

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
P. grandiflorum (PG) is a perennial herb of the Campanulaceae family found in East Asian countries. It comprises a long list of edible and medicinal ingredients. The roots of PG are known as Kilkyung or Doraji in Korea, Kikiyo in Japan, Jiigeng in China, and balloon flower in English [1]. Compared to other parts of the PG plant, the roots contain the highest amounts of nutrients. Their good taste and aroma make them a desirable ingredient in various foods such as salads, kimchi, and tea [2]. PG is not only effective in relieving symptoms of respiratory diseases such as bronchitis and asthma but also has anti-inflammatory, antioxidant, anti-tumor, anti-diabetic, hepatoprotective and other activities in various diseases [3]. The main components of PG are amino acids, fibers, vitamins, and other essential trace elements, which are beneficial to human health [4]. PG also contains bioactive compounds such as flavonoids, phenolic acids, saponins, and sterols (beta-sitosterol and spinasterol). The main bioactive components of PG are saponins such as platycodin A, platycodin C, and platycodin D. In particular, platycodin D is not only present in large amounts but also exerts excellent physiological activity. Hence, the pharmacological activity of PG is considered to be mainly due to platycodin D [2].
Although PG has been applied as an effective treatment for various diseases, its use as a food remains much higher. In Japan, 150 tons of PG is consumed annually. The annual PG production in China is 1,000 tons, of which half is exported. Imports of fresh PG from many countries are rapidly increasing. When used for medicine, the economic value of PG increases by about 2.5 times compared to its use as a vegetable [2]. Research is therefore focusing on improving the bioactivity of PG. Recently, there has been a high interest in health functional foods derived from various natural substances that are excellent in function and exert health benefits without side effects. Therefore, research to maximize the functionality of PG is worthwhile.
In order to develop a therapeutically efficacious product, it is necessary to increase the extraction ratio of the active ingredients from PG [5]. One way is to ferment PG with fungi to form a fermented PG. Although PG itself contains large amounts of various physiologically active ingredients, it is reported that fermentation results in an increase in the active ingredients [6]. Generally, fermentation is known to improve biological activities [7]. In addition, microstructures of low-molecular biologically active substances can be decomposed into bio-absorbable structures of beneficial enzymes using microbial fermentation [8]. Reports indicate that incubation of the PG extract (PGE) with Saccharomyces cerevisiae increases the relative platycodin D content, and a fermented PGE exhibits higher anti-inflammatory efficacy than PGE [9].
The current study was conducted to verify whether a hydrolyzed and fermented PGE (HFPGE), which is expected to have a higher active ingredient content by adding a hydrolysis process to PG in addition to the conventional fermentation technique, has an immune-stimulatory effect. We analyzed the components of HFPGE by high-performance liquid chromatography (HPLC), and examined the effect of HFPGE on the proliferation, phagocytosis, and cytokine secretion in RAW 264.7 cells. In addition, we also investigated the molecular mechanism by which HFPGE affects the immune system.
MATERIALS AND METHODS
Materials
The reagents and materials used in this study were purchased from the indicated suppliers: Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum (FBS), and penicillin/streptomycin from Welgene Inc. (Gyeongsan, Korea); 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), Folin-Ciocalteu’s phenol reagent, ginsenoside-Re, platycodin D, quercetin, and tannic acid from Sigma-Aldrich Co. (St. Louis, MO, USA); Trizol from Invitrogen Life Technologies (Carlsbad, CA, USA); HyperScriptTM RT master mix kit from GeneAll Biotechnology (Seoul, Korea); Rotor-GeneTM SYBR Green kit from Qiagen (Valencia, CA, USA); proteome profilerTM mouse cytokine array kit and enzyme-linked immunosorbent assay (ELISA) kits for interleukin (IL)-1β, IL-6, tumor necrosis factor (TNF)-α, C-X-C motif chemokine ligand (CXCL)10, granulocyte-colony-stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), and monocyte chemoattractant protein (MCP)-1 from R&D Systems (Minneapolis, MN, USA); antibodies against nuclear factor (NF)-κB p65, p-p65, extracellular signal-regulated kinase (ERK), p-ERK, c-Jun N-terminal kinase (JNK), p-JNK, and β-actin from Cell Signaling Technology (Beverly, MA, USA).
Preparation of HFPGE
The HFPGE used in the study was produced by World Food Services Co., Ltd. (Gangneung, Korea). Dried roots of PG cultivated for 3 years in Korea were used to produce the HFPGE. Briefly, dried roots of PG were rinsed, dried, and pulverized. The powder of PG was homogenized with water in a 1:9 ratio of powder:water and heated at 100°C for 30 min. The slurry was added to 3% (w/w) Bromelain (Novozymes, Copenhagen, Denmark) and incubated at 50°C for 3 h, after which it was added to 3% (w/w) Celluclast (Novozymes) and incubated at 50°C for 3 h. The slurry hydrolyzed by Bromelain and Celluclast was subsequently heated at 100°C for 30 min to inactivate these enzymes, followed by cooling. To ferment the hydrolyzed slurry, Lactobacillus casei cultured in tangerine shells was inoculated at a concentration of 1 mL per kg of the hydrolyzed slurry. This mixture was incubated at 37°C for 24 h, followed by heating at 100°C for 30 min. The final slurry was filtered through Whatman filter paper, and the resultant filtrate was freeze-dried using a vacuum freeze-dryer (Operon, Kimpo, Korea). This final powder was used as the HFPGE. In addition, PGE without hydrolysis and fermentation was produced as a control extract of HFPGE. Briefly, powdered dried roots of PG were homogenized with water in a 1:9 ratio of powder:water, heated at 100°C for 30 min, and filtered. This filtrate was freeze-dried and used as PGE.
