

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a chronic, relapsing, and highly itching inflammatory skin disease [12]. In Europe and the Unites States, approximately 20% of children and 7% to 14% of adults suffer from AD [3]; the prevalence, unfortunately, is increasing. Although, the pathogenesis of AD has been intensively studied in recent decades, the optimal approaches to manage AD have not been well established and corticosteroids are still the main stream treatment for AD [45]. However, the long-term use of corticosteroids might lead to irreversible skin atrophy and perturb normal biological activities [5]. Recently, immunosuppressive drugs including tacrolimus and pimecrolimus, have been used for the treatment of AD [67]. The use of tacrolimus to treat moderate to severe AD and pimecrolimus to treat mild to moderate AD was shown to be effective and safe [8]. However, these agents showed poor absorption into the skin and a burning sensation especially during the application of tacrolimus [9]. Therefore, exploring a safer and more efficient therapy is still of great value and is experiencing an upsurge in demand.
NC/Nga mice have been used as an AD model for human dermatitis since 1957 by Kondo et al. [10]. NC/Nga mice will become dry and scaly skin under conventional conditions [1011]. Skin lesions also developed immunologically and histologically around neck and back within several weeks. At the same time, serum IgE level moderately increased demonstrating a biochemical change [1012]. Elevated IgE levels are the key to the pathogenesis of AD, resulting in the activation of mast cells and leading to the release of mediators such as histamine [13141516]. These mediators were positively correlated with skin lesions like prurience and eczema [141516]. Thus, controlling the inflammation status in AD is thought to be important.
Pears, one of the oldest plants cultivated by humans, are consumed all over the world [17]. Yet, pear pomace, the main waste byproduct in pear processing, is hitherto poorly understood. Some studies have suggested that pear pomace showed beneficial effects in increasing fecal bulking, toxic component binding, and the regulation of gut microbiota structures in high-fat diet-fed rats due to the high dietary fiber content [18192021]. In our preliminary results, pear pomace water extract (PPW) inhibited adipocyte differentiation by inducing apoptosis in the early stages of differentiation and reduced hepatic lipid peroxidation [2223]. In addition, pear pomace ethanol extract (PPE) effectively reduced insulin resistance in vitro and in vivo [24]. We also found that PPE was more effective at inhibiting inflammation than PPW in RAW 264.7 cells after lipopolysaccharide (LPS) stimulation (data not shown). However, the anti-inflammatory effect of PPE, especially in AD, has not been examined.
In the present study, we hypothesized that PPE could prevent AD by regulating inflammatory responses in 2, 4-dinitrochlorobenzene (DNCB)-induced AD in NC/Nga mice and LPS-induced inflammation in RAW 264.7 macrophage. In addition, we measured whether nuclear factor kappa B (NF-κB) was involved in the action mechanism of PPE.
Go to : 
MATERIALS AND METHODS
Preparation of PPE
The ethanol extract of pear pomace was prepared according to previous protocols [24]. Briefly, pear pomace, the cake left after extracting the juice, was extracted with 50% ethanol, filtered, freeze-dried, and stored in a −80°C freezer. The yield obtained was 27.5%
Animals
Five-week-old male NC/Nga mice were purchased from Central Laboratory Animal Inc. (Seoul, Korea), and were housed individually in ventilated cages under conventional conditions with a 12 h light/dark cycle at 22 ± 2°C room temperature and 50 ± 10% relative humidity during the experimental period. Mice had free access to water and were fed an AIN-93G control diet. All experiments were approved by the Laboratory Animal Care and Use Committee of Mokpo National University (MNU-IACUC-2016-005).
Induction of AD using DNCB
DNCB was an efficient tool to induce AD in NC/Nga mice within a short time period topical application. We slightly modified the AD protocol according to published papers [252627]. After 2 week of acclimatization, the dorsal hair of NC/Nga mice was removed using an electric shaver (hair clipper model 6000AD; Thrive Japan, Kawasaki, Japan) and depilatory cream (Veet; Reckitt, Slough, UK). To induce AD-like skin lesions, the mice were divided into 2 groups (normal control [Cont] group and AD group); DNCB (Sigma-Aldrich, St. Louis, MO, USA) was applied to the AD group. The mice were sensitized with 200 uL of 1% DNCB solution (dissolved in a 3:1 mixture of acetone and olive oil) twice over a week. Then, the dorsal skin was challenged with 150 uL of 0.4% DNCB solution 3 times per week for 7 weeks. Three weeks after the challenge, the AD group was further divided into 3 groups: AD control (AD-C), AD + PPE 200 (PPE 200 mg/kg body weight), and AD + PPE 400 (PPE 400 mg/kg body weight). PPE extract was dissolved in distilled water and administered daily by oral gavage for 4 weeks. Distilled water was administered to the Cont and AD-C groups. Fig. 1A illustrates the schematic diagram of AD induction and treatment with PPE.
