

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Obesity is the central component of the pathogenesis of obesity-related diseases, including metabolic syndrome, insulin resistance, type 2 diabetes, non-alcoholic fatty liver disease (NAFLD), dyslipidemia, hypertension, atherosclerosis, and several forms of cancer. The increased susceptibility to these diseases in obese individuals has become a significant public health issue worldwide [1]. The mechanisms linking obesity to these metabolic and vascular disorders have been proposed, but they are not completely understood. Recent evidence highlighted the action of adipose tissue in triggering a systemic inflammatory response and the role of the altered adipokines levels in the pathogenesis of obesity-associated complications [2].
Obesity is characterized by an enlarged adipose tissue mass. Adipose tissue is involved in the storage and mobilization of lipids. In addition, it is also an endocrine organ that releases numerous adipokines, affecting the metabolism and vascular function [3]. Adipokines play critical roles in different biological processes, including inflammation, glucose and lipid metabolism, blood pressure, hemostasis, endothelial function, and feeding behavior, thereby regulating the metabolism and function of many organs, such as muscle, liver, vasculature, and brain [4]. Therefore, changes in adipokines secretion promote susceptibility to inflammatory, metabolic, and cardiovascular obesity-related disorders, but the pathophysiologic mechanisms are unclear [56].
Leptin and adiponectin are 2 major adipokines related to the risk of metabolic syndrome, type 2 diabetes, and cardiovascular disease (CVD), playing contrasting roles in the pathogenesis of many obesity-related diseases [7]. Leptin enhances lipotoxicity and insulin resistance and the production of pro-inflammatory cytokines [8]. In contrast, adiponectin reduces the pro-inflammatory cytokines and oxidative stress, protecting against obesity-related disorders, including type 2 diabetes, metabolic syndrome, and CVD [9]. Hence, the development of obesity-related disorders has been attributed to an imbalance in adipokines. Recent accumulating data have suggested the leptin-to-adiponectin (L/A) ratio or adiponectin-to-leptin (A/L) ratio as a useful marker to determine the insulin resistance in patients with and without diabetes [1011].
The incidence of NAFLD has also increased in parallel with the prevalence of obesity [12]. Excessive lipids usually accumulate in the liver when the influx of lipids or synthesis of hepatic lipids exceeds their export or oxidation. In an obese state, the free fatty acids from adipose tissue directly enter the liver via the portal circulation, which promotes the accumulation of lipids in the liver [1314]. Adiponectin is an adipokine with potential beneficial effects on NAFLD, whereas others, such as leptin, resistin, and interleukin (IL)-6, have an adverse effect on NAFLD. Adiponectin reduces hepatic lipogenesis by suppressing the lipogenic transcription factor, sterol regulatory element-binding protein1-c (SREBP1-c), and enhances glucose utilization and fatty acid oxidation in the liver through activation of adenosine monophosphate-activated protein kinase (AMPK). In addition, adiponectin exerts anti-inflammatory properties that might prevent the progression of hepatic steatosis to fibrosis [1516]. Leptin is one of the potent regulators of NAFLD. A positive correlation of leptin with hepatic steatosis or serum ALT has been reported. Leptin has also been reported to play a role in liver fibrosis leading to cirrhosis by activating hepatic stellate cells [17].
Many bioactive food components have been assessed for their ability to prevent a wide array of chronic disorders. Natural bioactive compounds found in fruits and vegetables have attracted increasing attraction owing to their antioxidant activities and health benefits in obesity and obesity-associated metabolic abnormalities [18]. Among the natural products, mulberry, which belongs to the genus Morus of the Moraceae family, has been cultivated in many countries, including China, Korea, Japan, and Taiwan [19]. Although there are limited studies on the beneficial health effects of mulberry fruits, recent studies have reported that mulberry fruit has protective properties against dyslipidemia, diabetes, obesity, and liver damage [18]. Mulberry extracts have potential effects, including inhibiting hepatic triglyceride (TG) accumulation, anti-obesity, and hypolipidemic effects, and preventing atherosclerosis in a non-alcohol-induced liver disease model [20].