Determination of total phenolic and flavonoid content
The total phenolic content of the extracts was measured by the Folin and Ciocalteu colorimetric method [10], with slight modifications. A total of 0.5 mL extract (1 mg/mL), was mixed with 2.5 mL of 0.2 N Folin-Ciocalteu’s phenol reagent. After 5 min incubation, 2.0 mL sodium carbonate solution (7.5% w/v in water) was added to the mixture and mixed evenly. The reactive mixture was kept in the dark at room temperature for 30 min, and the absorbance was subsequently measured at 760 nm. The total phenolic content was calculated as tannic acid equivalents (mg [TAE]/g) on the basis of a standard calibration curve of tannic acid.
The aluminum chloride colorimetric method [11] was applied (with minor modifications) to evaluate the total flavonoid content. Briefly, 0.5 mL extracts (1 mg/mL) were mixed with 0.5 mL aluminum chloride solution (10% w/v in water). The reactive mixture was kept in the dark at room temperature for 30 min, after which the absorbance was measured at 420 nm. The total flavonoid content was calculated as quercetin equivalents (mg [QUE]/g) on the basis of a standard calibration curve of quercetin.
Determination of crude saponin content
The total saponin content of the extracts was determined according to the method described by Chua et al. [12], with slight modification. A total of 0.1 mL of the extracts (1 mg/mL) was mixed with 0.3 mL of vanillin solution (8% w/v in ethanol) and 4 mL of sulfuric acid (72% w/v in water). The reactive mixture was allowed to stand at room temperature for 20 min, and the absorbance was then measured at 545 nm. A standard calibration curve was constructed using ginsenoside-Re as the standard saponin. The crude saponin content was expressed as ginsenoside-Re equivalents (mg [GE]/g).
Determination of platycodin D content
HPLC analysis was performed to analyze the levels of platycodin D in the extracts. For HPLC analysis, PGE, HFPGE, and platycodin D used as a reference standard were dissolved in methanol and filtered through 0.45 μm filter. Both PGE and HFPGE were analyzed using an HPLC system (A1100; Agilent Technologies, Santa Clara, CA, USA) with YMC Triart C18 column (250 × 4.6 mm, 5 μm; YMC Co., Kyoto, Japan). The diode array detector was set at 203 nm. The mobile phase solvents were water (A) and acetonitrile (B). The gradient elution program was as follows: (A) 20% and (B) 80% in 0–15 min, (A) 50% and (B) 50% in 15–20 min, (A) 20% and (B) 80% in 20–40 min. The sample injection volume was 20 μL with the flow rate set to 1.0 mL/min.
Cell culture and cell viability assay
RAW 264.7 murine macrophages were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in DMEM supplemented with 10% FBS, 100,000 U/L penicillin, and 100 mg/L streptomycin in an incubator maintained at 37°C and humidified atmosphere of 5% CO2/95% air. To investigate the effect of PGE and HFPGE on cell viability, RAW 264.7 cells were plated at a density of 1 × 105 cells/well in a 24-well plate. After 24 h incubation, cells were treated with varying concentrations (0–500 μg/mL) of PGE or HFPGE, and incubated for further 48 h. Cell viability was measured by the MTT assay, as reported previously [13].
Determination of phagocytic activity
The phagocytic activity of RAW 264.7 cells was measured using the Phagocytosis assay (zymosan substrate) Kit (Abcam, Cambridge, England), following the manufacturer’s instructions. Briefly, cells were plated at density of 1 × 105 cells/well in a 96-well plate. After 24 h incubation, cells were treated with varying concentrations (0–50 μg/mL) of PGE or HFPGE and incubated for 24 h. Cells treated with lipopolysaccharide (LPS, 0.1 μg/mL) were used as the positive control and cultured under the same conditions as other groups. Subsequently, 10 μL pre-labeled zymosan suspension was added per well as a pathogen for triggering phagocytosis, followed by incubation at 37°C for 1 h. Cells were thereafter fixed, blocked, and subjected to permeabilization. The reaction was initiated by adding 100 μL/well of substrate, followed by incubation at 37°C for 20 min. The absorbance was then measured at 405 nm.
Determination of nitric oxide (NO) production
RAW 264.7 cells were plated at a density of 1 × 105 cells/well in a 24-well plate and incubated for 48 h. Cells were then serum-starved with serum-free DMEM for 2 h, followed by treatment with the indicated concentration of LPS (0.1 μg/mL) or HFPGE (0, 5, 10 or 50 μg/mL) in serum-free DMEM for 24 h. The 24-h conditioned media were collected, and the nitrite contents were determined using the Griess reagent system (Promega, Madison, Wisconsin, USA), following the manufacturer’s instructions.
