

 PDF
PDF Citation
Citation Print
Print


Introduction
Owing to their ability to form dentin throughout life, odontoblasts play an important role in protecting tooth pulp from external stimuli. Odontoblasts undergo apoptosis after certain serious tooth injuries, following which new progenitor/stem cells then migrate to the injury site and differentiate into replacement odontoblasts, which synthesize reparative dentin (1, 2). These progenitor/stem cells are commonly known as dental pulp stem cells (DPSCs) (3), and odontogenic differentiation of DPSCs is an important step in the repair of pulp injury.
Changes in intracellular and extracellular pH can affect the differentiation of mesenchymal stem cells (MSCs). Small changes in extracellular pH leads to significant changes in osteogenic differentiation markers associated with bone marrow stromal cells (BMSCs); for instance, alkaline phosphatase activity and collagen synthesis decreased two-fold when pH was decreased from 7.5 to 6.6 (4). Cariporide contributed to the osteogenic differentiation of MSCs by inhibiting the activity of Na+/H+ exchange protein 1 (NHE1), which decreased intracellular pH (5). The pH of DPSCs may vary depending on the physiological and pathological conditions. For instance, the pH of carious dentin is significantly lower than that of normal dentin, with pH varying from 4.4 to 6.9 during the progression of caries (6). Mineral trioxide aggregate (MTA), a common material used for direct pulp capping, has an initial pH of 10.2, which increases to pH 12.5 after 3 h of mixing (7). The homeostasis of intracellular pH depends on transmembrane acid and base transporters, of which electrogenic sodium bicarbonate cotransporter 1 (NBCe1) is the most important (8). NBCe1 is a Na+-coupled HCO3− transporter encoded by the solute carrier 4A4 (SLC4A4) gene. This gene is expressed in tissues and cells throughout the body and plays crucial roles in maintaining intracellular and whole-body pH balance (9). Chen et al. (10) demonstrated that NBCe1 regulates intracellular pH, resulting in a resting intracellular pH higher than 7.2 in human DPSCs. Our previous research showed that SLC4A4 is highly expressed in odontoblasts and plays a crucial role in tooth development (11). Furthermore, interference with SLC4A4 expression by siRNA significantly inhibited DPSC migration, without a corresponding effect on proliferation (unpublished data).
Given the importance of NBCe1 in DPSCs, we used the compound S0859, a selective, high-affinity generic NBCe1 inhibitor, to block the activity of NBCe1 (12). The goal of this study was to investigate the effect of NBCe1 on the migration and osteogenic differentiation of DPSCs, and to explore the putative mechanism. In the present study, we demonstrate for the first time that the NF-κB pathway plays a key role in the NBCe1-mediated odontogenic differentiation of DPSCs, this may provide new clues for further interpreting the mechanism of DPSCs repair.
Materials and Methods
Isolation and culture of DPSCs
Samples were harvested from the third molars of patients aged 18∼25 years, with the informed consent of the patients. This study was approved by the Ethics Commit-tee of Huashan Hospital of Fudan University (Shanghai, China). DPSCs were isolated and cultured as described previously (13). Briefly, pulp tissues were removed on a clean bench, minced into approximately 1 mm3 sections, and then digested with 3 mg/ml collagenase type I (Sigma-Aldrich, USA) for 45 min at 37℃. After centrifugation at 1000 rpm for 5 min, DPSCs were cultured in a T25 flask with DMEM F12 (Hyclone, USA) supplemented with 10% fetal bovine serum (ScienCell, USA), 100 μg/ml streptomycin, and 100 U/ml penicillin (Gibco, USA). Cells were incubated at 37℃ with 5% CO2. Cell subculture was performed with 0.25% trypsin (Gibco) until 80% confluence. DPSCs were used for subsequent analyses after 3∼5 passages.
Flow cytometry
To identify isolated DPSCs, flow cytometry was performed for the following surface molecular markers of human MSCs: CD44-PE, CD73-APC, CD90-FITC, CD105-PerCP-CyTM5.5, CD34-PE, CD11b-PE, CD19-PE, CD45-PE, and HLA-DR-PE (BD Biosciences, USA). The DPSCs were then digested, resuspended, counted, and incubated with antibodies against the aforementioned molecular markers according to the manufacturer’s instructions at 37℃ in the dark for 30 min. After washing the cells twice with BD Pharmingen Stain Buffer, the cells were examined using a BD FACS Calibur system (BD Biosciences).
