet and administered L-methionine orally at 268.8 mg/kg bw; M75, basal diet and administered L-methionine orally at 322.5 mg/kg bw; M100, basal diet and administered L-methionine orally at 430.0 mg/kg bw.
*P < 0.05, compared with C.
|
In this study, negative correlations were observed between L-methionine intake and the expression of PI3K/AKT (PI3K, r = −0.8434, P < 0.05; AKT, r = −0.7983, P < 0.05). Furthermore, significant positive correlations were noted between the nuclear contents of NF-κB and expression of PI3K (p50, r = 0.9410, P < 0.05; p65, r = 0.9503, P < 0.05) and AKT (p50, r = 0.7968, P < 0.05; p65, r = 0.8079, P < 0.05).
Protein expressions and mRNA levels of IL-10 and GST
After 14 days of feeding, L-methionine regulated the protein expression and mRNA level of IL-10, which was an anti-inflammatory mediator. As shown in Fig. 8A, compared to group C, the protein expressions of IL-10 were augmented significantly in groups M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05). The mRNA levels of IL-10 were also increased markedly enhanced in groups M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05), with increases ranging from 29.23% to 126.56% (Fig. 8B). The results showed a significant positive correlation between L-methionine intake and the expression of IL-10 (r = 0.9225, P < 0.05).
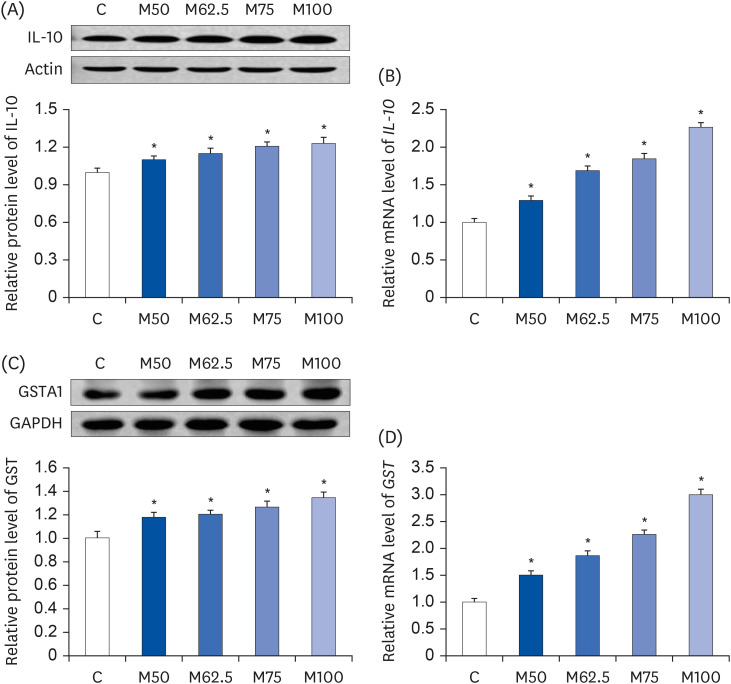 | Fig. 8Hepatic protein expression and mRNA levels of IL-10 and GST in growing rats. (A) Hepatic protein expressions of IL-10. (B) Hepatic mRNA levels of IL-10. (C) Hepatic protein expressions of GST. (D) Hepatic mRNA levels of GST. The values are expressed as the means ± SEM (n = 6).IL-10, interleukin-10; GST, glutathione S-transferase α1; C, control; M50, basal diet and administered L-methionine orally at 215.0 mg/kg bw; M62.5, basal diet and administered L-methionine orally at 268.8 mg/kg bw; M75, basal diet and administered L-methionine orally at 322.5 mg/kg bw; M100, basal diet and administered L-methionine orally at 430.0 mg/kg bw.