Determination of cytokines production
Cells were treated with LPS (0.1 μg/mL) or HFPGE (0, 5, 10 or 50 μg/mL), after which the 24-h conditioned media were collected as described above. The 24-h conditioned media were thereafter applied to a proteome profilerTM mouse cytokine array kit to investigate changes in the expressions of cytokines subsequent to LPS or HFPGE exposure, in accordance with the manufacturer’s instructions. The relative abundance of each protein spot was quantified using an ImageQuantTM LAS500 imaging system (GE Healthcare Bio-Sciences AB, Uppsala, Sweden). The expression levels were normalized to the reference protein spots, after which the levels of TNF-α, IL-1β, IL-6, CXCL10, G-CSF, GM-CSF, and MCP-1 were measured using the relevant ELISA kits, following the manufacturer’s instructions.
Quantitative real-time reverse transcription-polymerase chain reaction (RT-PCR)
By following the manufacturer’s protocol, total RNA was extracted with Trizol after 24 h treatment with LPS (0.1 μg/mL) or HFPGE (0, 5, 10 or 50 μg/mL). The content and purity of total RNA were determined using a micro-volume spectrophotometer (BioSpec-nano; Shimadzu, Kyoto, Japan). Complementary DNA was synthesized from 2 μg of total RNA using the HyperScriptTM RT master mix kit. Real-time PCR of complementary DNA was conducted using a Rotor-Gene 3000 PCR (Corbett Research, Mortlake, Australia) and a Rotor-GeneTM SYBR Green kit, according to the manufacturer's instructions. The primer sequences used were as follows: inducible nitric oxide synthase (iNOS), sense, 5’-GGCAGCCTGTGAGACCTTTG-3’, antisense, 5’-CATTGGAAGTGAAGCGTTTCG-3’; TNF-α, sense, 5’-ACTGAACTTCGGGGTGATTG-3’, antisense, 5’-GCTTGGTGGTTTGCTACGAC-3’; IL-1β, sense, 5’-CACCTTCTTTTCCTTCATCTTTG-3’, antisense, 5’-GTCGTTGCTTGTCTCTCCTTGTA-3’; IL-6, sense, 5’-TGATGGATGCTTCCAAACTG-3’, antisense, 5’-GAGCATTGGAAGTTGGGGTA-3’; CXCL10, sense, 5’-GACGGTCCGCTGCAACTG-3’, antisense, 5’-CTTCCCTATGGCCCTCATTCT-3’; G-CSF, sense, 5’-ATCCCGAAGGCTTCCCTGAGTG-3’, antisense, 5’-AGGAGACCTTGGTAGAGGCAGA-3’; GM-CSF, sense, 5’-ACCACCTATGCGGATTTCAT-3’, antisense, 5’-TCATTACGCAGGCACAAAAG-3’; MCP-1, sense, 5’-CCCAATGAGTAGGCTGGAGA-3’, antisense, 5’-TCTGGACCCATTCCTTCTTG-3’; glyceraldehyde 3-phosphate dehydrogenase (GAPDH), sense, 5’-AGGTTGTCTCCTGCGACT-3’, antisense, 5’-TGCTGTAGCCGTATTCATTGTCA-3’. The amplification reaction was run to complete 40 cycles at 95°C for 20 s, 60°C for 20 s, and 72°C for 20 s. The results were analyzed with the Rotor-Gene 6000 series System Software program, version 6 (Corbett Research). The relative expressions of target genes were normalized to the expression of GAPDH.
Western blot analysis
RAW 264.7 cells were seeded and treated with LPS (0.1 μg/mL) or HFPGE (0, 5, 10 or 50 μg/mL) for 16 h, as described above, followed by subjecting the cells to lysis, as described in the previous paper [14]. Protein contents of the cell lysates were determined using a BCA protein assay kit (Thermo Scientific, Rockford, IL, USA). Western blot analyses were conducted as reported previously [14]. Blots were visualized with LuminataTM Forte Western HRP Substrate (Millipore Corporation, Billerica, MA, USA). The relative protein abundance was quantified using an ImageQunatTM LAS 500 imaging system (GE Healthcare Bio-Sciences AB). Each protein expression level was normalized to β-actin.
Statistical analyses
Statistical analysis was performed with the SAS for Windows version 9.1 software (SAS Institute, Cary, NC, USA). All results are presented as the mean ± SEM and analyzed by analysis of variance. Differences between the treatment groups were analyzed by one-way analysis of variance and Duncan’s multiple range test. Differences are considered significant at P < 0.05.