This study examined the protective effects of mulberry (Morus alba L.) fruit ethanol extract (MBEE) on metabolic disturbances using high-fat diet-fed rats. The relationship between lipid metabolism disturbance and adipokine balance was examined in high-fat diet-induced obese rats. The effects of an MBEE treatment on dyslipidemia, hepatic fat accumulation, and adipokine imbalance were also evaluated with respect to the leptin/adiponectin and plasminogen activator inhibitor-1 (PAI-1)/adiponectin ratios.
Go to : 
MATERIALS AND METHODS
Animal care and experimental design
Male Sprague-Dawley rats at 6 wk of age were acclimatized for 1 wk with normal rat chow pellet at room temperature. After the acclimatization period, the rats were assigned randomly to one of 4 groups of 6 rats each. The normal control group (CON) received a normal control diet (Table 1). Three groups were given a high-fat diet, of which the HFD group was given no MBEE, and the LMB and HMB groups received low-dose (150 mg/kg/day) and high-dose of MBEE (300 mg/kg/day), respectively. The rats in the LMB and HMB groups were administered MBEE through an oral gavage for 6 wk, and those in the CON and HFD groups were given an equal volume of water. The HFD, LMB, and HMB diets included 2% soybean oil to ensure sufficient essential fatty acid. MBEE was prepared from dried mulberry (Morus alba L.), which was harvested in South Chungcheong Province. Briefly, the cleaned mulberry fruits were finely powdered and extracted with 20 volumes of 80% ethanol. After incubation for 2 h in a shaking water bath set to 40°C, the extract was filtered, and the solvent was removed using a vacuum rotary evaporator. Subsequently, the filtrate was collected and lyophilized to obtain MBEE and stored at −20°C. The animals were given free access to tap water and feed. The feed consumption and weight gain were recorded weekly. This experimental protocol was approved by the Dongeui University Laboratory Animal Care Committee (R2018-010).
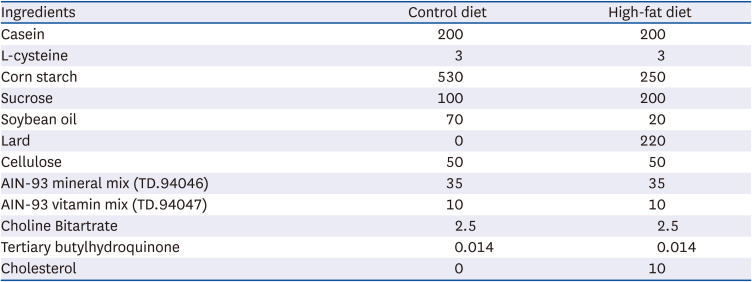
Table 1
Composition of the experimental diets (g/kg)
Rats were fed either a control diet, high- fat diet with no mulberry fruit ethanol extract (MBEE), high-fat diet with MBEE of 150 mg/kg/day, or high-fat diet with MBEE of 300 mg/kg/day.
![]()
Preparation of samples
After 6 wk of feeding, all animals were fasted overnight and euthanized. Blood from the abdominal aorta was collected into a lithium-heparinized tube and centrifuged at 3,000 rpm for 15 min at 4°C. The plasma was separated and stored at −70°C for later analysis. The epididymal adipose tissue was harvested and rinsed with ice-cold saline. The liver was perfused with ice-cold saline to remove the residual blood, excised, and rinsed with ice-cold 0.1 M phosphate-buffered saline.
Determination of plasma lipids
The TG, total cholesterol (TC), and high-density lipoprotein cholesterol (HDL-C) concentrations were quantified in plasma by spectrophotometry using a commercially available enzymatic colorimetric kit (Asan Pharmaceuticals, Seoul, Korea). The TG levels were determined using a lipase-glycerol phosphate assay. The cholesterol measurements were based on the cholesterol oxidase method. HDL-C was determined after separating the HDL fraction using a heparin-manganese precipitation procedure. The low-density lipoprotein cholesterol (LDL-C) concentration was calculated using the Friedewald formula [21].