Alizarin red S
To investigate their multipotent differentiation potential, DPSCs were cultured in odontogenic and adipogenic induction medium. DPSCs were first seeded at a density of 3×104 cells/well into 6-well culture plates with complete culture medium. At 70% confluence, the cell medium was replaced with an osteogenic differentiation basal medium composed of α-MEM (Hyclone),10 nM dexame-thasone (Sigma-Aldrich), 10 mmol/l β-glycerophosphate, and 50 mg/ml ascorbic acid (Sigma-Aldrich). This medium was changed every 3∼4 days for 21 days. After removal from medium, the cells were gently washed with PBS three times and fixed with 4% paraformaldehyde for 30 min. After washing with distilled water, the cells were stained with 1% alizarin red S solution (pH 4.2); (Solarbio, China) for 30 min. Finally, the excess dye was removed and washed with distilled water. After drying, alizarin red was diluted in 10% cetylpyridinium chloride (CPC assay) to quantify mineralized nodules. Formation of mineralized nodules was observed using an inverted microscope with optical density determined at 562 nm.
Oil red O staining
For Oil Red O staining, DPSCs (2×105 cells/well) were first seeded in 6-well plates with growth medium until 100% confluence. The cells were then moved to a human DPSC adipogenic differentiation basal medium (Cyagen, China) containing glutamine, insulin, IBMX, rosiglitazone, and dexamethasone. This medium was changed at a schedule according to the manufacturer’s instructions. After 3∼4 cycles of induction/maintenance, the wells were washed with PBS, fixed in 4% paraformaldehyde, and stained with Oil Red O solution (Cyagen).
Cell counting kit-8
We performed a cell counting kit-8 assay (CCK8) to detect the cytotoxicity of S0859 (MedChemExpress, USA) to DPSCs. In brief, DPSCs were seeded in 96-well plates at a density of 2×103 cells/well. When the cells were completely adhered to the walls after 24 h, S0859 was added to the medium at a final volume of 100 μl/well at 1, 10, 30, 50, and 100 μM. After drug treatment for 0, 1, 3, 5, and 7 days, cells in each well were incubated with 10 μl of CCK-8 solution (Beyotime Biotechnology, China) for 3 h. Optical density (OD) was measured at 450 nm using a microplate reader (Biotek Instruments, USA). To further clarify the role of the NF-κB pathway in the odontogenic differentiation DPSCs, pyrrolidine dithiocarbamate (PDTC, a specific inhibitor of NF-κB) was added to suppress NF-κB activity. Before S0859 treatment at each concentra-tion, DPSCs were incubated with PDTC for 60 min. In a similar fashion, the effects of PDTC were analyzed by repeating the above-mentioned experiment with the addition of PDTC at concentrations of 1, 10, 100, 200, 400, 800, and 1000 nM at 0, 1, 3, 5, and 7 days. All solutions were fresh-ly prepared prior to use. The culture medium containing S0859 or PDTC was gently replaced every day, and 1-hour PDTC treatment was performed during every change.
Intracellular pH
The intracellular pH of DPSCs was assessed by confocal laser scanning microscopy using the intracellular pH fluorescent indicator BCECF-AM. Cells were washed with HEPES buffer solution and labeled with BCECF-AM (Invitrogen, USA). BCECF-AM was excited at 488 nm and emitted at wavelengths of 530 nm and 640 nm. The ratio of fluorescence intensity (530 nm/640 nm) reflects the intracellular pH. To obtain a calibration curve, intracellular pH was calibrated with Nigericin (Sigma, USA) to obtain a linear regression equation between pH 6.2 and 7.4. To illustrate the change in NBCe1 activity, the fluorescence intensity (530 nm/640 nm) of the cells treated with or without S0859 for 24 h was detected using BCECF-AM. Subsequently, the intracellular pH was calculated according to the standard curve.
Real-time polymerase chain reaction
Total RNA was extracted using an RNA-Quick Purifi-cation Kit (Esunbio, China). Approximately 1 μg of total RNA was used to synthesize complementary DNA (cDNA) using the PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa, Japan). Dentin sialophosphoprotein (DSPP), dentin matrix acidic phosphoprotein1 (DMP1), runt-related transcription factor 2 (RUNX2), osteopontin (OPN), and osteocalcin (OCN) were evaluated. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal reference. RT-PCR was performed using TB Green PCR master mix reagent (TaKaRa, Japan) in a Roche480 qPCR system (Roche, USA). Ct values were obtained for each sample. Relative gene expression was calculated using the 2−ΔΔCt method. Primer sequences are listed in Supplementary Table S1.
Western blotting
To investigate the expression of NF-κB pathway-related proteins, DPSCs were cultured in DMEM F12 medium and induced using S0859, PDTC or S0859+PDTC for 14 days. Total protein was extracted using a radioimmuno-precipitation assay lysis buffer (50 mM TrisHCl, pH 7.4, 150 mM NaCl, 1% NP40, 0.25% Na-deoxycholate) containing phenylmethanesulfonylfluoride (PMSF, Beyotime, China). Equivalent amounts of diluted protein samples were separated using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF, Sigma-Aldrich) membranes. Then, the membranes were blocked in 5% BSA (Beyotime Biotechnology, China) at room temperature for 1 h and incubated with the primary antibodies p65, p-p65, and GAPDH overnight at 4℃. After incubation with secondary antibodies at 4℃ for 1 h, the antibody-antigen reaction was detected using ECL western blotting substrate (Affinity, USA). The intensity of the target proteins was normalized to the GAPDH protein using Image-J1.52p.