RESULTS
HFPGE has higher contents of bioactive compounds
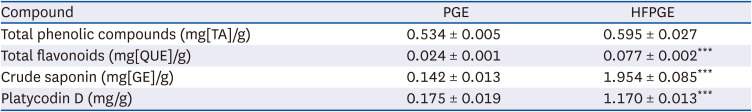
To investigate the effect of hydrolysis and fermentation processes on the bioactive compound contents of PGE, we measured the total phenolic compounds, flavonoids, crude saponin, and platycodin D contents in PGE and HFPGE. As presented in Table 1, significantly higher contents of total flavonoid, crude saponin, and platycodin D were obtained in HFPGE than PGE. However, the total phenolic content was similar in both HFPGE and PGE. This indicates that hydrolysis and fermentation processes increase the bioactive compound contents of the PGE.
Table 1
Total phenolic compounds, flavonoids, crude saponin, and platycodin D in P. grandiflorum extract
HFPGE promotes cell growth in RAW 264.7 cells
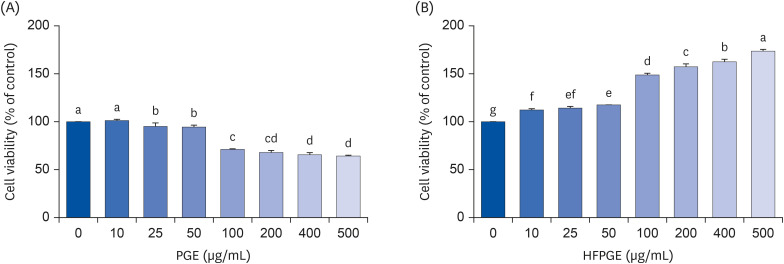
To investigate the effect of PGE and HFPGE on the growth of macrophages, we evaluated the cell viability by performing the MTT assay in RAW 264.7 cells. PGE significantly decreases the cell viability. In contrast, HFPGE treatment resulted in a significant and dose-dependent increase in the cell viability (Fig. 1). This indicates that HFPGE exerts an immunostimulatory effect via increasing the RAW 264.7 cell growth.
Fig. 1
Effect of PGE and HFPGE on the viability of RAW 264.7 cells. Cells were seeded at density of 1 × 105 cells/well in 24-well plates and incubated in DMEM supplemented with 100 mL/L FBS. After 24 h, cells were treated with varying concentrations of PGE (A) and HFPGE (B), and incubated for 48 h. Cell viability was determined by the MTT assay. Each bar expresses the mean ± SEM (n = 6). Means without the same letter differ, P < 0.05.
DMEM, Dulbecco’s Modified Eagle’s Medium; FBS, fetal bovine serum; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; PGE, Platycodon grandiflorum extract; HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract.
HFPGE increases the phagocytic activity of RAW 264.7 cells
Since phagocytosis is a representative immune response of macrophages, we determined the effect of PGE and HFPGE on the phagocytic activity to investigate the influence on the immune response. PGE did not affect the phagocytic activity of RAW 264.7 cells in the 0–10 μg/mL concentration range. However, at 50 μg/mL, treatment with PGE resulted in a significant increase in the phagocytic activity (Fig. 2A). HFPGE up-regulated the phagocytic activity of RAW 264.7 cells in a dose-dependent manner, with a remarkable increase (61%) being observed at 50 μg/mL, compared to the untreated control group. The degree of increased phagocytic activity after treatment with 50 μg/mL HFPGE showed no significant difference from the increase in phagocytic activity after treatment with 0.1 μg/mL LPS, a positive control (Fig. 2B).
Fig. 2
Effect of PGE and HFPGE on the phagocytic activity in RAW 264.7 cells. Cells were seeded at density of 1 × 105 cells/well in a 96-well plate and incubated. After 24 h, cells were treated with varying concentrations of PGE (A) and HFPGE (B), and incubated for 24 h. The phagocytic activity was measured using the Phagocytosis assay (zymosan substrate) kit. Each bar expresses the mean ± SEM (n = 6). Means without the same letter differ, P < 0.05.
PGE, Platycodon grandiflorum extract; HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; LPS, 0.1 μg/mL lipopolysaccharide.

HFPGE regulates NO production and iNOS expression in RAW 264.7 cells
NO, a non-specific effector molecule, plays a crucial role in attacking various pathogenic microorganisms. We therefore investigated whether HFPGE promotes NO production, and observed that HFPGE regulates NO production in RAW 264.7 cells. Treatment of cells with 50 μg/mL HFPGE significantly increased the NO production, but no effect was observed at 5 or 10 μg/mL treatment (Fig. 3A).
Fig. 3
Effect of HFPGE on NO production and iNOS mRNA expression in RAW 264.7 cells. Cells were seeded at a density of 1 × 105 cells/well in a 24-well plate and incubated. After 48 h, cells were serum-starved with serum-free DMEM for 2 h, and treated with the indicated concentration of HFPGE in serum-free DMEM for 24 h. The 24-h conditioned media were collected. (A) NO production was determined using the Griess reagent system. (B) Total RNA was extracted, reverse-transcribed, and real-time PCR was performed. iNOS mRNA expression was normalized to GAPDH and presented relative to the 0 μg/mL group. Each bar expresses the mean ± SEM (n = 6). Means without the same letter differ, P < 0.05.