*P < 0.05, compared with C.
|
Similarly, the protein expressions (Fig. 8C) and mRNA levels (Fig. 8D) of GST, which could exert a HNE-scavenging action, were elevated dramatically by M50 (P < 0.05), M62.5 (P < 0.05), M75 (P < 0.05), and M100 (P < 0.05). Compared to group C, the protein expression of GST was stimulated by L-methionine in a dose-dependent manner (Fig. 8C). In addition, the mRNA levels of GST (Fig. 8D) were stimulated by L-methionine intake, with increases ranging from 50.13% (M50, P < 0.05) to 199.76% (M100, P < 0.05) in growing rats.
Go to :
DISCUSSION
The regulatory effect of L-methionine on inflammation was investigated in this study. After oral administration for 14 days, L-methionine had anti-inflammatory effects in growing rats. As major inflammatory biomarkers, the expression of COX-2, TNF-α, IL-1β, IL-6, and iNOS revealed the inflammatory process [2425]. Thus, the depression of COX-2, TNF-α, IL-1β, IL-6, and iNOS expressions was suggested to be a useful target for preventing inflammation. Hence, in this study, the anti-inflammatory action of L-methionine was reflected primarily by the down-regulated expression of these inflammatory mediators in growing rats. Moreover, L-methionine feeding could enhance the expression of IL-10, which is an important anti-inflammatory mediator [24]. Thus, the present study showed that L-methionine feeding could depress the inflammation in growing rats, which is dependent on the availability of L-methionine.
Here, the question might arise as to why L-methionine could exert an anti-inflammatory function after 14 days of feeding. To answer this question, the view that oxidative stress can initiate an inflammatory process should be considered first. HNE, which is a major end product of lipid peroxidation, can induce an inflammatory process by stimulating COX-2 expression [5]. In this study, hepatic HNE contents were significantly reduced by L-methionine feeding. More significantly, the decreased expression of the inflammatory mediators (COX-2, IL-1β, IL-6, TNF-α, and iNOS) was attributed to the decreased HNE accumulation, confirming that HNE is an important inducer of inflammation. Thus, L-methionine can inhibit HNE accumulation and prevent HNE-derived inflammation.
The view that scavenging reactive oxygen species (ROS) is a key step to inhibiting HNE-induced inflammation has merit because HNE is produced by oxidative stress [56]. L-methionine may have antioxidant capacity [2627]. In particular, in this series of studies, L-methionine could scavenge ROS [17182228], suggesting that L-methionine might decrease the formation of HNE. In this study, with oral administration, the inhibitory effect of L-methionine on hepatic HNE accumulation was observed in growing rats fed M50, M62.5, M75, or M100. Accordingly, the depression of HNE-induced inflammation resulted from attenuating the ROS-induced oxidative damage to lipids in growing rats fed with L-methionine.
This study examined the role of L-methionine in HNE-scavenging action to explain why L-methionine could exert an anti-inflammatory function after 14 days of feeding. With its strong hydrophobicity, HNE can react with DNA, proteins, and other molecules that contain nucleophilic thiol (-SH) or amino (-NH2) groups. Principally, HNE can bind to three different side-chain amino acids, i.e., cysteine (Cys), histidine (His), and lysine (Lys), to form Michael adducts via Michael addition, in which cysteine exhibits the strongest reactivity [2930]. Thus, L-methionine feeding could stimulate the transsulfuration pathway to increase the supply of cysteine [17], which can exert HNE-scavenging activity. Moreover, for a low-molecular compound, hydrogen sulfide (H2S) can also react with HNE to exert protective activity against HNE-induced toxicity. Increasing evidence suggests that the HNE formation can be inhibited markedly by the increased synthesis of endogenous H2S, which is produced by the transsulfuration pathway [2630]. Therefore, a satisfactory explanation can be further drawn from the authors’ series of studies, in which L-methionine feeding effectively stimulated the expression of cystathionine β-synthase (CBS) and cystathionine γ-lyse (CTH) to increase the supply of cysteine and promote H2S synthesis [171822]. Accordingly, the HNE-scavenging activity could be enhanced by increasing the intake of L-methionine, leading to an anti-inflammatory action in growing rats after 14 days of feeding.