Results
Characteristics of DPSCs
Most primary DPSCs had a fibroblast morphology, while some exhibited a spindle morphology (Fig. 1A). Pri-mary DPSCs were fused and grew in a swirl on the 13th day of primary culture (Fig. 1B). As expected, mineralized nodules revealed by Alizarin Red S staining demonstrated that DPSCs possessed an osteogenic/odontogenic differentiation capability (Fig. 1C and 1D). Similarly, Oil Red O staining demonstrated that DPSCs could be induced to differentiate into adipocytes (Fig. 1E and 1F). Flow cytometry revealed that the DPSCs showed positive expressions of MSC markers CD44, CD73, CD90, and CD105, and negative expressions of hematopoietic cell markers CD34, CD11b, CD19, CD45, and HLA-DR; Fig. 1G).

Fig. 1
Characterization of human dental pulp stem cells (DPSCs). (A) Primary DPSCs showed typical fibroblast or spindle morphology. (B) DPSCs were fused and grown in a swirl on the 13th day of primary culture (Scale bar=50 μm). (C) Control group for alizarin red S staining. (D) Alizarin red S staining of mineralized nodules after osteogenic-induced culture (Scale bar=200 μm). (E) Control group for oil red O staining. (F) Oil red O positive staining after adipogenic differentiation (Scale bar=200 μm). (G) Flow cytometry assay showing DPSCs with positive expression for CD44, CD73, CD90 and CD105, and negative expression CD34, CD11b, CD19, CD45 and HLA-DR.
![]()
Effects of NBCe1 on the proliferation of DPSCs
A CCK-8 assay was used to analyze the proliferation of DPSCs (Fig. 2A). Low concentrations of S0859 (1 μM and 10 μM) had no influence on cell proliferation, however, higher doses of S0859 (50 μM and 100 μM) decreased the proliferation of DPSCs after the first day. To avoid the cytotoxicity caused by high concentrations of S0859, we chose 10 μM S0859 for subsequent experiments.
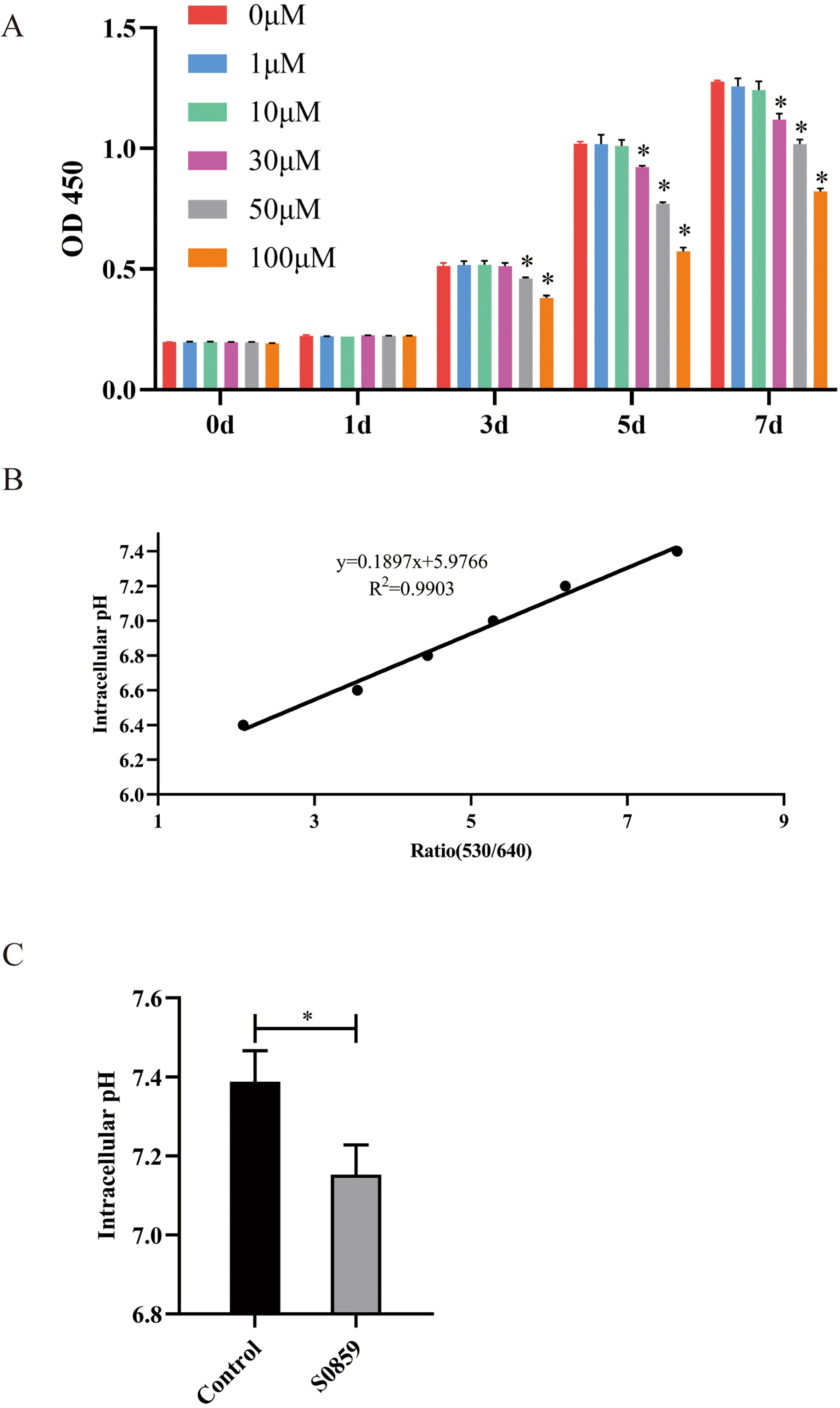
Fig. 2
Effect of S0859 on prolifera-tion and intracellular pH of DPSCs. (A) A CCK-8 assay revealed that low S0859 concentration had little effect on DPSCs viability, and a high dose inhibited proliferation (*p<0.05). (B) Standard curve for pH and its effect on DPSCs obtained using BCECF-AM. The pH/fluorescence ratio (530/640) conforms to the linear regression equation y=0.1897x+5.9766 (R2= 0.9903). (C) Effect of S0859 on intracellular pH of DPSCs observed using BCECF-AM. The intracellular pH of DPSCs treated with S0859 decreased compared to the control group (*p< 0.05).
