

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Aging is a time-dependent reduction of functional capacity in living organisms and is characterized by a steady loss of biological integrity, resulting in impaired function and increased mortality. The process of aging is based on four hallmarks, including; genomic instability, telomere attrition, epigenetic alterations and loss of proteostasis [1]. These ‘hallmarks’ can be caused by the generation of reactive oxygen and nitrogen species (ROS and RNS). ROS and RNS could lead to protein unfolding, lipid breakdown or peroxidation, and DNA damage [2]. In addition, ROS and RNS also cause a dysregulation of the immune response and development of metabolic syndrome resulting in chronic inflammation and oxidative stress [3].
Oxidative stress is a natural biological phenomenon that plays a significant role in aging [4]. It is a state of excess free radicals due to an imbalance between their production and accumulation in cells and tissues but also the inability of detoxification of free radicals [5]. Antioxidative defenses in vivo protect cellular macromolecules from the detrimental effects of these ROS that accumulate with age. The antioxidative defense system includes enzymatic antioxidants (i.e., superoxide dismutase [SOD], glutathione peroxidase [GPx], and catalase [CAT]), nonenzymatic antioxidants (i.e., glutathione [GSH] and thioredoxin), and dietary antioxidants (i.e., vitamin C, vitamin E, and selenium) [6]. The oxidative stress theory of aging is a theory that accounts for the role of varieties of ROS in controlling aging [7]. According to this theory, aging is a state of chronic oxidative stress that reduces the levels of antioxidant enzymes such as CAT, SOD and GPx [8] causing an “exhausted” state of the antioxidant defense system. Therefore, we hypothesized that lifestyle habits that reduce pro-oxidant ROS, boost antioxidant defenses or a mix of both approaches will ameliorate oxidative stress.
Many studies have proved that lifestyle plays an important role in maintaining the health of individuals throughout their lifespan. For example, high-fat diet (HFD) consumption has been reported to be a significant health risk factor. The consumption of HFD showed increased oxidative stress and dysfunctional mitochondria in many organs such as liver, kidney, skeletal muscle and heart [9101112]. Also, previous studies have shown that HFD can induce obesity and promote inflammation, leading to development of metabolic syndrome by increasing fasting blood glucose, dyslipidemia, and liver damage [13]. Many previous studies have indirectly connected HFD consumption to oxidative stress exists [1415]. However, there are a few studies regarding the direct association among HFD consumption, aging and oxidative stress. Thus, this study aimed to investigate the direct role of HFD on the induction of oxidative stress in aging mice.
Green perilla (Perilla frutescens) is an annual medicinal herb belonging to the mint family (Lamiaceae). It is commonly cultivated and used in Asian cuisines in countries such as South Korea, Thailand, Japan, and China [16]. Perilla seeds and leaves are used in traditional Asian medicine to treat cold and coughs, headaches, abdominal fullness and bloating, and food poisoning from fish and crabs [171819]. Previous research has shown perilla to exhibit a number of beneficial health effects, including anti-inflammatory [20], antiallergic [21] and glucosidase inhibitory activities [22], suggesting that perilla leaf could serve as an antioxidant booster to combat oxidative stress damages. Studies have demonstrated that perilla seeds and leaves contain phenolic and flavonoid antioxidant compounds such as catechins, caffeic acid, rosmarinic acid, apigenin and luteolin [23]. Rosmarinic acid in particular is reportedly a major polyphenol in perilla leaves and has been shown to exhibit several biological functions, including antioxidant activity [17222324].
There exist studies on perilla leaf and its antioxidant properties, however, the preventive effects of perilla on oxidative stress against aging and long term HFD intake are not fully investigated. In this study, we investigated whether green perilla leaf extract (PE) can exhibit inhibitory and protective effects against oxidative stress and damage resulting from increased lipid peroxidation induced by a HFD in aging mice via the improvements of the ROS scavenging activity, anti-inflammatory and antioxidant system.
Go to : 
MATERIALS AND METHODS
PE and freeze-drying
Perilla leaves were acquired from Wanju County, South Korea, washed and dried. The dried leaves (1.8 kg) were extracted with 70% ethanol and reflux extraction (5 L × 3 times). After filtration using a funnel, the total filtrate was concentrated on a rotary evaporator at 55°C to produce a condensed extract (482 g) and lyophilized at −125°C and 5 mTorr for 48 h using a freeze dryer (Ilshin Valve, Siheug, Korea). The dry weight of the final product was 304 g. Our study utilized 70% ethanol for extraction similar to the extraction conditions in our referenced study for the characterization of perilla leaves. The presence of water increases the polarity of the extraction agent (the dielectric constant of water is about 80 while ethanol is 25.2) and is used to swell the plant material, which increases the contact surface between the plant matrix and the solvent. As the proportion of organic solvent in aqueous solutions increases, the content of phenolic compounds decreases [25].