Determination of adipokines
Adipokines in plasma were analyzed by an enzyme-linked immunosorbent assay using commercially available kits. Leptin, PAI-1, and adiponectin were measured on a Sunrise microplate reader (Tecan, Grödig, Austria), using an ELISA kit (Enzo Life Sciences, Farmingdale, NY, USA), Simple Step ELISA™ kit (Abcam, Cambridge, MA, USA), and Quankine ELISA kit (R&D Systems, Minneapolis, MN, USA), respectively. All procedures were performed according to the manufacturer’s instructions.
Determination of hepatic lipids
The hepatic lipids were extracted using a modified method reported by Folch et al. [22]. Briefly, the liver tissues were homogenized with chloroform/methanol (v/v, 1:2) using a Potter-Elvehjem tissue grinder. Chloroform/distilled water (v/v, 1:1) was then added and mixed thoroughly. The mixture was then centrifuged at 3,000 rpm for 15 min at 4°C to separate into 2 phases. The lower chloroform layer was collected and evaporated under N2 gas. The liver lipid extract was dissolved in chloroform/methanol and stored at −20°C. The hepatic TG and cholesterol contents in the lipid extract were quantified using a colorimetric assay with a commercial kit (Asan Pharmaceuticals) according to the manufacturer’s instructions.
Determination of activities of alanine aminotransferase (ALT) and aspartate aminotransferase (AST)
The plasma ALT and AST activities were determined using a colorimetric assay with commercially available kits (Asan Pharmaceuticals). The absorbance was read at 505 nm according to the manufacturer’s protocols.
Statistical analysis
Statistical analyses were conducted using SAS software (version 9.3; SAS Inc., Cary, NC, USA). The values were presented as the means and SEM. One-way analysis of variance (ANOVA) followed by a Duncan’s multiple range test was used to determine the differences between the means of the experimental groups. A P-value < 0.05 was considered significant.
Go to :
RESULTS
Changes in body weight and fat mass
All 4 groups showed a similar initial body weight and feed intake, as listed in Table 2. The average weight gain and final body weight were higher in the HFD group than in the CON group, but were similar in the HFD, LMB, and HMB groups. Epididymal fat mass was significantly higher in the HFD group than in the CON group (P < 0.05). The significance remained, even when represented as mass per 100 g body weight. On the other hand, the fat mass and fat index were similar in the HFD, LMB, and HMB groups, but there was a 10% reduction of each in the HMB group compared to HFD.
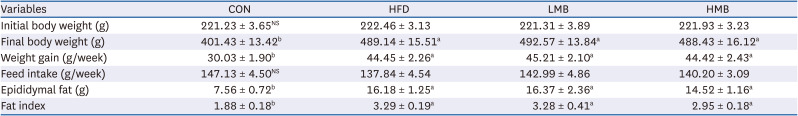
Table 2
Effects of the mulberry ethanol extract on the body weight and epididymal fat mass
Data are presented as mean ± SEM (n = 6). Fat index is fat mass per 100 g body weight.
CON, control diet; HFD, high-fat diet with no mulberry fruit ethanol extract (MBEE); LMB, high-fat diet with MBEE of 150 mg/kg/day; HMB, high-fat diet with MBEE of 300 mg/kg/day.
a,bValues within raw not sharing common superscript letters are significantly different using Duncan’s multiple range test (P < 0.05).
![]()
Changes in plasma markers of dyslipidemia
The cholesterol and LDL-C levels and TC/HDL-C ratio were significantly higher in the HFD group than in the CON group (Table 3). On the other hand, the HDL-C was lower in the HFD group than in the CON group (P < 0.05). Significant decreases in cholesterol, LDL-C, and TC/HDL-C were observed in MBEE-treated groups, particularly in the LMB group, indicating mulberry extracts had plasma lipid-lowering effects. There was no significant difference in HDL-C with MBEE administration, even though HDL-C was increased in the LMB group. No significant difference in the plasma TG level was observed among the groups. On the other hand, there was a higher tendency of TG in the HFD group, whereas the HMB group showed lower levels than the HFD group. The plasma glucose levels were significantly higher in the HFD group than in the CON group. The plasma glucose level of the HFD group was not attenuated by the MBEE treatment, but the LMB and HMB groups tended to be lower than the HFD group.