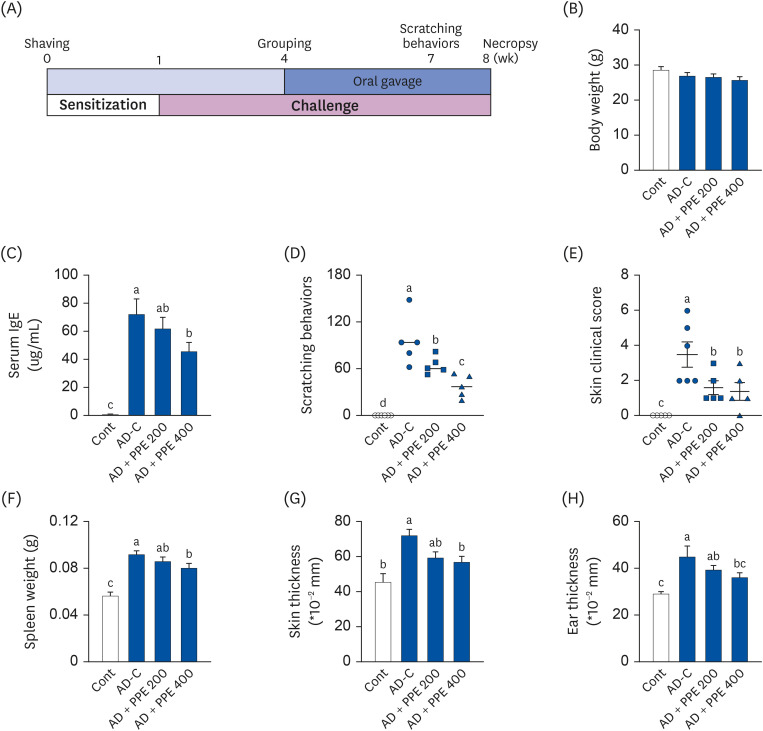 | Fig. 1Pear pomace ameliorated AD symptoms in DNCB-induced AD skin lesion in NC/Nga mice. (A) Experiment design. To induce AD-like lesions, DNCB was applied to NC/Nga mice (n = 7). The mice were sensitized with 1% DNCB for 1 week and then challenged with 0.4% DNCB solution for 7 weeks. The mice were administered PBS or PPE daily for 4 weeks. (B) Final body weight. (C) Serum levels of IgE. (D) Total scratching behavior scores 7 weeks after sensitization. (E) Clinical severity score for dermatitis on the necropsy day. (F) Spleen weight. (G) Skin thickness. (H) Ear thickness.AD, atopic dermatitis; DNCB, 2, 4-dinitrochlorobenzene; PBS, phosphate-buffed saline; PPE, pear pomace ethanol extract; Cont, no sensitization; AD-C, DNCB treatment with oral administration of PBS (AD control); AD + PPE 200, DNCB treatment with oral administration of PPE 200 mg/kg body weight; AD + PPE 400, DNCB treatment with oral administration of PPE 400 mg/kg body weight.
Different letters are significantly different by Duncan’s multiple range test (P < 0.05).
|
Measurement of basic characteristics in NC/Nga mice
To investigate the scratching behavior of DNCB-induced AD in NC/Nga mice, the mice were placed individually in clear cages. Seven weeks after sensitization, the mice were treated with PPE upon 0.4% DNCB treatment. For 20 min after DNCB treatment, the total number of mice scratching themselves only with the hind paw was counted; the scores ranged from 0 to 4: 0 (no scratching), 2 (scratching for less than 1.5 sec), and 4 (scratching for more than 1.5 sec). The clinical severity score for dermatitis was evaluated macroscopically on the day of the necropsy using a specific scoring procedure. The skin clinical severity score was expressed as the sum of the individual scores for the 4 symptoms of erythema/hemorrhage, dryness/scaling, edema, and excoriation/erosion. For each skin symptom, 0 was defined as no symptoms, 1 as mild symptom, 2 as moderate symptoms, and 3 as severe symptoms [2528]. To determine the ear thickness, both the left and right ears of the mice were removed, and the thickness was measured using a digital caliper (Mitutoyo Corporation, Kawasaki, Japan), and calculated as the mean thickness value of both sides. Additionally, the spleen was dissected for weighing.