Antioxidant assay of DPPH radical scavenging activity of perilla extract in vitro
In this study, quercetin was used as reference to investigate the radical-scavenging activity of 1,1-diphenyl-2-picryl-hydrazyl (DPPH). Briefly, 0.2 g of plant extract and quercetin were each dissolved in 20 mL of reagent grade methanol in their respective flasks and stirred for 30 min using an automatic stirrer. The mixture was centrifuged at 6,000 rpm and 4°C for 10 min and the collected supernatant was used for the assay. Free-radical activity was then assessed by DPPH using a modified version of a method described by Bektas et al. [26]. A DPPH solution with a concentration of 0.1 mM was prepared and 0.5 mL of the solution was added to 1.5 mL of each supernatant solution at various concentrations (6.25, 12.5, 25, 50, and 0.01 mg/mL). The mixtures were vortexed and left to stand at room temperature for 30 min, after which the absorbance was measured at 517 nm using a spectrophotometer. The DPPH radical scavenging activity was calculated using the formula:
where A0 is the absorbance of the control reaction (i.e., without DPPH) and A1 is the absorbance of samples with DPPH present. Tests were conducted in duplicate and the average value was taken.
Assay of ABTS•+scavenging activity of perilla extract
ABTS•+ [2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid scavenging activity)] assay measures the antioxidant activity of lipid soluble and water-soluble antioxidants, and can be used for natural source extracts such as plant extracts. ABTS•+working solution was produced by reacting 38.43 mg ABTS (final concentration 7 mM) and 6.90 mg potassium persulfate (final concentration 2.55 mM) and 10 mL distilled H2O in a 15 mL tube and the mixture was left to incubate at room temperature in the dark for 12 h. The dark blue-green ABTS•+ solution was diluted with 100% ethanol till it gave an absorbance of 0.70 ± 0.02 at 734 nm. ABTS•+ adjusted solution (180 μL) and 100% ethanol (20 μL) was blank and absorbance at 734 nm corresponding to blank (E1) was recorded using a 96 well plate microplate. Then test sample (20 μL) at different concentrations 6.25, 12.5, 25, and 50 mg/mL was mixed with ABTS•+ adjusted solution (180 μL) per well in triplicates. The reaction mixtures were allowed to incubate in room temperature for 2 min (E2). The ABTS•+ scavenge % was calculated as:
The ABTS•+ scavenge % was plotted against the antioxidant extract concentration and the values were calculated.
Animals and diets
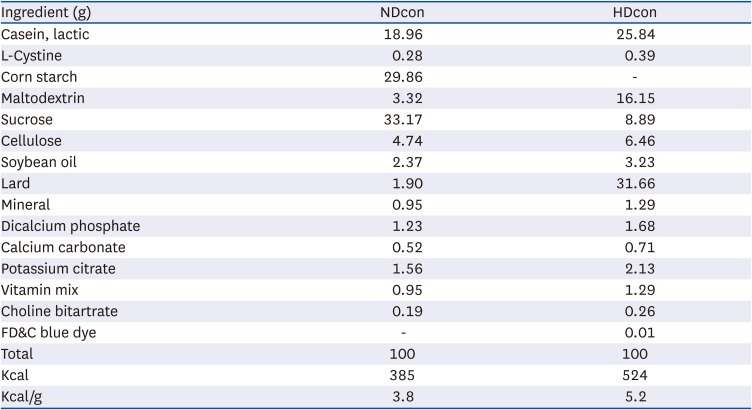
Four-week-old male C57BL/6J mice were purchased from Central Lab Animal Inc. (Seoul, South Korea), and the diets were purchased from Duyeolbiotech, (South Korea). Details of the diet compositions are listed in Table 1. After one week of adaptation, animals were divided equally into two groups. One group was fed a normal diet of 10% kcal fat (ND) and the other a 60% high-fat diet (HD) for 53 weeks before the start of the experiment. After which they were further divided into 3 groups of n = 8. The ND control group (NDcon), high-fat diet control group (HDcon), and the high-fat diet PE treated group (HDPE) given 400 mg mg/kg of bodyweight concentration of PE by oral gavage every morning daily for the whole experimental duration at 0.1 mL volume. Control groups (NDcon and HDcon) were given same volume of distilled water at the same frequency. The animals were then fed a normal diet and HFDs for another 12 weeks respectively. Fig. 1 details a graphical representation of the experimental model. The mice were housed in a temperature-regulated animal room with a 12-h light/dark cycle. PE in powdered form was diluted in water and administered orally daily to the treatment group and an equal volume of distilled water was administered orally to the control groups. The feed intake was measured twice a week and body weights were recorded weekly. The Animal and Use Committee of Jeonbuk National University approved the experimental protocol (CBNU 2020-026).
 | Fig. 1Graphical experimental model design in C57BL/6J mice.NDcon, normal diet fed reference group; HDcon, high-fat diet fed group; HDPE, high-fat diet PE treated group (400 mg/kg of body weight).
|
Table 1
Composition of experimental diets
Normal diet (ND group) is a modified diet containing 4% fat (10% fat calories) content. High-fat diet (HD group) is a modified high fat diet with 35% fat (60% fat calories).