![]()
Inhibition of NBCe1 decreased the intracellular pH of DPSCs
The nigericin calibration made the intercellular pH and the extracellular pH consistent. The fluorescence images at 530 nm, and 640 nm under different pH conditions (7.4, 7.2, 7.0, 6.8, 6.6, 6.4) were obtained (Supplementary Fig. S1). The fluorescence ratio of 530/640 was calculated under different pH conditions using ImageJ Pro Plus. The standard curve between pH and fluorescence ratio (Supplementary Fig. S1) fit the equation y=0.1897x+5.9766 (R2=0.9903; Fig. 2B). The cells were cultured for 24 hours after treatment with S0859. The ratio of 530/640 fluorescence intensity in the groups was analyzed, and the standard curve was used to calculate the intercellular pH. The intracellular pH of DPSCs was about 7.37, and the NBCe1 inhibitor S0859 decreased the intracellular pH of the DPSCs to about 7.16 (Fig. 2C, Supplementary Fig. S2). The results showed that S0859 treatment could decrease the intercellular pH by reducing the transport activity of NBCe1 in DPSCs.
NBCe1 inhibition promoted the odontogenic differentiation of DPSCs
To explore whether NBCe1 inhibition influenced the odontogenic differentiation of DPSCs, we induced odontogenic differentiation of DPSCs with or without S0859. Real-time PCR demonstrated that DSPP, DMP1, RUNX2, OCN, and OPN genes were upregulated (Fig. 3A) in the DPSCs cultured with S0859 compared to those in the negative (NC, control medium) or positive control group (MM, mineralized-inducing medium). More mineralized nodules were found in the MM+S0859 group compared to the MM group, whereas alizarin red staining was negative in the NC group (Fig. 3B). CPC assays showed significantly higher concentrations of mineralized nodules in the MM+S0859 group compared to the MM group (Fig. 3C). These results indicate that NBCe1 inhibition stimulates odontogenic differentiation of DPSCs.