Measurement of serum IgE levels in NC/Nga mice
Blood samples were collected, separated immediately by centrifugation, and stored at −70°C. Serum IgE levels were determined using the mouse IgE ELISA kit (Fujifilm Wako Shibayagi Corporation, Shibukawa, Japan) following the manufacturer’s instructions.
Histological analysis of NC/Nga mice
To measure the epidermal inflammation and infiltration of mast cells, dorsal skin was obtained and fixed in 10% formalin. The samples were embedded in paraffin, sectioned at 4 μm, and stained with hematoxylin and eosin (H&E) or toluidine blue (Sigma-Aldrich). The stained sections were observed under a microscope at 100× magnification.
Cell culture and viability
Murine cell line RAW 264.7 macrophages (American Type Culture Collection, Manassas, VA, USA) were cultured in Dulbecco’s modified Eagle’s medium (Gibco-BRL, Grand Island, NY, USA) which was supplemented with a 10% fetal bovine serum (Gibco-BRL) and 1% penicillin (Gibco-BRL). The cells were maintained at 37°C in a humidified atmosphere with 5% CO2. At 80% of confluence, the cells were subdivided for future experiments. To examine the cytotoxicity of PPE, the cells were seeded in a 96-well plate at a density of 10 × 103 cells/well and were subsequently treated with either phosphate-buffed saline (PBS), 100, or 250 ug/mL of PPE. After 24 h of incubation, 2 mg/mL of 3-(4,5-dimethylthiazol-2-yl)-2,5 diphenyl tetrazolium bromide (MTT) solution (Sigma-Aldrich) was added to each well and the cells were incubated further for another 4 h. The medium was then removed, and the formazan precipitate was solubilized in dimethyl sulfoxide (Sigma-Aldrich). Absorbance was measured at 540 nm using microplate reader (Multiskan EX; Thermo Fisher Scientific, Waltham, MA, USA). The relative cell viability was calculated as the percentage of the control cells.
Measurement of nitrite oxide (NO) production in RAW 246.7 macrophages
The nitrite concentration in the culture medium was measured as an indicator of NO production. RAW 264.7 macrophages were seeded at a density of 20 × 104 cells/well in a 24-well plate and pretreated with PBS or PPE for 1 h before stimulation with 1 ug/mL of LPS (Sigma-Aldrich). After 24 h incubation, the culture supernatant was collected and centrifuged at 2,400 g for 10 min. The cell culture medium was combined with an equal volume of Griess reagent (Sigma-Aldrich) in a 96-well plate and incubated for 15 min at room temperature. Then, the absorbance was measured at 540 nm using microplate reader.
Western blot analysis
Mouse skin samples and cells were homogenized in ice-cold radioimmune precipitation assay buffer (Thermo Fisher Scientific) with a 1% protease inhibitor cocktail and a phosphatase inhibitor. The protein concentrations were determined by the Bradford (Sigma-Aldrich) method and equal amounts of protein (40 μg each) were separated on 10% SDS-PAGE gels and transferred onto a nitrocellulose membrane (Millipore, Billerica, MA, USA). The membrane was blocked for 1 h at 4°C with 5% skim milk in TBS-T buffer (10 mM Tris-HCl [pH 6.8], 100 mM NaCl, and 0.1% Tween 20). The membranes were incubated with primary antibodies and then with horseradish peroxidase-conjugated secondary antibody (goat anti-rabbit or goat anti-mouse [1:1,000]) Antibodies targeting receptor for advanced glycation end products (RAGE; #6996), p-extracellular signal-regulated kinase (ERK) 1/2 (#4370), ERK1/2 (#4695), NF-κB p65 (#8242), inducible nitrite oxide synthase (iNOS; #2982), and cyclooxygenase (COX)-2 (#12282) were purchased from Cell Signaling Technology (Beverly, MA, USA). B-actin antibody (#47778) was purchased from Santa Cruz Biotechnology (Dallas, TX, USA). Bands were detected using chemiluminescent substrate (IMGENEX, San Diego, CA, USA), and protein expression was quantified using UVP (imaging system for the chemiluminescent Western blot; UVP, Upland, CA, USA) and Vision Works TMLS (analysis software; UVP).