NDcon, normal diet fed reference group; HDcon, high-fat diet fed group.
![]()
Serum and tissue sample collection
The mice were sacrificed after 12 h of fasting at the end of the experimental period. The mice were exposed to ether as anesthesia for 10 sec, and then blood for serum extraction was taken using capillary tubes via the retro-orbital vein leading to death by exsanguination. After which following ethical procedures liver tissue was harvested and dissected. Collected blood samples were left to stand for 30 min at room temperature. Serum was then separated by centrifugation at 1,500 g and 4°C for 15 min and stored at −72°C until analysis. Liver tissues were also collected and frozen in liquid nitrogen, and stored at −72°C for later analysis.
Fasting blood glucose concentrations
Fasting blood glucose was measured after fasting overnight before sacrifice. Blood glucose was measured using a commercial Roche Accu-Chek Performa blood-glucose meter and test strips (Roche Diabetes Care GmbH, Mannheim, Germany) and blood from the tail veins of the mice.
Lipid profiles in liver and serum
Total triglyceride (TG) and total cholesterol (TC), HDL cholesterol, aspartate aminotransferase (AST) and alanine aminotransferase (ALT) parameters were measured via an enzymatic process with commercially available detection kits. For biochemical analysis of hepatic lipid profiles such as TG and TC, and other lipid profiles, hepatic liver tissue homogenates were prepared; 0.1 g of liver tissue was weighed in a fresh 1.5 mL Eppendorf tube and 0.3 mL 1X PBS added. The tissues were chopped up finely and then homogenized with a sonicator. A 0.5 mL of Chloroform: Methanol (2:1) mixture was added and vortexed for 30 seconds. The mixture was then centrifuged at 8,000 rpm/15 min/4°C. The clear liquid bottom layer was then transferred to a fresh Eppendorf tube and kept overnight in a fume hood to vaporize the organic solvent. The resulting yellowish paste at the bottom of the tube was then dissolved with 0.2 mL 1% triton X (diluted with PBS) to the paste and mixed thoroughly till it completely dissolved. The dissolved mix was vortexed for 10 seconds and then used immediately for analysis of TG, TC and other lipid profile tests. LDL values were calculated using Friedewald’s equation.
Histology analyses
Same section of liver from each mice was harvested after dissection and washed with PBS then fixed in 10% formalin. Then each section was taken and processed for paraffin embedding. Preparation of paraffin sections of liver tissue samples as well as hematoxylin and eosin phloxine (H&E) staining was outsourced to KP&T (Cheongju, Korea). The stained areas were viewed later using an Axiophot Zeiss Z1 microscope (Carl Zeiss, Gottingen, Germany) at 100× magnification at Jeonbuk National University’s Center for University-Wide Research Facilities.
Biochemical analyses
Commercially available ELISA kits were used to carry out the biochemical analyses of serum tumor necrosis factor alpha (TNF-α) levels in serum and the oxidative-stress state in serum and livers. Commercial SOD, CAT, GPx, hydrogen peroxide (H2O2) and malondialdehyde (MDA) kits were purchased from an Abcam Biotechnology (Cambridge, UK) and utilized according to the stated protocols in the kit manuals.
Total mRNA isolation and qRT-PCR
To assess the expression of genes associated with oxidative stress, inflammation and antioxidant enzymes, quantitative real-time polymerase chain reaction (qRT-PCR) amplification was performed using liver tissues. Total RNA isolation was carried out using a Trizol reagent and concentration purity was checked with a BioDrop μLITE spectrophotometer (Biodrop, United Kingdom). One microgram of the isolated RNA was then reverse transcribed into cDNA using Takara PrimeScript RT Master Mix (Tokyo, Japan). The degree of RNA expression was then quantified by qRT-PCR using an SYBR Green Realtime PCR Master Mix and a 7500 Real-Time PCR system (Applied Biosystems, Foster City, CA, USA) following the manufacturer’s protocol. The sequences of primers used in this procedure are shown in Table 2.