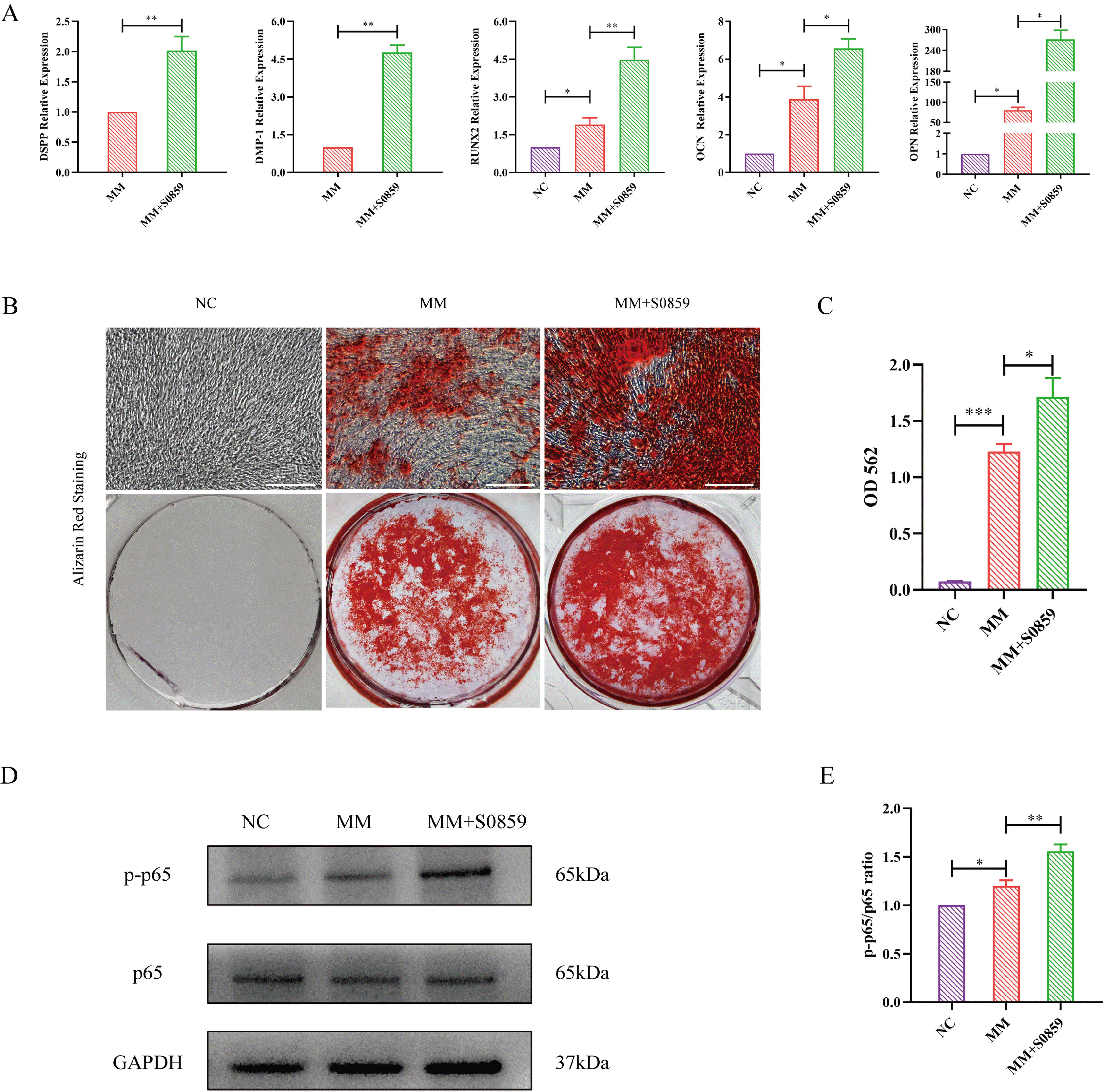
Fig. 3
NBCe1 inhibition promoted the odontogenic differentiation of DPSCs. (A) Real-time PCR indicated higher expressions of odontogenic genes (DMP1, DSPP, Runx2, OCN and OPN) in the MM+S0859 group compared to the NC or MM groups (**p<0.01; *p<0.05). (B) Alizarin red staining showed obvious mineralized nodules in the MM+S0859 group compared to the control group (Scale bar=50 μm). (C) A CPC assay quantified the mineralized nodules and revealed that the concentration of mineralized nodules was higher in the MM+S0859 group compared to the MM group (***p<0.001, *p<0.05). (D) Protein expression levels of p65 and p-p65. (E) The ratio of p-p65/p65 in the S0859-treated group was higher compared to the NC and MM group (*p<0.05, **p<0.01). NC: control medium, MM: mineralized-inducing medium, CPC: cetylpyridinium chloride.
![]()
NBCe1 inhibition activated the NF-κB pathway in DPSCs
Treatment with S0859 did not affect the expression of p65. The phosphorylation of p65 (p-p65) was upregulated by S0859 compared to that in the NC and MM groups. These results indicate that the p65-NF-κB signaling pathway may be involved in NBCe1 regulation of odontogenic differentiation of DPSCs (Fig. 3D and 3E).