Statistical analysis
Statistical analyses were performed using the SPSS program (version SPSS 23; IBM, Chicago, IL, USA). The results are presented as the mean ± SE of 3 independent experiments. Differences between the groups were evaluated using one-way analysis of variance. Multiple comparisons were performed by Duncan’s multiple range test. A P-value of < 0.05 was considered statistically significant.
Go to :
RESULTS
Effect of PPE on the basic characteristics of NC/Nga mice
To induce AD-like skin lesions, NC/Nga mice were sensitized with a high dose of DNCB solution for 1 week. After sensitization, a low dose of DNCB was applied on the dorsal skin of the mice with or without PPE treatment (200 or 400 mg/kg body weight) (Fig. 1A). As shown in Fig. 1B, no significant differences in body weight were observed among the groups at the end of the experimental period, indicating that PPE could be used for in vivo applications safely with no visual side effects. Importantly, serum IgE levels in the AD-C group were notably higher by 76-fold (72.55 ug/mL) compared to 0.94 ug/mL in the Cont group. The high-dose PPE treatment reduced serum IgE levels to about 37.2% (45.53 ug/mL) compared to the AD-C group (Fig. 1C). Investigations were carried out to evaluate the clinical symptoms, scratching behaviors and skin clinic severity score. The application of DNCB remarkably increased scratching behavior; PPE at 200 and 400 mg/kg decreased scratching behaviors by 23.6% and 40%, respectively, versus the AD-C group (Fig. 1D). Additionally, PPE treatment markedly improved skin clinical severity score relative to the AD-C group (Fig. 1E). In previous studies by Ko et al. [29] and Shin et al. [30], NC/Nga mice treated with DNCB showed an enlarged spleen, the organ responsible for maintaining systemic immune responses. Spleen weight in the AD-C group markedly increased compared to that in the Cont group, whereas PPE treatment lowered spleen weight (Fig. 1F). Furthermore, the thickness of the skin and ear was also suppressed by PPE treatment (Fig. 1G and H).
PPE reduced the aggravation of AD-like skin lesions in DNCB-induced NC/Nga mice
To investigate the histological changes in the dorsal skin of NC/Nga mice, skin samples were stained with H&E and toluidine blue. A thicker epidermis was observed in the dorsal skin of AD-C specimens compared to specimens from the Cont group, whereas specimens from the PPE groups exhibited decreases in epidermal thickness (Fig. 2A). The regulation of mast cells in the allergic response is an essential strategy in autoimmune diseases such as AD [31]. The AD-C group showed significantly higher numbers of mast cells in the dorsal skin than the Cont group. Nevertheless, the infiltration of mast cells diminished with PPE supplementation in AD-induced NC/Nga mice (Fig. 2B). Taken together, these results indicated that the administration of PPE attenuated AD-like lesions in DNCB-induced NC/Nga mice.
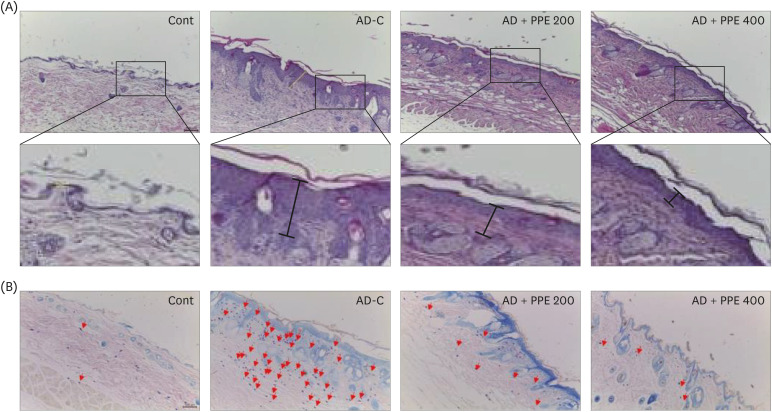 | Fig. 2Histological changes in the dorsal skin in pear pomace-treated NC/Nga mice. (A) H&E staining in skin tissue. (B) Toluidine blue staining. The arrows indicate mast cells in the skin tissue.AD, atopic dermatitis; DNCB, 2, 4-dinitrochlorobenzene; H&E, hematoxylin and eosin; PPE, pear pomace ethanol extract; Cont, no sensitization; AD-C, DNCB treatment with the oral administration of PBS (AD control); AD + PPE 200, DNCB treatment with the oral administration of PPE 200 mg/kg body weight; AD + PPE 400, DNCB treatment with oral administration of PPE 400 mg/kg body weight.