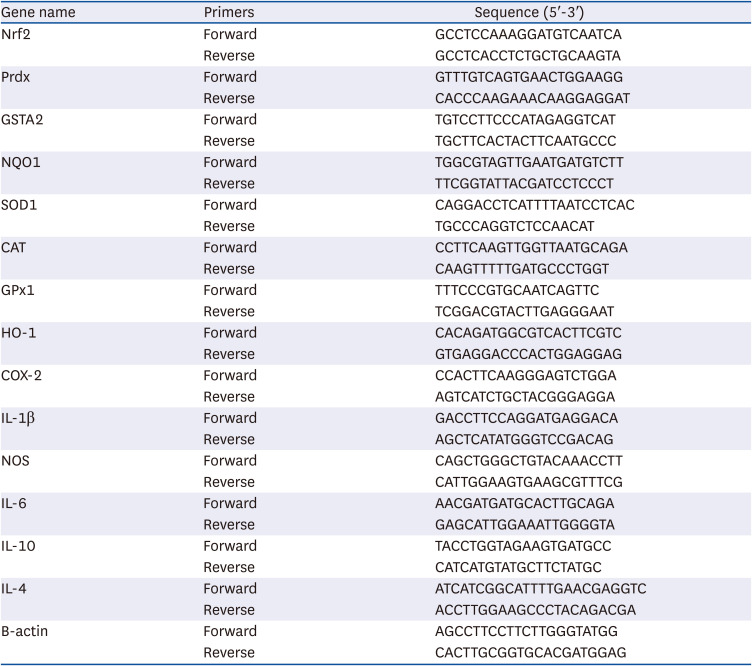
Table 2
Quantitative polymerase chain reaction primer sequences
Nrf2, nuclear factor erythroid 2-related factor 2; Prdx, peroxiredoxin; GSTA2, glutathione S-transferase alpha 2; NQO1, NAD(P)H Quinone Dehydrogenase 1; SOD1, superoxide dismutase 1; CAT, catalase; GPx1, glutathione peroxidase 1; HO-1, heme oxygenase 1; COX-2, cyclooxygenase-2; IL-1β, interleukin 1 beta; NOS, nitric oxide synthase; IL-6, interleukin 6; IL-10, interleukin 10; IL-4, interleukin 4; β-actin, beta actin.
![]()
Statistical analysis
Results are shown as the mean ± SE. Statistical analysis of all data was carried out using one-way analysis of variance and independent t-test using SPSS 17.0 (SPSS Inc., Chicago, IL, USA). Differences between the means were evaluated using Duncan’s multiple-range test with a P value < 0.05 considered statistically significant. Different superscripts or letters on the values indicate a significant difference between groups.
Go to :
RESULTS
DPPH scavenging activity of PE
The DPPH radical scavenging activity of PE and quercetin decreased dose-dependently (6.25, 12.5, 25, and 50 mg/mL). The PE scavenging capacity decreased dose-dependently. The scavenging values ranged from 66.2% to 50.7% for perilla and 101 to 97.1% for quercetin (Fig. 2A). Quercetin produced higher DPPH radical scavenging activity in all concentrations compared with perilla.
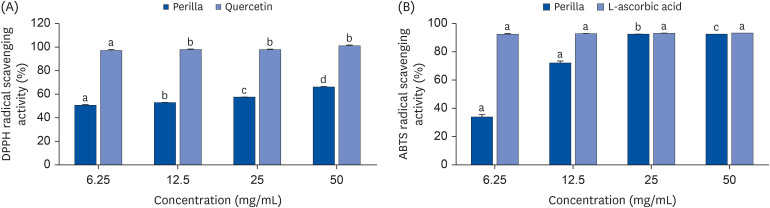 | Fig. 2Radical Scavenging activity of PE. (A) DPPH radical scavenging activities of PE and quercetin at various concentrations (6.25, 12.5, 25 and 50 mg/mL). The bar graphs of respective concentrations with different upper scripts differs significantly from each other when compared based on ANOVA Duncan’s multiple comparison test. Data is represented as mean ± SE. Each sample carried out in triplicates. (B) ABTS+ radical scavenging activities of PE and ascorbic acid at various concentrations (6.25, 12.5, 25 and 50 mg/mL). The bar graphs of respective concentrations with different upper scripts differs significantly from each other when compared based on ANOVA Duncan’s multiple comparison test. Data is represented as mean ± SE. Each sample carried out in triplicates.
|
Assay of ABTS•+ scavenging activity of PE
The ABTS•+ scavenging activity in percentage of PE was compared to a well- known antioxidant, L-ascorbic acid [27]. The ABTS•+ scavenging activity was dose-dependently reduced. PE gave results with scavenging values ranging from 92.49% to 33.91% for the first three highest concentrations; 50, 25 and 12.5 mg/mL (Fig. 2B).