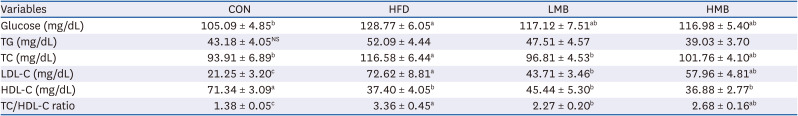
Table 3
Effects of mulberry ethanol extract on the plasma glucose and lipid metabolic parameters
Data are presented as mean ± SEM (n = 6).
CON, control diet; HFD, high-fat diet with no mulberry fruit ethanol extract (MBEE); LMB, high-fat diet with MBEE of 150 mg/kg/day; HMB, high-fat diet with MBEE of 300 mg/kg/day; TG, triglyceride; TC, total cholesterol; LDL-C, low-density lipoprotein cholesterol; HDL-C, high-density lipoprotein cholesterol.
a-cValues within raw not sharing common superscript letters are significantly different using Duncan’s multiple range test (P < 0.05).
![]()
Changes in plasma markers of inflammation
Adiponectin level was not different among the 4 groups, as shown in Fig. 1. The leptin level was significantly greater in the CON and HFD groups but lower in the LMB and HMB groups (P < 0.05). The HFD group showed a significant increase in the PAI-1 level compared to the CON group, whereas the PAI-1 levels tended to be lower in the LMB and HMB groups. The leptin/adiponectin and PAI-1/adiponectin ratios in the HFD group were similar to those in the CON group. The MBEE treatment reduced both the leptin/adiponectin and PAI-1/adiponectin ratios, showing a significant decrease, especially in the HMB group (P < 0.05). Overall, the MBEE treatment alters the adipokine balance to the anti-inflammatory state by reducing leptin and the leptin/adiponectin and PAI-1/adiponectin ratios.
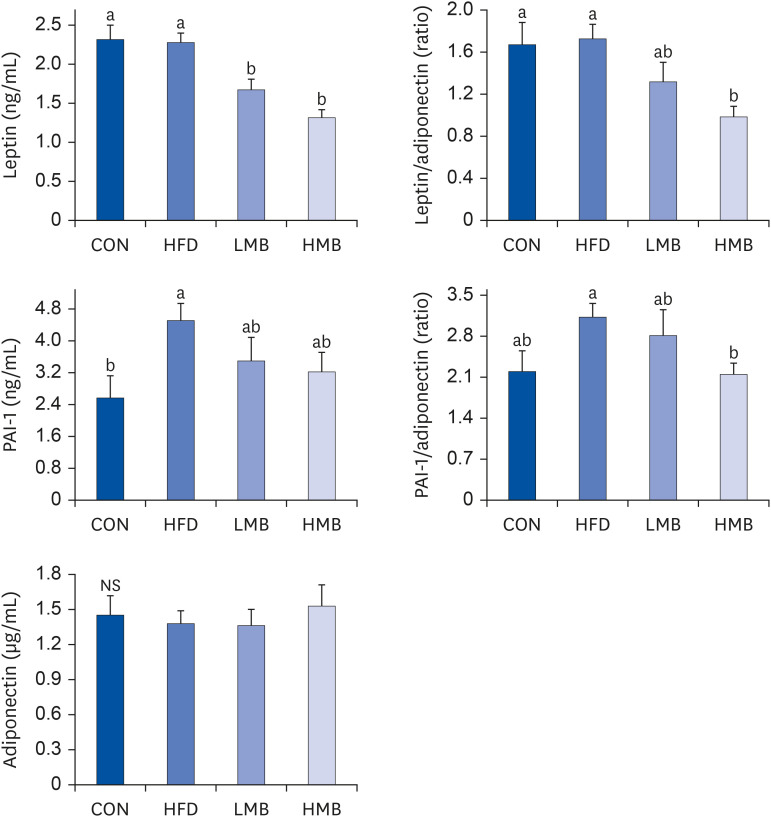 | Fig. 1Effects of mulberry ethanol extract on the plasma leptin, PAI-1, and adiponectin levels.Data are presented as mean ± SEM (n = 6).