DMEM, Dulbecco’s Modified Eagle’s Medium; HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; LPS, 0.1 μg/mL lipopolysaccharide; NO, nitric oxide; iNOS, inducible nitric oxide synthase; PCR, polymerase chain reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
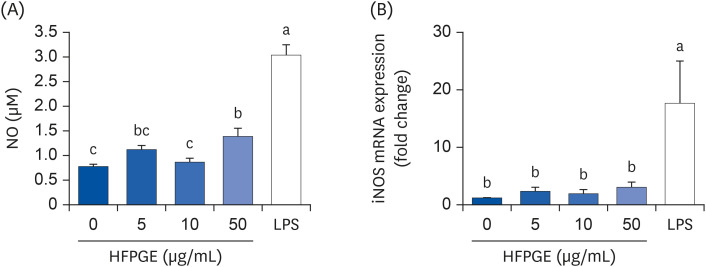
Since iNOS is responsible for most NO synthesis, we next investigated the effect of HFPGE on the expression of iNOS mRNA. As presented in Fig. 3B, treatment with HFPGE resulted in up-regulated iNOS mRNA expression. However, no significant difference was obtained in the iNOS mRNA expression levels among the HFPGE-treated groups at different concentrations (Fig. 3B).
HFPGE promotes the production and mRNA expression of cytokines in RAW 264.7 cells
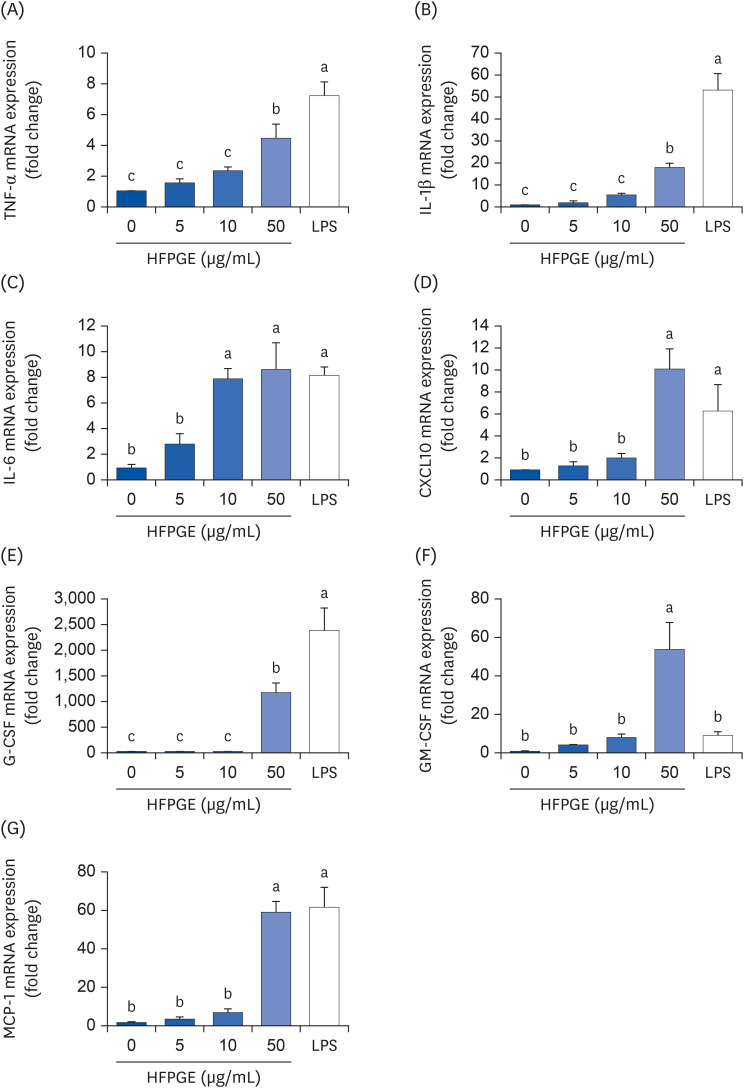
Multiplex cytokine assay was performed using the proteome profilerTM mouse cytokine array kit to screen alterations in cytokine production by HFPGE treatment. Among the 40 cytokines detectable with this kit, the levels of TNF-α, IL-1β, IL-6, CXCL10, G-CSF, GM-CSF, and MCP-1 were remarkably increased in the conditioned media of cells treated with 50 μg/mL HFPGE (data not shown). To confirm these results, ELISA assays were subsequently performed for TNF-α, IL-1β, IL-6, CXCL10, G-CSF, GM-CSF, and MCP-1. As shown in Fig. 4, the levels of TNF-α, IL-1β, IL-6, CXCL10, G-CSF, GM-CSF, and MCP-1 in the conditioned media were significantly increased subsequent to HFPGE treatment. Compared to the untreated control group, the increase in levels obtained after treatment with 50 μg/mL HFPGE were: IL-1β 2.41 times, IL-6 13.44 times, TNF-α 13.75 times, CXCL10 1.3 times, G-CSF 11.07 times, GM-CSF 2.1 times, and MCP-1 1.74 times (Fig. 4). Consistent with these results, HFPGE treatment markedly increased the mRNA expressions of TNF-α, IL-1β, IL-6, CXCL10, G-CSF, GM-CSF, and MCP-1 (Fig. 5).