Effect of PE on body weight, liver weight, and feed intake
For body weight changes (Table 3), our findings revealed significant reduction in the NDcon group (29.13 ± 0.96 g) and HDPE group (54.67 ± 4.9 g) compared with the HDcon group (61.07 ± 1.03 g). There was no significant difference in feed intake between groups. The liver weight was significantly reduced in the NDcon (2.78 ± 0.27 g) and HDPE group (1.12 ± 0.04 g) compared with the HDcon group (1.72 ± 0.24 g).
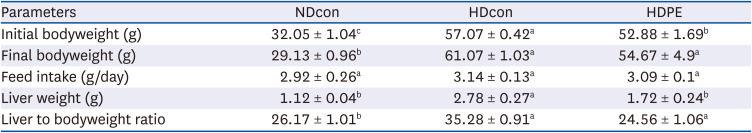
Table 3
Effect of PE on anthropometric parameters: body and tissue weight.
The experimental details are as described in the text. Results are given as mean ± SE (n = 8).
NDcon, normal diet fed reference group; HDcon, high-fat diet fed group; HDPE, high-fat diet PE treated group (400 mg/kg of body weight).
Alphabet upper scripts differ significantly from each other when compared based on ANOVA Duncan’s multiple comparison test. P < 0.05.
![]()
Effect of PE on fasting blood glucose and serum TNF-α
To test whether PE could improve glucose tolerance, we measured fasting blood glucose levels. Compared with the NDcon group (93.25 ± 3.15 mg/dL), HDcon group had significantly higher fasting blood-glucose levels (135.75 ± 4.77 mg/dL). However, supplementation of PE significantly reduced fasting blood-glucose concentration in the HDPE group (109.25 ± 4.52 mg/dL) (Table 4). In addition, serum TNF-α levels in the NDcon (52.20 ± 1.54) and HDPE group (51.60 ± 1.05 pg/mL) was significantly lower than HDcon group (61.90 ± 4.81 pg/mL) (Table 4).
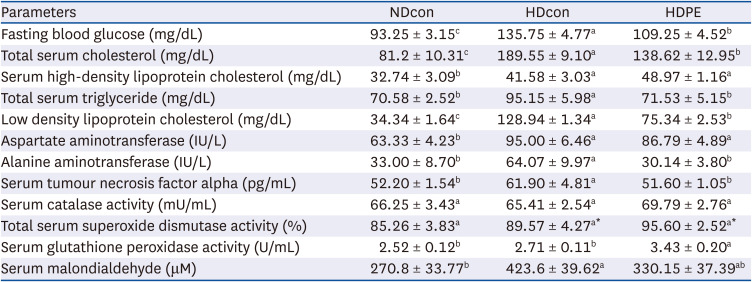
Table 4
Effect of PE on serum chemistry and lipid profiles
The experimental details are as described in the text. Results are given as mean ± SE (n = 8).
NDcon, normal diet fed reference group; HDcon, high-fat diet fed group; HDPE, high-fat diet PE treated group (400 mg/kg of body weight).
Alphabet upper scripts differ significantly from each other when compared based on ANOVA Duncan’s multiple comparison test. P < 0.05.
![]()
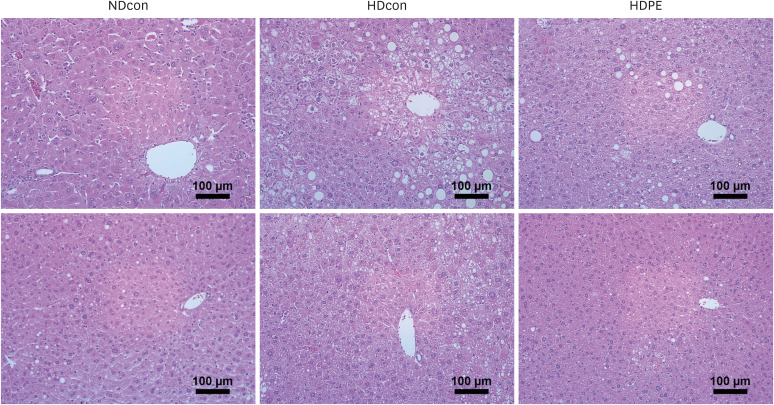
Effect of PE on hepatic histology, serum and liver lipid profiles
H&E staining of liver tissues in the HDcon groups revealed lipid accumulation and ballooning of hepatocytes, indicating the development of steatosis (Fig. 3). However, treatment with PE showed a noticeably reduced steatosis and resulted in a morphology similar to that of NDcon group.