CON, control diet; HFD, high-fat diet with no mulberry fruit ethanol extract (MBEE); LMB, high-fat diet with MBEE of 150 mg/kg/day; HMB, high-fat diet with MBEE of 300 mg/kg/day; PAI-1, plasminogen activator inhibitor-1.
a,bData not sharing common superscript letters are significantly different using Duncan’s multiple range test (P < 0.05).
|
Changes in markers of hepatic lipid accumulation
Fig. 2 shows fat accumulation in the liver and the ALT and AST activities. The hepatic TG level was higher in the HFD group than in the CON group and significantly lower in the LMB and HMB groups. The hepatic cholesterol was higher in the HFD group than in the CON group, but was similar in the HFD, LMB, and HMB groups. The HFD group showed higher ALT and AST activities than the CON group (P < 0.05). On the other hand, the AST activity was markedly lower in the LMB and HMB groups, and the ALT activity was lower in the HMB. Moreover, the HMB group showed lower ALT and AST activities. These results suggest that HFD caused hepatic lipid accumulation accompanied by an increase in the ALT and AST activities, which could be improved by MBEE.
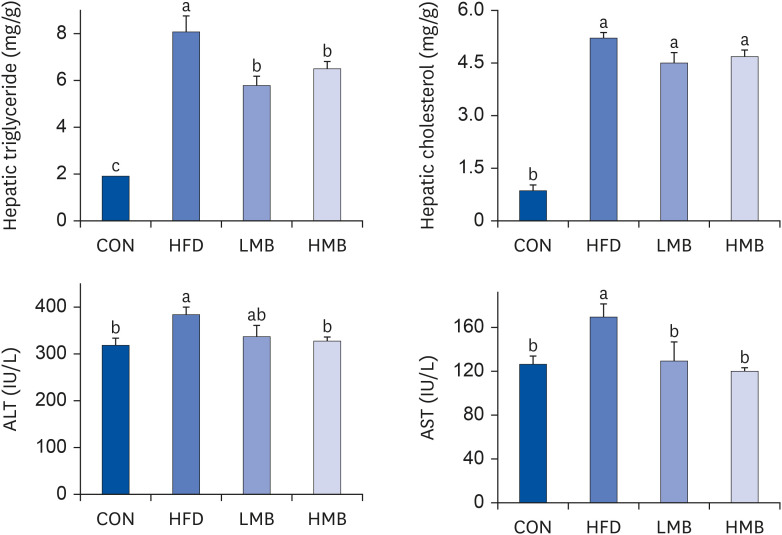 | Fig. 2Effects of mulberry ethanol extract on the accumulation of hepatic triglyceride and cholesterol and activities of ALT and AST.Data are presented as mean ± SEM (n = 6).
CON, control diet; HFD, high-fat diet with no mulberry fruit ethanol extract (MBEE); LMB, high-fat diet with MBEE of 150 mg/kg/day; HMB, high-fat diet with MBEE of 300 mg/kg/day; ALT, alanine aminotransferase; AST, aspartate aminotransferase.
a,bData not sharing common superscript letters are significantly different using Duncan’s multiple range test (P < 0.05).
|
Go to :
DISCUSSION
Obesity, particularly visceral obesity, is related to a metabolic and cardiovascular disturbances, including insulin resistance, dyslipidemia, and NAFLD. The mechanism whereby the increased body fat has an adverse effect is poorly understood. An expansion of adipose tissue induces the filtration of macrophages and an imbalance of pro- and anti-inflammatory adipokines secreted by adipose tissue, thereby affecting inflammation, insulin resistance, and metabolic dysregulation of lipid. Adipose tissue can affect the metabolism and function of many tissues, such as the liver, skeletal muscle, and heart, by releasing free fatty acids and producing several adipokines [23]. The present study investigated the protective effects of MBEE supplementation against high-fat-induced dyslipidemia, hepatic steatosis, and adipokine imbalance between pro-inflammatory leptin and PAI-1 and anti-inflammatory adiponectin.