Fig. 4
Effect of HFPGE on various cytokines production in RAW 264.7 cells. Cells were seeded and treated with HFPGE as described in Fig. 3. Levels of (A) TNF-α, (B) IL-1β, (C) IL-6, (D) CXCL10, (E) G-CSF, (F) GM-CSF, and (G) MCP-1 in 24 h-conditioned media were measured using the relevant ELISA kit. Each bar expresses the mean ± SEM (n = 6). Means without the same letter differ, P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; LPS, 0.1 μg/mL lipopolysaccharide; TNF, tumor necrosis factor; IL, interleukin; CXCL, C-X-C motif chemokine ligand; G-CSF, granulocyte-colony-stimulating factor; GM-CSF, granulocyte-macrophage colony-stimulating factor; MCP, monocyte chemoattractant protein; ELISA, enzyme-linked immunosorbent assay.
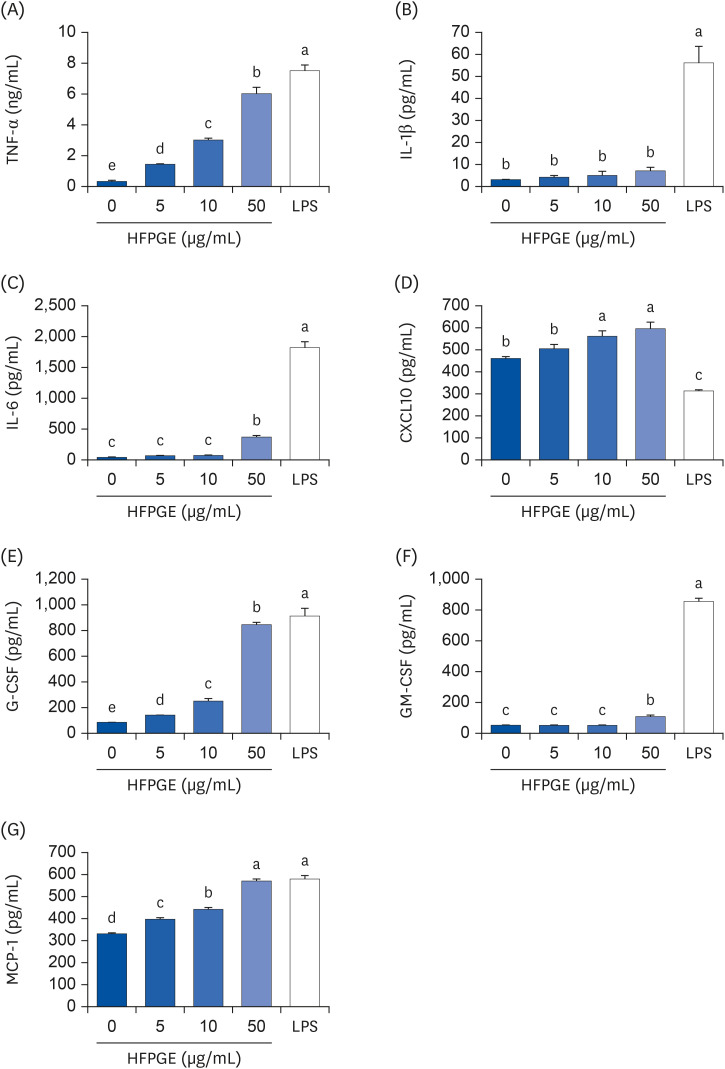
Fig. 5
Effect of HFPGE on mRNA expressions of various cytokines in RAW 264.7 cells. Cells were seeded and treated with HFPGE as described in Fig. 3. Total RNA was extracted, reverse-transcribed, and real-time PCR was performed. The mRNA expressions of (A) TNF-α, (B) IL-1β, (C) IL-6, (D) CXCL10, (E) G-CSF, (F) GM-CSF, and (G) MCP-1 were normalized to GAPDH and presented relative to the 0 μg/mL group. Each bar expresses the mean ± SEM (n = 6). Means without the same letter differ, P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; LPS, 0.1 μg/mL lipopolysaccharide; PCR, polymerase chain reaction; TNF, tumor necrosis factor; IL, interleukin; CXCL, C-X-C motif chemokine ligand; G-CSF, granulocyte-colony-stimulating factor; GM-CSF, granulocyte-macrophage colony-stimulating factor; MCP, monocyte chemoattractant protein; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
HFPGE stimulates MAPK and NF-κB signaling pathways in RAW 264.7 cells
To further elucidate the molecular mechanisms underlying the up-regulation of immune mediators, we investigated the effect of HFPGE on MAPK and NF-κB activation by Western blot analysis. As shown in Fig. 6, HFPGE significantly increases the phosphorylation of p-65, ERK, and JNK. In addition, the ratio of p-p65/p65, p-ERK/ERK, and p-JNK/JNK were also increased by HFPGE treatment (Fig. 6).