Analysis of serum lipid profiles revealed that PE significantly reduced TG and TC levels compared with the HDcon group (P < 0.05). Serum TG and TC levels in the NDcon group (70.58 ± 2.52 and 81.2 ± 10.31 mg/dL) and HDPE group (71.53 ± 5.15 and 138.62 ± 12.95 mg/dL) was significantly lower than HDcon group (95.15 ± 5.98 and 189.55 ± 9.10 mg/dL). In addition, serum HDL cholesterol levels were slightly increased in HDPE group (48.97 ± 1.16 mg/dL) compared to HDcon (41.58 ± 3.03 mg/dL) groups, while those were significantly higher than NDcon (32.74 ± 3.09 mg/dL).
As the liver plays an important role in regulating lipid metabolism, hepatic TG and TC levels and histopathology were analyzed. Hepatic TG and TC levels of the HDPE group decreased by 19.8% and 18.4% respectively compared with HDcon group. Hepatic TG level of HDcon group (56.36 ± 3.45 mg/g) was significantly higher than NDcon (36.06 ± 6.99 mg/g) and HDPE (45.19 ± 7.83 mg/g) groups. Also, PE supplementation (24.63 ± 3.87 mg/g) significantly decreased the total hepatic TC levels compared to HDcon group (30.17 ± 2.81 mg/g) (Fig. 4).
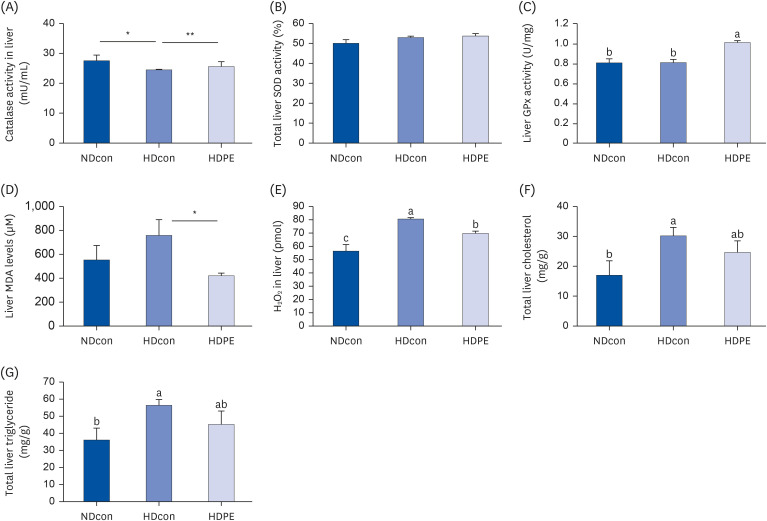 | Fig. 4Effect of PE on liver lipid profile and oxidative stress parameters (A) and (B) liver catalase and SOD show an increase pattern in liver tissue. (C) shows a significant increase in GPx levels in liver. (D) and (E) show significant decrease in oxidative stress products MDA and hydrogen peroxide levels in liver tissue of HDPE group compared to HDcon group. (F) and (G) reports a significant decrease in total liver cholesterol and total liver triglycerides in the liver of HDPE compared to HDcon. Each bar represents mean ± SE. ANOVA followed by independent t-test were used to find out statistical significance at P < 0.001. Bars with different upper scripts differs significantly from each other and (*,**) indicates significant difference according to independent t-test P < 0.05 (n = 8).NDcon, normal diet fed reference group; HDcon, high-fat diet fed group; HDPE, high-fat diet PE treated group (400 mg/kg of body weight).
|
Effect of PE on oxidative stress markers in serum and liver
The activities of the antioxidant enzymes SOD, CAT and GPx as well as MDA and H2O2 levels in serum and liver tissues are shown in Table 4 and Fig. 4 respectively. Significantly increased activity in the antioxidant enzymes SOD, CAT and hepatic GPx were observed in liver and serum in the NDcon and HDPE group when compared with HDcon mice. MDA levels in serum and liver showed significant reductions in the HDPE group compared with HDcon mice, falling from 423.6 ± 39.62 to 330.15 ± 37.39 µM and from 758 ± 131.75 to 419.75 ± 22.61 µM respectively. The liver H2O2 levels in the NDcon and HDPE group (56.33 ± 5.01 and 69.74 ± 1.70 pmoL) were significantly reduced in the comparison with the HDcon group (80.55 ± 1.05 pmoL).