In most experimental obesity studies, the degree of obesity has been assessed by comparing the body weight or body fat of the high-fat-fed animals with the control group given a low-fat diet [24]. After feeding the experimental diets for 6 wk in this study, the body weight of the HFD group increased. Moreover, the HFD rats gained significantly higher visceral adipose tissue measured by epididymal fat mass than CON rats, resulting in an obese state. On the other hand, the MBEE treatment in LMB and HMB groups did not markedly reduce the high-fat diet-induced weight gain and adipose tissue increase. One study reported that the rats supplemented with 5% or 10% mulberry fruit powder for 4 wk showed no significant decrease in high-fat-induced body weight gain [25]. By contrast, it was reported that administration of mulberry ethanol extract for 14 wk or purified anthocyanins for 12 wk resulted in a significant decrease in weight gain and adipose hypertrophy in high-fat-fed mice, suggesting mulberry has an anti-obesity effect [2627]. These studies suggest that the anti-obesity effects of mulberry fruits might differ according to the treatment duration and supplementation as whole food extract or isolated compounds and their preparation methods, resulting in different biological compounds.
Obesity is linked to an increased prevalence of dyslipidemia. According to Woods et al. [28], high-fat-fed rats weighed more and developed more adipose tissue than the control, having insulin resistance and hyperlipidemia typically associated with obesity. One of the major mechanisms behind dyslipidemia is the increased flux of free fatty acid (FFA) from adipose tissue to the liver through the portal vein when visceral TG stores are increased. Increased lipolytic activity in obese subjects triggers the release of FFA by adipocytes. The uncontrolled release of FFA from adipose tissue, particularly visceral adipose tissue, causes the increased flux of FFA to the liver and stimulates the synthesis and secretion of very-low-density lipoprotein (VLDL), as well as the suppression of lipoprotein lipase in adipose tissue and skeletal muscle, leading to hypertriglyceridemia [2930]. Hypertriglyceridemia promotes the formation of small-dense LDL particles and a decrease in HDL by cholesteryl ester transport protein and subsequent hepatic lipase [31].
HFD rats caused a disturbance of the lipid profile in the present study. The cholesterol and LDL-C plasma levels and the TC/HDL-C ratio were significantly higher in the HFD group than in the CON group. The HDL-C level was significantly lower in the HFD group than in the CON group. Significant decreases in the cholesterol and LDL-C levels and TC/HDL-C ratio were observed in the MBEE-treated groups compared to the HFD group, particularly in the LMB group. On the other hand, the plasma triglyceride levels were lower in the HMB group, but the difference was not significant. Based on these results, the dyslipidemic picture induced by a high-fat diet in the present study was alleviated by the MBEE treatment. Nevertheless, the HMB group appeared to show fewer effects on LDL-C and TC/HDL-C than a low-dose in the LMB group. Despite the lower efficacy of HMB in lowering plasma lipids, a high-dose of MBEE in HMB helped reduce the plasma lipids when comparing each variable in the HFD and HMB groups.
Adipose tissue is considered an endocrine organ releasing many bioactive mediators termed adipokines, which have diverse actions on lipid and glucose metabolism, hemostasis, blood pressure, inflammation, and endothelial function. The well-characterized adipokines are adiponectin, leptin, IL-6, tumor necrosis factor-α, and PAI-1, which could be divided into those having a positive or negative impact [32]. Leptin has pro-inflammatory and atherogenic activities, whereas adiponectin has insulin-sensitizing, anti-inflammatory, and anti-atherogenic properties, as well as anti-steatotic activity in non-adipose tissues [33]. PAI-1 is involved in the inflammatory processes and inhibition of endogenous fibrinolysis. Thus, an increased PAI-1 level is responsible for thrombosis and endothelial dysfunction. PAI-1 is increased in obese subjects, which is related to dyslipidemia, hyperinsulinemia, and high blood pressure. Therefore, when aberrant production of adipokines is persistent, the body is likely to face an inflammatory and prothrombotic milieu, which can contribute to the development of obesity-linked metabolic diseases [5634]. A single adipokine may not cause metabolic syndrome, but the interaction between the pro- and anti-inflammatory adipokines induces systemic abnormalities while a balance between the pro- and anti-inflammatory adipokines is disrupted as perivascular and visceral obesity increases. The balance between the anti- and pro-inflammatory adipokines is vital for preventing metabolic syndrome, particularly in central obese subjects [35]. The correlations of the adiponectin/leptin ratio with variations in systolic blood press, insulin sensitivity, TC, and LDL have been observed in obese metabolic syndrome patients [36]. According to a study on the relevance of the L/A ratio and lipid profiles in NAFLD, the L/A ratio correlated directly with the triglyceride level and TC/HDL-C ratio and inversely with HDL-C [37]. The L/A ratio was proposed as a parameter to determine the risk of obesity- and metabolic syndrome-related CVD. Regarding the capacity to discriminate the risk of metabolic syndrome, the L/A ratio was better than adiponectin and leptin alone among healthy individuals, suggesting that the ratio could be a significant predictor of the risk of metabolic syndrome [38].