Fig. 6
Effect of HFPGE on MAPK and NF-κB signaling pathways in RAW 264.7 cells. Cells were seeded and treated with HFPGE as described in Fig. 3. Cell lysates were analyzed by Western blotting using the corresponding antibodies. (A, C, E) Images of chemiluminescent detection of the blots, which are representative of 3 independent experiments. The relative expression ratio of p-p65 to p65 (B), p-ERK to ERK (D), and p-JNK to JNK (F). Protein band were quantified, with the control levels set at 100. Each bar expresses the mean ± SEM (n = 3). Means without the same letter differ, P < 0.05.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; LPS, 0.1 μg/mL lipopolysaccharide; NF, nuclear factor; ERK, extracellular signal-regulated kinase; JNK, c-Jun N-terminal kinase.

DISCUSSION
PG contains triterpenoid saponins, flavonoids, phenolic acids, polyacetylene, sterols, and other compounds such as inulin, grandoside, 8 polysaccharides, starch grains, fatty acids, inorganic elements, and amino acids. The main active ingredient of PG is saponin. All saponins isolated from PG are oleanane-type pentacyclic triterpenoid saponins, and about 70 species have been identified so far. According to the sapogenins present, the saponins are divided into 5 types of platycodins (A–E). They have varied structures and a wide range of biological and pharmacological activities, including apophlegmatic, antitussive, immunostimulatory, anti-inflammatory, anti-oxidant, anti-tumor, anti-diabetic, anti-obesity, hepatoprotective, and cardiovascular activities [3].
To maximize the bioactive components, the HFPGE used in this study was fermented with L. casei for 24 h. In addition, a 3 h process of hydrolysis with bromelain and Celluclast, a protein-digesting enzyme mixture, was added before fermentation. Treatment with the resultant HFPGE increased the total flavonoids, crude saponin, and platycodin contents by 3.21 times, 13.76 times, and 6.69 times, respectively, compared to PGE (Table 1). In the study of Kim et al. [15], in which PG was fermented with a 1:1 mixture of Lactobacillus plantarum (P1201) and Lactobacillus brevis (BMK184) for 5 days, the contents of flavonoids, phenolic acid, and platycodin were increased 3 times, 5.24 times, and 8.67 times, respectively. Compared to this, the contents of crude saponin and platycodine were significantly increased in our study. Several fungi and enzymes can be used to ferment PG. Among them, Lactobacillus fermentation is known to be the most effective. Lactic acid bacteria are non-invasive and non-pathogenic gram-positive symbiotic microorganisms, and are recognized as health-promoting [16]. Lactobacillus has a strong glucosidase activity [17] which is largely effective in increasing the active ingredient by modifying the physiologically active compound to increase the content of available saponin [6]. Unlike the study of Kim et al. [15] which reported that the content of phenolic acid in HFPGE was significantly increased, we obtained no significant difference in total phenolic compounds content between HFPGE and PGE (Table 1). This may be due to the difference in the type of Lactobacillus, and also because the content of bioactive compounds was changed by hydrolysis. By including the hydrolysis process to the extraction process, we expect an increased extraction of active ingredients and the generation of new active ingredients. In fact, Li et al. [18] detected a new oleanane-type triterpenoid saponin by acid hydrolysis of the PG root. In addition, polysaccharides of PG are broken down into monosaccharides through hydrolysis, which helps the fermentation process thereafter.
In the current study, the non-cytotoxic concentration of PGE was determined to be 50 µg/mL, whereas HFPGE exerted no cytotoxicity even at 500 µg/mL (Fig. 1). This confirms that PG subjected to the fermentation process results in an increase in the contents of active ingredients and reduced cytotoxicity. Kim et al. [15] reported that while PG showed cytotoxicity at a concentration of 0.5 mg/mL, fermented PG showed no cytotoxicity even at 5 mg/mL. A study by Jung et al. [19] investigated the acute and 13 weeks sub-chronic toxicity in SD rats. They reported that the no observed adverse effects level of HFPGE was 3,000 mg/kg, indicating that HFPGE has a safe preclinical profile.
Treatment of RAW 264.7 cells with 0–50 μg/mL PGE or HFPGE revealed that the phagocytic activity of PGE was significantly increased only at a 50 μg/mL, whereas treatment with HFPGE showed increased activity at concentrations as low as 5 μg/mL, and showed a dose-dependent increasing trend (Fig. 2). Immunostimulatory activity of PG has also been reported in previous studies. PG was reported to promote the proliferation of macrophages and increase phagocytosis [2021]. In order to investigate the mechanism of HFPGE’s phagocytosis-promoting action, we measured the changes in NO production and iNOS expression in RAW 264.7 cells treated with HFPGE. HFPGE significantly increased the NO production at 50 μg/mL. However, the mRNA expression of iNOS showed no significant difference at HFPGE concentrations of 0–50 μg/mL, but showed a tendency to slightly increase in proportion to the concentration (Fig. 3).