The inflammatory process can be inhibited by dietary antioxidants [24]. Hence, the antioxidant effects of L-methionine must also be noted to explain the findings of this study. Previous studies reported the molecular antioxidant mechanism exerted by L-methionine [171822]. Wang et al. [17] stated that L-methionine could activate Nrf2, a key transcription factor for antioxidant mechanism, and up-regulated ARE-dependent gene expression to scavenge ROS. The results showed that L-methionine could induce the endogenous antioxidant response by increasing GSH synthesis to depress ROS-derived oxidative stress. As an endogenous antioxidant, GSH can bind HNE rapidly, showing a HNE-detoxifying effect of GSH [429303132]. Furthermore, as a major determinant of the intracellular concentration of HNE, GST also plays a role in detoxification. GST regulates the HNE metabolism via its conjugation to GSH, resulting in the depression of HNE formation during lipid peroxidation [303334]. Overall, L-methionine could decrease the HNE level, depending on enhanced GSH synthesis and stimulated GST expression exerted by L-methionine. In addition, upon the activation of Nrf2, L-methionine could stimulate the gene and protein expression of heme oxygenase 1 (HO-1) [17182223], which was another anti-inflammatory mediator [1324]. Thus, L-methionine can depress the HNE-induced inflammation, possibly due to the stronger antioxidant capacity of L-methionine.
The regulatory effect of L-methionine on the NF-κB pathway was investigated to explain how L-methionine can depress the inflammatory process after oral administration for 14 days. The major inflammatory mediators, e.g., COX-2, TNF-α, IL-1β, IL-6, and iNOS, can be up-regulated with NF-κB activation [925]. Thus, NF-κB is a master regulator for the inflammatory process and plays a crucial role in initiating inflammation [9]. Upon the activation of NF-κB, the major complexes of NF-κB, e.g., p50 and p65, can translocate to the nucleus, initiating an inflammatory process [935]. In this study, L-methionine could inhibit the activation of NF-κB by stimulating IκBα expression, which was an inhibitor of NF-κB [9]. More importantly, the significant finding was that L-methionine could inhibit the translocation of p50 and p65 to the nucleus. Consequently, the inflammatory response was inhibited by L-methionine feeding by depressing NF-κB activation.
The activation of NF-κB is closely linked to oxidative damage to lipids [2435]. As a critical marker of lipid peroxidation, HNE can regulate NF-κB activation [78]. The findings observed in this study are also consistent with this view. Therefore, the present study confirmed that L-methionine could depress NF-κB activation, which might be because of the decreased HNE accumulation by L-methionine, confirming that L-methionine could depress HNE-induced inflammation in growing rats.
NF-κB activation was also regulated by PI3K/AKT. As one of the targets of the PI3K/AKT pathway, NF-κB signaling can be activated via the up-regulation of PI3K/AKT [363738]. Wang et al. reported that the down-regulation of PI3K/AKT might be a switch to the depression of NF-κB activation involving the anti-inflammatory action exerted by rice protein [24]. Consistent with this view, this study found that the decreased nuclear contents of NF-κB were closely linked to the decreased expression of PI3K/AKT in growing rats fed L-methionine. Nevertheless, the present study clearly showed that the anti-inflammatory action of L-methionine can be explained partly by the depression of NF-κB activation via down-regulating PI3K/AKT.