In the present study, the adiponectin level was similar in the 4 groups. Leptin was not increased in the HFD group, but there was a significant increase in PAI-1 in the HFD group. Leptin was significantly lower in both LMB and HMB groups with MBEE treatment, and the PAI-1 level, albeit not significant, tended to be lower. Subsequently, leptin/adiponectin and PAI-1/adiponectin ratios also decreased with MBEE. As a result, the HFD group appeared to show an imbalance in the production of anti-/pro-inflammatory adipokines based on the higher PAI-1 level and the higher tendency of the PAI-1/adiponectin ratio. The MBEE treatment, especially in the HMB group, altered the balance of adipokines to an anti-inflammatory state, as shown by the decrease in leptin level and the leptin/adiponectin and PAI-1/adiponectin ratios. Hence, the dyslipidemia-improving effect appears to be associated with regulating the adipokine balance.
NAFLD is one of the most common complications of obesity and is generally regarded as a hepatic manifestation of metabolic syndrome. Hepatic steatosis, the hallmark of NAFLD, is the progression of excessive triglyceride accumulation by the metabolic disturbances between lipid uptake, synthesis, catabolism, and secretion [39]. The HFD group showed significantly higher hepatic TG and cholesterol levels than the CON group and higher plasma ALT and AST activities, which are serum indicators of liver injury and function. The hepatic TG was higher in the HFD groups but lower in the LMB and HMB groups. In addition, the ALT and AST activities were significantly lower in the MBEE-treated groups, notably in the HMB group. Overall, the HFD induced hepatic lipid accumulation, accompanied by an elevation of ALT and AST activities. The MBEE treatment improved the hepatic steatosis and lowered the ALT and AST activities.
A previous study reported that body fat mass and liver fat were positively correlated with the amount of VLDL1 apolipoprotein B and VLDL1 TG production rate [40] and that hypertriglyceridemia in metabolic syndrome is related to the over-secretion of TG-rich VLDL particles [30]. On the other hand, a positive correlation was not always observed between the plasma TG and hepatic steatosis [41]. Some studies suggested that the effects of mulberry on the relationship between the plasma TG and hepatic TG were inconsistent. The MBEE in the present study induced a significant decrease in hepatic TG but not at the plasma TG level. In agreement with the present findings, the administration of mulberry ethanol extract (100 mg/kg/day) to high-fat-fed mice revealed a decrease in hepatic TG and no change in plasma TG [26]. By contrast, supplementation of high-fat-fed rats with freeze-dried mulberry powder reduced the serum and hepatic TG levels [25]. Furthermore, mice fed black elderberry extract rich in anthocyanins showed a significant reduction in serum TG level but no improvement in the hepatic TG [42].
The pathogenesis of NAFLD involves dysfunction in adipose tissue, insulin resistance, and inflammation, but the mechanism is not yet completely understood [39]. The roles of leptin and adiponectin have attracted increasing attention regarding the NAFLD pathophysiology, diagnosis, and even treatment. A recent study examined the role of circulating adipokines as a potential non-invasive biomarker for NAFLD diagnosis and stratification. Higher circulating leptin was related to a higher serum lipid profile and a higher hepatic steatosis grade when evaluated using histology and imaging.
Adiponectin was negatively correlated with the liver fat contents, suggesting a protective role of adiponectin by causing lipid disposal from the liver into circulation [43]. Furthermore, adiponectin inhibits hepatic lipid deposition by stimulating fatty acid oxidation via peroxisome proliferator-activated receptor-α (PPAR-α) [44]. In addition, the adiponectin level is inversely correlated with the serum indicators of liver injury, ALT and AST, suggesting a role in preventing liver damage. The serum leptin level was higher in patients with severe liver steatosis, whereas adiponectin decreased with the severity of steatosis [43].