|
PPE inhibited inflammation markers in the skin of AD-induced NC/Nga mice
Next, we detected inflammation-related protein expression in the dorsal skin, as shown in Fig. 3. The protein levels of RAGE, a marker of inflammation, were elevated upon DNCB treatment, whereas PPE reversed those changes in the dorsal skin of AD induced NC/Nga mice. The application of DNCB drastically increased the p-ERK/ERK ratio relative to the Cont group. However, PPE dose-dependently reduced the p-ERK/ERK ratio compared to the AD-C group; moreover, PPE 400 effectively returned the p-ERK/ERK levels to those of the Cont level. In accordance with the p-ERK/ERK results, the protein expression of NF-κB showed a significant drop in the dorsal skin of the PPE groups (Fig. 3). Compared to the elevation in iNOS protein in the AD-C group, PPE treatment considerably suppressed iNOS expression (Fig. 3). Taken together, these results indicated that PPE treatment could help to prevent DNCB-induced AD in NC/Nga mice.
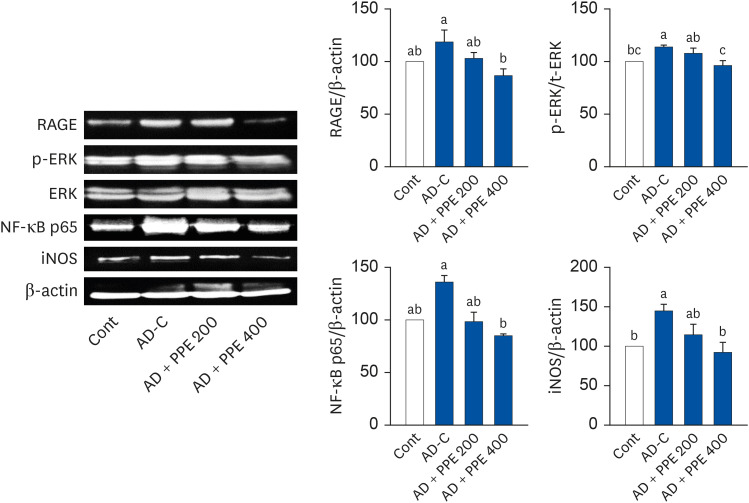 | Fig. 3Applications of pear pomace inhibited inflammation-related proteins in the skin tissue of NC/Nga mice. β-actin was used as the control for quantification.Cont, no sensitization; AD-C, DNCB treatment with oral administration of PBS (AD control); AD + PPE 200, DNCB treatment with the oral administration of PPE 200 mg/kg body weight; AD + PPE 400, DNCB treatment with the oral administration of PPE 400 mg/kg body weight; RAGE, receptor for advanced glycation end products; ERK, extracellular signal-regulated kinase, NF-κB, nuclear factor kappa B; iNOS, inducible nitrite oxide synthase; PPE, pear pomace ethanol extract.