Inhibition of NF-κB pathway attenuated odontogenic differentiation of DPSCs
The CCK8 assay showed that low doses of PDTC had no effect on the proliferation of DPSCs; however, 200 nM and 400 nM PDTC significantly inhibited cell prolifera-tion, and 800 nM and 1 μM PDTC were highly toxic to cells, even causing cell death (Supplementary Fig. S3). Therefore, we chose 100 nM PDTC for subsequent experiments. A western blotting assay was performed to detect the efficiency of 100 nM PDTC in the NF-κB pathway. After administration of PDTC, the phosphorylation of p65 was downregulated (Fig. 4D and 4E). Real-time PCR showed that DSPP, DMP1, Runx2, OCN, and OPN mRNA expression levels were lower in the S0859+PDTC group than in the S0859 group (Fig. 4A). Similarly, after 21 days of odontogenic differentiation, alizarin red staining showed that mineralized nodules significantly decreased in the S0859+PDTC group compared with that in the S0859 group (Fig. 4B). CPC assay showed that the calcium concentration in the S0859+PDTC group was lower than that in the S0859 group (Fig. 4C). These results show that the NF-κB pathway may be involved in the odontogenic differentiation of DPSCs.
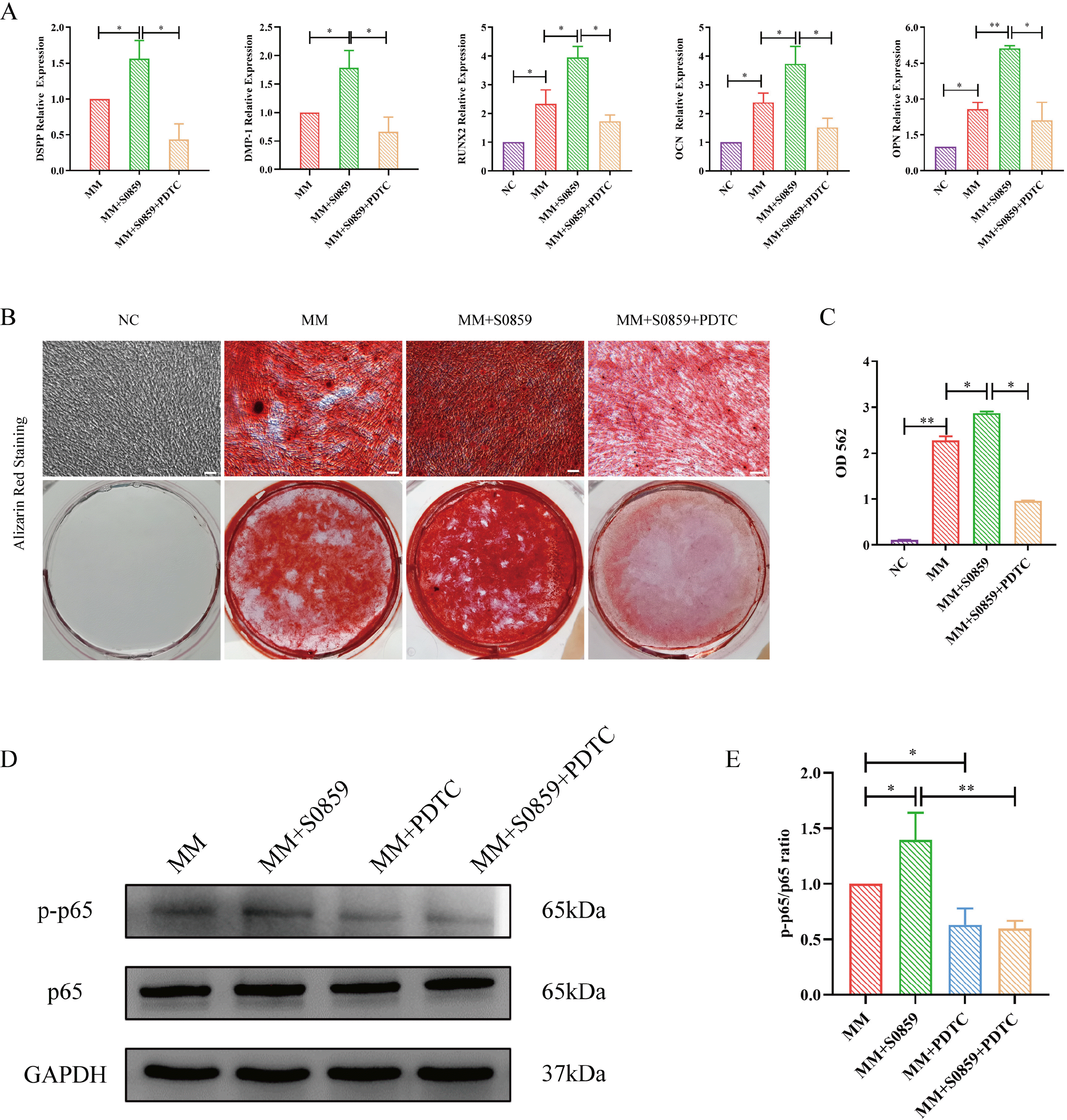
Fig. 4
Effect of NF-κB inhibitor on the odontogenic differentiation of DPSCs treated with S0859. (A) A Real-time PCR assay demonstrated that the mRNA expression levels of DSPP, DMP1, Runx2, OCN, and OPN in the S0859+PDTC group were decreased compared to that in the S0859 group (*p<0.05, **p<0.01). (B) Alizarin red staining of mineralized nodules was more obvious in the S0859 group than in the S0859+PDTC group (Scale bar=50 μm). (C) CPC quantification showed that the concentration of mineralized nodules in the S0859 group was higher than in the S0859+PDTC group (*p<0.05, **p<0.01). (D) Protein levels of p65 and p-p65 in the MM group, MM+S0859 group, MM+PDTC group, and MM+S0859+PDTC group. (E) Quantification of the p-p65/p65 ratios after inhibition of the NF-κB pathway (*p<0.05, **p<0.01). NC: control medium, MM: mineralized-inducing medium, CPC: cetylpyridinium chloride.