Effect of PE on mRNA expression in oxidative stress related proteins and inflammatory cytokines in liver
To determine the oxidative-stress states of the control and PE supplemented groups, we measured the mRNA expression levels of oxidative stress and inflammation–related genes and enzymes. The mRNA expression levels of CAT, Nrf2, Peroxiredoxin (prdx), and NAD(P)H Quinone Dehydrogenase 1 (NQO1) in the HDPE group were significantly higher (P < 0.05) compared with the HDcon group, with increases of 87%, 63%, 40%, and 154%, respectively. In addition, hepatic inflammatory markers, such as COX-2, NOS, interleukin (IL)-1β, showed significant decrease in the HDPE group compared with the HDcon group. IL-6 reported a significant decrease as well from 0.16 to 0.015. On the other hand, anti-inflammatory IL-10 and IL-4 were increased significantly in the HDPE group compared with HDcon group (Fig. 5).
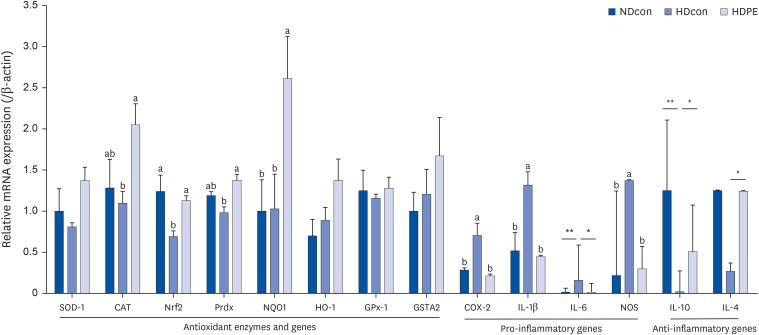 | Fig. 5Effect of PE on hepatic gene expression. mRNA expression levels in liver tissue. Bar graphs depicting the mean ± SE mRNA expression levels of antioxidant enzymes and genes, pro-inflammatory and anti-inflammatory genes in liver tissue of male CB57J6 mice. Mice divided into three groups; NDcon mice fed regular normal diet, HDcon mice fed high-fat diet and HDPE mice fed high-fat diet with daily oral gavage of PE 400 mg/kg bodyweight (n = 8). Multiple group comparisons were done by one-way ANOVA with a Duncan’s posthoc test. The two HD groups were compared against each other and NDcon. A significant increase in the expression of CAT, Nrf2 and some of its downstream genes and also a substantial non-significant increase in GSTA2 was observed between HDcon and HDPE mice. *,** indicates significant difference according to independent t-test P < 0.05.SOD1, superoxide dismutase 1; CAT, catalase; Nrf2, nuclear factor erythroid 2-related factor 2; Prdx, peroxiredoxin; NQO1, NAD(P)H Quinone Dehydrogenase 1; HO-1, heme oxygenase 1; GPx1, glutathione peroxidase 1; GSTA2, glutathione S-transferase alpha 2; COX-2, cyclooxygenase-2; IL-1β, interleukin 1 beta; IL-6, interleukin 6; NOS, nitric oxide synthase; IL-10, interleukin 10; IL-4, interleukin 4.
|
Go to :
DISCUSSION
The usage of herbs, plants, and nutraceuticals has been applied for the management of various diseases such as respiratory conditions, hypertension and obesity [2829]. A previous study investigated the phenolic and flavonoid contents in perilla leaves extracted with 60–100% ethanol.; and this study showed that all extractions contain various antioxidant compounds, but the amounts of each antioxidant compounds were differed based on ethanol concentrations [30]. Although the amounts of antioxidant compounds in our PE extracted samples were not measured, we presumed that our samples contain similar concentrations of antioxidant compounds with 70% ethanol extracted samples in previous study [30]. Our findings showed PE possessed antioxidant free-radical-scavenging capacity. This result is consistent with previous studies [31] indicating that PE possess high levels of metabolite content that have potential antioxidant activities against DPPH and ABTS radicals. Thus, our observations suggest the potentials of PE as an antioxidant and a radical scavenger in practical applications.
The effects of antioxidant PE [17] on age and HFD induced oxidative stress in aging obese mice were examined in this study. Our study administered PE to mice at a dose of 400 mg/kg body weight which according to Nair and Jacob [32] is equivalent to 1,946 mg per day for a 60 kg typical body weight in human dosage. Previous clinical investigations on the protective effects of a variety of natural products against obesity and its comorbidities at that concentration have reported no adverse effects [33]. In our findings, PE significantly reduced serum ALT and AST levels, which are common biomarkers for liver toxicity and pathology [34]. According to the result, the mice exhibited no adverse effects to PE supplementation at the administered dosage.