Considering the relationship between leptin and adiponectin and liver steatosis, these hepatoprotective results by MBEE appeared to be related to the lower leptin level, and the lower leptin/adiponectin and PAI-1/adiponectin ratios. A recent study reported that ALT and AST had a positive correlation with hepatic triglyceride accumulation [45]. The high-fat diet-induced increases in hepatic TG and cholesterol levels and serum ALT and AST levels were reduced significantly by the mulberry ethanol extract [26]. The anti-hepatosteatosis effects of mulberry were attributed to its ability to inhibit fatty acid and triglyceride synthesis and promote fatty acid oxidation [2646]. In ethanol-induced liver injury mice, a mulberry water extract (MWE) reduced both hepatic triglyceride and cholesterol and reduced the levels of liver injury biomarkers, ALT and AST. The hepatoprotective effect of MWE might be mediated by multiple pathways. A previous study suggested that MWE treatment inhibits lipogenesis and hepatic lipid accumulation and increases the hepatic expression of carnitine palmitoyltransferase-1 (CPT-1) and microsomal triglyceride transfer protein for fatty acid oxidation or transport by activating AMPK and PPAR-α. In addition, MWE suppresses lipid peroxidation and inflammation by enhancing the antioxidative activity and decreasing pro-inflammatory mediators [20]. Based on previous reports and the present results, mulberry might be a potential candidate for preventing NAFLD by reducing hepatic lipid deposition and inhibiting hepatic steatosis.
Polyphenols provide health benefits. Mulberry fruit contains considerable amounts of biologically active ingredients, including phenolics, flavonoids, anthocyanins, and many other antioxidants, which might have some beneficial activities, such as anti-obesity, anti-cholesterol, and hepatoprotective effects [18]. Given the complexity and breadth of polyphenols as a chemical family, the physicochemical characteristics of polyphenols are likely to contribute to the disparity in results between studies. As the main components of mulberry fruit, anthocyanins could play a significant role in promoting health. On the other hand, it was suggested that the health outcomes observed might not be due to the action of a single bioactive compound in mulberry fruit but to the cooperative action of diverse compounds, including anthocyanins, flavonoids, and phenolics [47]. Furthermore, freeze-dried mulberry fruit powder has high dietary fiber contents, antioxidant vitamins, and linoleic acid, which might play a role in the hypolipidemic effects [25]. Therefore, further studies will be needed to determine the main bioactive components and mechanisms that may be responsible for the effects of mulberry on metabolic disturbances. In addition, it is important to consider the adverse effects of MBEE. Information on the safe dose of mulberry regarding the possible adverse effects on animal health is lacking. No adverse effects on the changes in liver or renal functions were reported in rabbits fed high cholesterol diet supplemented with a 0.5% and 1.0% water extract of mulberry fruit [48]. The 2 levels of MBEE used in the present study also appear harmless, based on no significant decreases in growth and feed intake and no significant liver damage. Nevertheless, the safe dose of mulberry fruit and its treatment duration remains to be determined owing to limited data regarding adverse effects.
This study showed that a high-fat diet induced dyslipidemia with increases in plasma cholesterol and LDL-C level and TC/HDL-C ratio. Increases in hepatic triglyceride and cholesterol accumulation and higher activities of ALT and AST were also observed, suggesting the risk of developing hepatic steatosis. The administration of MBEE improved the plasma cholesterol and LDL-C levels and TC/HDL-C ratio. In addition, it also attenuated the hepatic triglyceride level and ALT and AST activities. Moreover, the adipokine equilibrium was altered towards a more anti-inflammatory state, as revealed by the lower leptin level and leptin/adiponectin and PAI-1/adiponectin ratios. Therefore, mulberry fruit ethanol extract could have anti-lipidemic and hepatoprotective effects by regulating the balance between pro-inflammatory and anti-inflammatory adipokines.
Go to :
 XML Download
XML Download