Results of a cytokine array revealed that RAW 264.7 cells treated with 50 μg/mL of HFPGE showed relatively high changes in the secretions of certain cytokines, including TNF-α, IL-1β, IL-6, CXCL10, G-CSF, GM-CSF, and MCP-1 (data not show). Through a follow-up experiment, we found that the secretion of these cytokines increased in a dose-dependent manner subsequent to HFPGE treatment (Fig. 4). Moreover, RT-PCR confirmed that the mRNA expressions of these proteins were significantly increased (Fig. 5). Macrophages secrete various proinflammatory cytokines, anti-inflammatory cytokines, and chemokines. Lee et al. [9] reported that levels of TNF-α, IL-1β, and IL-6 were increased after treatment with extract of PG fermented with yeast (Saccharomyces cerevisiae). It is well-known that the immunostimulatory effect of PG is generally caused by pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6. However, in the current study, it is meaningful to reveal that in addition to these cytokines, the production of other proinflammatory cytokines and chemokines, such as CXCL10, G-CSF, GM-CSF and MCP-1, is also increased. The colony-stimulating factor G-CSF is secreted from other immune cells and epithelium as well as from macrophages, and stimulates the survival, proliferation, differentiation, and function of neutrophils. It is applied in the treatment for neutropenia, and recently it has been reported that G-CSF reduces inflammation, amyloid beta burden, and cognitive impairment in an experimental animal model of Alzheimer’s disease [22]. GM-CSF is another known colony-stimulating factor. Whereas G-CSF specifically promotes neutrophil proliferation and maturation, GM-CSF is known to contribute to the defense of infection by influencing the activation of more diverse immune cells and their immune functions [23]. MCP-1, also called the chemokine (C-C motif) ligand 2, plays a role in attracting immune cells to the site of inflammation, and is implicated in several inflammatory diseases [24]. CXCL10, also called the interferon gamma (IFN-γ)-induced protein 10, is secreted from macrophages and other cells in response to IFN-γ. This helps the anti-inflammatory and anticancer action of various immune cells, including promoting chemoattraction of macrophages [25]. In the current study, we found that the production of various cytokines was increased after treatment of HFPGE, thereby confirming the possibility of the immunostimulatory effect of HFPGE in these cytokines.
It has previously been reported that macrophage activation of PG occurs through the TLR4/NF-κB signaling pathway, which activates MAPK and AP-1. Thus, multiple intracellular signaling pathways, including NF-κB and MAPK, are triggered by PG, eventually leading to the generation of reactive oxygen species, NO, and inflammatory cytokines/chemokines [26]. In our study, treatment with HFPGE resulted in dose-dependently increasing the phosphorylation of the MAPK family, including the NF-κB p65 subunit, ERKs, and JNK (Fig. 6). These results confirm that HFPGE exerts an immunostimulatory effect through the NF-κB-MAPK signaling pathway. This is schematically illustrated in Fig. 7.
Fig. 7
Summary of the effects of HFPGE on the immune response in RAW 264.7 cells.
HFPGE, hydrolyzed and fermented Platycodon grandiflorum extract; TNF, tumor necrosis factor; IL, interleukin; CXCL, C-X-C motif chemokine ligand; G-CSF, granulocyte-colony-stimulating factor; GM-CSF, granulocyte-macrophage colony-stimulating factor; MCP, monocyte chemoattractant protein; TLR, toll-like receptor; TRAF, tumor necrosis factor receptor-associated factor.
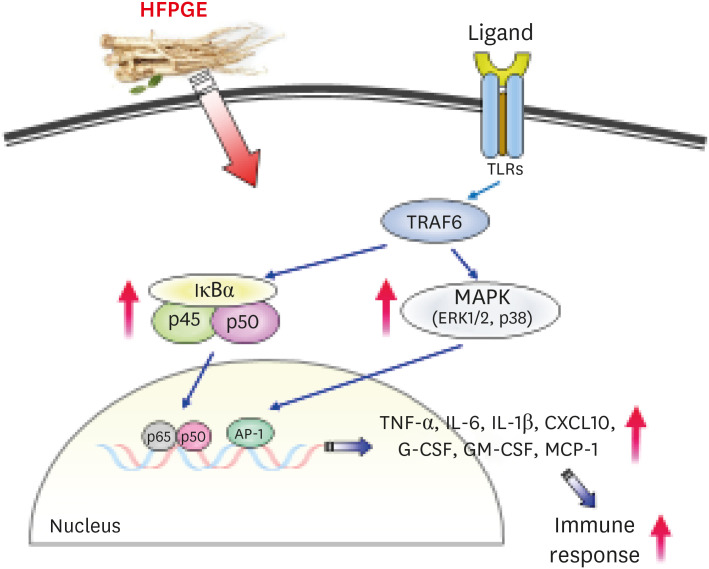
In conclusion, our data shows that HFPGE exerts lesser cytotoxicity and greater phagocytic activity than PGE. This bioactivity of HFPGE is determined to be due to increased NO production and increased secretion of cytokines such as TNF-α, IL-1β, IL-6, CXCL10, G-CSF, GM-CSF, and MCP-1 via the NF-κB and MAPK signaling pathways. These results indicate the potential to apply HFPGE as an effective immunostimulatory agent. In the future, additional studies are required to determine safe doses and the effects on applied diseases in animals and humans.
 XML Download
XML Download