![]()
Discussion
In the present study, we demonstrate for the first time that the NF-κB pathway plays a key role in NBCe1-mediated odontogenic differentiation of DPSCs.
Almost all intracellular and extracellular biological pro-cesses are affected by pH, and maintaining acid-base homeostasis is crucial (14). In normally differentiated adult cells, intracellular pH is usually 7.2, versus the typical extracellular pH of 7.4 (15). However, cancer cells and stem cells have a higher intracellular pH and lower extracel-lular pH, and higher intracellular pH is important for maintaining the stemness of stem cells (5). Intracellular pH influences the proliferation and differentiation of various stem cells, including embryonic stem cells, induced pluripotent stem cells, cancer stem cells, and MSCs (16). Intracellular pH homeostasis is mainly regulated by pH-regulatory transporters in the plasma membrane (8). NBCe1 is an important member of the solute carrier 4 (SLC4) family and plays important roles in intracellular pH and HCO3− transmembrane transporters (17). NBCe1 is expressed in various mammalian tissues, and Lacruz et al. (18) demonstrated for the first time that NBCe1 expression is required for the development of normal dentition.
We used the selective NBCe1 inhibitor S0859 to explore the effect of NBCe1 on the differentiation of DPSCs. To avoid cytotoxicity, we used a low concentration of 10 μM. This concentration did not affect cell proliferation or differentiation but significantly decreased intracellular pH. Inhibition of NBCe1 upregulated the mRNA expression of DSPP, DMP1, RUNX2, OCN, and OPN, suggesting that NBCe1 is involved in the odontogenic differentiation of DPSCs and dentin formation. DSPP, a significant marker for odontogenic differentiation, is a non-collagen protein mainly expressed and secreted by odontoblasts (19). DMP1 is a unique molecule located in the nucleus that initiates osteoblast differentiation; during later stages of osteoblast maturation, phosphorylated DMP1 is transported from the nucleus to the extracellular matrix to regulate extracel-lular mineralized matrix formation (20). DMP1 and DSPP may be synthesized and secreted by DPSCs into the developing dentin matrix. We did not detect DSPP and DMP1 mRNA expression in the negative control group. Gronthos et al. (21) did not detect DSPP expression using normal growth medium and proposed that cultured DPSCs were in an undifferentiated state and negatively expressed the odontoblast-specific marker.
NF-κB transcription factors are critical regulators of immunity, stress responses, apoptosis, and cell differentia-tion (22), and are specifically involved in regulating odontogenic differentiation of DPSCs (23, 24). For instance, tumor necrosis factor α (TNFα) promotes the osteogenic differentiation of human dental pulp stem cells through the NF-κB signaling pathway (25). Meanwhile, MTA enhances the odonto/osteogenic capacity of DPSCs from inflammatory sites by activating the NF-κB pathway (26). The NBCe1 promoter region contains an NF-κB binding site, and NF-κB is responsive to changes in pH in rodent incisors (27). CFTR/HCO3− activation of miR-125b regulates early embryonic development through the sAC-PKA-NF-κB cascade (28). Tescalcin regulates cell prolifera-tion, migration, and invasion via NHE1/pH as well as the AKT/NF-κB signaling pathway in renal cell carcinoma (29). Consistent with these diverse effects, we hypothesized that NF-κB is involved in NBCe1-mediated DPSC odontogenic differentiation. NBCe1 inhibition upregulated NF-κB p65 phosphorylation, suggesting that NF-κB signaling plays an important role in this process. To further test this, we used PDTC, a specific inhibitor of the NF-κB signaling pathway (30). PDTC can decrease NF-κB p65 subunit protein expression and phosphorylation (31, 32). Blocking the NF-κB signaling pathway significantly suppressed the odontogenic effects of NBCe1 on DPSCs as well as NF-κB p65 phosphorylation. These results suggest that NBCe1 regulates odontogenic differentiation of DPSCs through the NF-κB p65 signaling pathway. We speculate that NBCe1 regulates the odontogenic differentiation of DPSCs by affecting HCO3− transport, which in turn affects HCO3− concentrations and the phosphorylation of NF-κB p65. Another possibility is that NBCe1 changes intracellular pH and thus indirectly affects the phosphorylation of NF-κB p65 (33). Finally, it is also possible that the NF-κB binding site on NBCe1 directly affects phosphorylation of NF-κB p65 (27), thereby regulating the entire process (Fig. 5). All these possibilities are excellent potential avenues for future research.