In this study, L-methionine affected the anti-inflammatory action in a dose-dependent manner. With the lowest level of L-methionine, the hepatic HNE level was higher in the M50 group than in the M62.5, M75, or M100 groups. As a result, the protein expression and mRNA levels of the inflammatory mediators (COX-2, IL-1β, IL-6, TNF-α, and iNOS) were higher in the M50 group than in the M62.5, M75, or M100 groups. In contrast, the protein expression and mRNA level of the anti-inflammatory mediator (IL-10) were lower in the M50 group than in the M62.5, M75, or M100 groups. In this study, of interest was the finding that the M50 group showed significantly lower mRNA levels (P < 0.05) of the inflammatory mediators (COX-2, IL-1β, IL-6, TNF-α, iNOS, and AKT) compared to the control group. On the other hand, compared to the marked magnitude of the change in the mRNA levels (P < 0.05, the changes in the protein levels of these inflammatory mediators (COX-2, IL-1β, IL-6, TNF-α, iNOS, and AKT) were not large in the M50 group in compared to the control group (P > 0.05). This absence of a protein-mRNA correlation for investigated levels of genes and proteins of the inflammatory mediators in this study might be due to the relationship between the protein and mRNA, which is not strictly linear. Moreover, under the present experimental condition, M100 exhibited the strongest anti-inflammatory activity, whereas the weakest anti-inflammatory effect was induced by M50. The influence of L-methionine on the antioxidative capacity should be considered when explaining this phenomenon. These findings highlighted the critical role of L-methionine in the ROS-scavenging activity and activating the endogenous antioxidant system via the Nrf2-ARE pathway to inhibit lipid oxidation, which is dependent on the availability of L-methionine. A link between the higher composition of L-methionine and stronger ROS-derived HNE scavenging activity is conceivable. This is supported by the lower protein expression and mRNA level of GST, which could exert HNE-scavenging activity, in the M50 group than in the M62.5, M75, or M100 groups. The present study confirmed that higher levels of L-methionine might possess a stronger anti-inflammatory function.
In summary, L-methionine can inhibit hepatic HNE accumulation and depress HNE-derived inflammation in growing rats, which is dependent on the availability of L-methionine. With the oral administration of L-methionine, the hepatic HNE-scavenging activity was attributed to the stimulation of GST by L-methionine. With the decreased HNE level, the anti-inflammatory action is reflected by the suppressed expression of the inflammatory biomarkers (COX-2, TNF-α, IL-1β, IL-6, and iNOS) and the up-regulation of the anti-inflammatory mediator (IL-10). Of significance is the finding that the molecular mechanism underlying the anti-inflammatory capacity of L-methionine is to inhibit NF-κB activation by down-regulating PI3K/AKT (Fig. 9). Nevertheless, more detailed investigations will be needed to explore the precise mechanisms exerted by L-methionine to prevent inflammation.
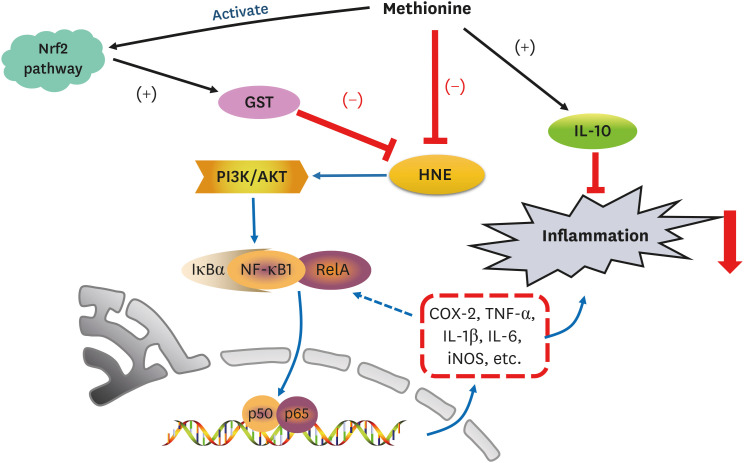 | Fig. 9L-Methionine inhibits HNE accumulation and depresses inflammation by inhibiting NF-κB activation and up-regulating GST.Nrf2, nuclear factor erythroid 2-related factor 2; GST, glutathione S-transferase; PI3K, phosphoinositide 3 kinase; AKT, protein kinase B; HNE, 4-hydroxy-2-nonenal; IκBα, inhibitory κBα; NF-κB, nuclear factor-κB; COX-2, cyclooxygenase-2; TNF-α, tumor necrotic factor alpha; IL, interleukin; iNOS, inducible nitric oxide synthase.
|
Go to :
 XML Download
XML Download