In this study we hypothesized that advanced age in combination with long-term consumption of HFD will induce chronic oxidative stress and depleted antioxidant defenses, leading to the damages in cells and tissues. Accumulation of oxidative stress promotes several pathophysiological events in the liver, including detrimental influences on hepatocytes, DNA, lipids, and proteins, and ultimately disruption of cellular homeostasis [3536]. In this study, H&E staining of liver tissue showed that HFD caused disordered lobule structure, micro-steatosis, and excessive lipid droplet accumulation, whereas PE supplementation improved liver steatosis and pathology, indicating that PE may be able to ameliorate liver damage by long-term HFD intake. Excessive TG accumulation in liver tissues and cells impairs cellular antioxidative capacity and results in overproduction of ROS and hepatic oxidative stress injury directly by attacking lipids, proteins, and nucleic acids [37]. Our findings reported the HFD elevated hepatic and serum TG, TC and LDL levels and glucose tolerance, while PE supplementation significantly attenuated these elevations. Thus, our results demonstrated that PE ameliorates hyperlipidiema and glucose tolerance in HFD-induced obese in aged mice. Our findings support that the previous study that showed PE lowered TG, TC, and low-density lipoprotein-cholesterol levels exhibiting hypolipidemic activity [38].
ROS can also indirectly activate several pro-inflammatory signaling pathways such as the TNF-α pathway to induce cellular injury. Previous studies have reported that dysregulation of pro inflammatory cytokines and chemokines such as TNF-α and IL-6 are significant culprits in the development of chronic inflammation and the immunosenescence process. The immunosenescence process increases the levels of TNF-α and IL-6 and their receptor expression in aged cells [39]. According to a previous study [40] aging and oxidative factors combined with long term HFD consumption increased synthesis of endotoxins such as lipopolysaccharide, increasing the production of proinflammatory cytokines. Consistent with previous findings [41], we found that PE supplementation significantly lowered serum TNF-α levels. We also discovered that pro-inflammatory hepatic IL-1β and IL-6 gene expression levels significantly decreased, while anti-inflammatory IL-10 and IL-4 hepatic gene expression were significantly increased. Another pathway altered by oxidative stress is the redox signaling pathways which also causes activation of pro-inflammatory genes like COX-2 and NOS [42]. COX-2 overexpression plays a role in the aging process and development of a number of age-related illnesses [43]. In chronic inflammatory conditions, NOS produces large quantities of harmful NO. Our study proved that hepatic COX-2 and NOS gene expressions were significantly reduced by PE administration, suggesting anti-inflammatory capacities of PE.
SOD, CAT and GPx enzyme activities increased by PE supplementations in HFD-induced obese in aged mice. We hypothesized that these increased activities attenuate the accumulation of reactive free radicals based on the previous studies [44]. In addition, reduction of harmful oxidative end products such as MDA and H2O2 presents an alternative mechanism for ameliorating oxidative stress damage [45]. Our results showed that MDA and H2O2 levels in the HDPE group were lower in comparison with the HD control group. From our findings in this study, we hypothesized that the increased antioxidant enzymes SOD, CAT and GPx levels decreasethe accumulation of free-radicals. To confirm our results and possible mechanism pathway, we focused on Nrf2 signalling pathway, which plays a well-established and vital role in protection against antioxidant and oxidative stress damage [464748]. In our study, hepatic mRNA expression of Nrf2, CAT and peroxiredoxin (Prdx) in HDPE was significantly upregulated by PE supplementation compared to the HDcon group. While PE supplementation did not significantly affect the expression of hepatic SOD-1, HO-1, gpx-1 and GSTA2, all four were increased compared to the HDcon group. Inhibition of the activities of antioxidant enzymes such as SOD, GPx plays a role in the development of oxidative stress. However, their activities are enhanced after supplementation of an antioxidant [49]. Our findings suggest that PE is an antioxidant that protects against oxidative damage by scavenging ROS and upregulating of related enzyme expressions.
There are two major limitations in this study that could be addressed in future research. First, the study focused on diet related control groups, lacking an age related control group that would have been evidence on the actual effect of perilla against oxidative stress on aging alone. Second, this study did not investigate the possible involvements of nonenzymatic antioxidants such as reduced glutathione (GSH). The oxidative stress of tissue generally involves the GSH system. However, previous studies [5051] reported that PE increased GSH levels in vivo and in vitro against tert-butyl hydroperoxide-induced oxidative hepatotoxicity, suggesting that PE could also improve non-enzymatic antioxidants system in HFD-induced obese in aged mice
In conclusion, perilla supplementation increased antioxidant enzyme expression, reduced inflammatory markers, ROS levels and oxidative stress products and damage associated with aging and HFD. Thus, our study provides the preclinical evidences that green perilla has an anti-aging, anti-inflammatory and anti-oxidative-stress effects against oxidative stress related damages resulting from aging and HFD. However, future studies should incorporate various doses and target models to confirm the effects of perilla as a functional food or nutraceutical product for the treatment of oxidative stress.
Go to :
 XML Download
XML Download