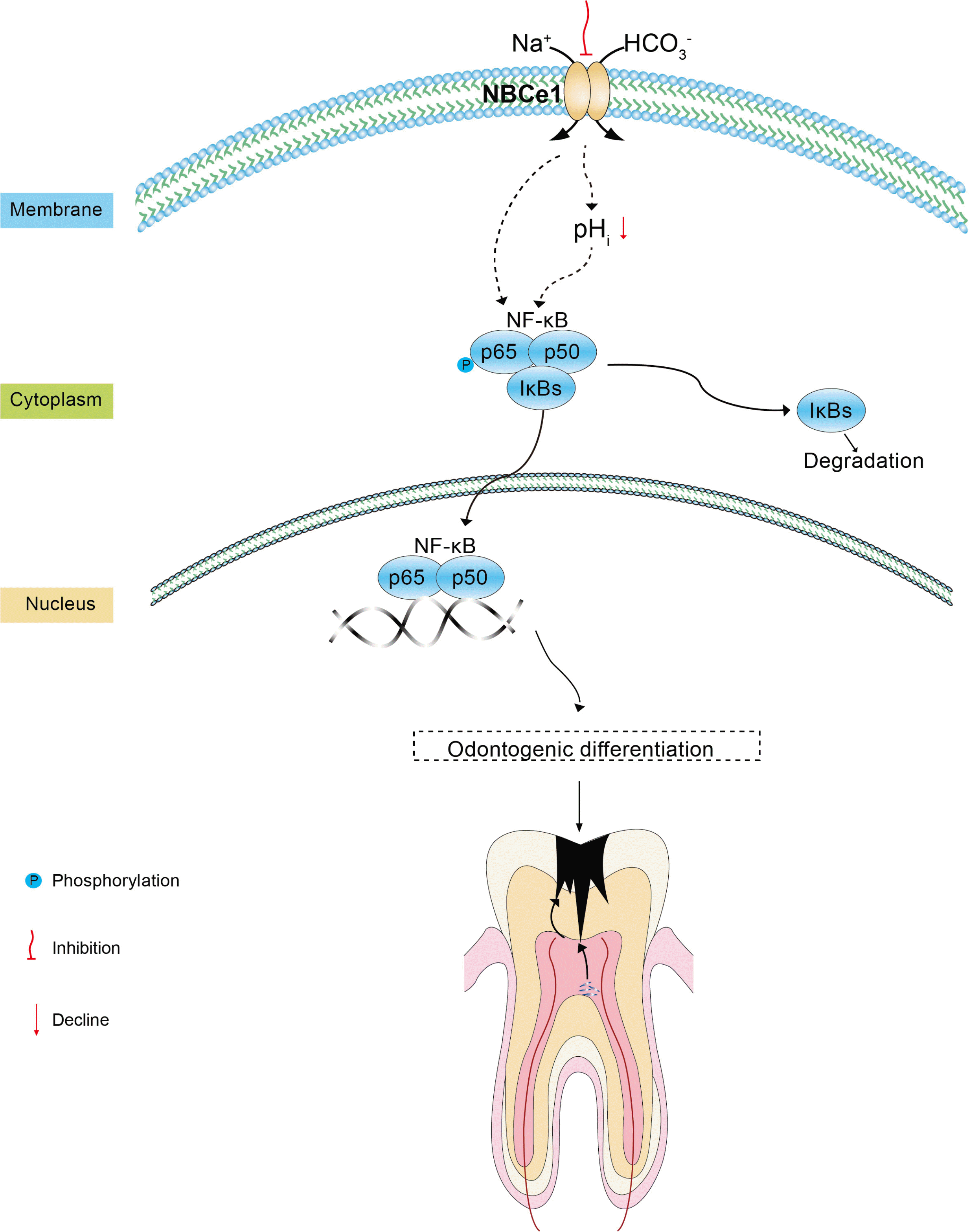
In conclusion, our study demonstrated that NBCe1 inhibition promoted the odontogenic differentiation of DPSCs by activating the NF-κB pathway. NBCe1 may play a crucial role in the odontogenic differentiation of DPSCs, and these findings may contribute to the development of clinical pulp therapy and our understanding of dental pulp regeneration.
Supplementary Materials
Supplementary data including one table and three figures can be found with this article online at https://doi.org/ 10.15283/ijsc21240.
 XML Download
XML Download