Different letters are significantly different by Duncan’s multiple range test (P < 0.05).
|
PPE suppressed inflammatory response in LPS-induced RAW 264.7 macrophages
Due to the anti-inflammation effect of PPE in vivo, we studied whether PPE could inhibit the inflammatory response upon LPS treatment in the RAW 264.7 macrophages. We first measured cell cytotoxicity in the RAW 264.7 cells using MTT assay. The cell viability was not different among the groups. To determine the inhibitory effect of PPE on NO production, RAW 264.7 cells were pre-treated with PPE. Stimulation with LPS remarkably aggravated NO production compared to the control; pretreatment of PPE ameliorated these increases in a dose-dependent manner (Fig. 4B). The elevated NO production in LPS treated cells was consistent with the increased expression of iNOS and COX-2 proteins (Fig. 4C). Conversely, treatment with a high dose of PPE dramatically reduced iNOS and COX-2 protein expression compared to the LPS control in RAW 264.7 macrophages. Consistent with the in vivo study results, LPS significantly raised the p-ERK/ERK ratio (Fig. 4D) and PPE remarkably reduced the p-ERK/ERK ratio in LPS-stimulated cells (Fig. 4D). Furthermore, the expression of NF-κB/p65 was decreased in the PPE treated groups. These results collectively suggested that PPE could potently suppress the inflammatory response in RAW 264.7 macrophages.
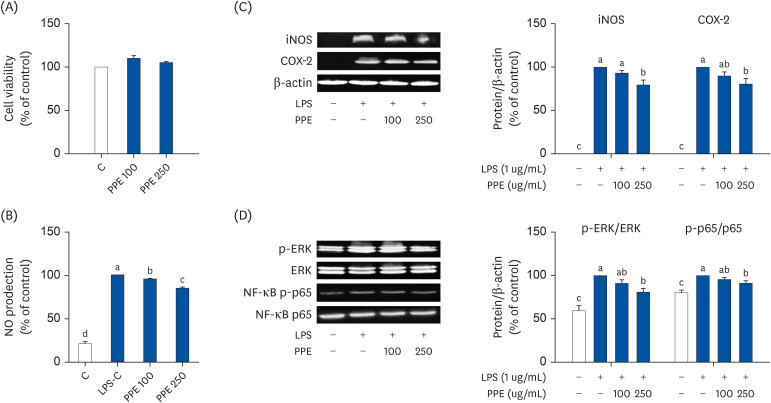 | Fig. 4Inhibitory effects of pear pomace on the inflammatory response in LPS-induced inflammation in RAW 264.7 macrophages. RAW 264.7 cells were pretreated with either 0, 100, or 250 ug/mL of PPE for 1 h before the addition of 1 ug/mL of LPS treatment for 24 h. (A) Cell viability. (B) NO production. (C) Protein expression of iNOS and COX-2. (D) Protein expression of ERK and NF-κB. β-actin was used as the control for quantification.LPS, lipopolysaccharide; NO, nitrite oxide; iNOS, inducible nitrite oxide synthase; COX-2, cyclooxygenase-2; ERK, extracellular signal-regulated kinase; NF-κB, nuclear factor kappa B; PPE, pear pomace ethanol extract.
Different letters are significantly different by Duncan’s multiple range test (P < 0.05).
|
Go to :
DISCUSSION
In the present study, we examined whether PPE alleviated the clinical features of AD using AD-like lesions in DNCB-induced NC/Nga mice and LPS-induced inflammation in RAW 264.7 macrophages. We found that the administration of PPE inhibited AD-like lesions (Figs. 1 and 2) and suppressed inflammation-related proteins in the dorsal skin of AD-induced NC/Nga mice (Fig. 3). We also identified that PPE decreased NO production and NF-κB expression in the LPS-induced inflammatory response in RAW 264.7 cells (Fig. 4). Taken together, our work indicates that PPE may be developed as a therapeutic agent to ameliorate AD.
AD is a chronic inflammatory skin disease characterized by the hyper production of IgE. Seventy percent of AD patients showed elevated levels of IgE which was positively associated with disease severity. The patients with low IgE level slowly displayed allergic reactions due to IgE-mediated sensitization later in life [32]. The IgE level is considered as an authentic prognostic indicator of AD and a key element in the pathogenesis of AD [14]. During the allergic reaction, IgE is increased and binds to mast cells with high affinity. IgE-activated mast cells release mediators including histamine, prostaglandins, and leukotrienes. In addition, the over-secretion from mast cells results in the infiltration of inflammatory epidermal cells, macrophages, and eosinophils [151628]. Recently, Emrick et al. [33] demonstrated that the Tmem79 gene, a newly identified gene in AD animals and humans, played a key role in skin inflammation and itching [3435]. The mutation of Tmem79 in mice showed the dysregulation of inflammation, dermal thickness, and skin lesions. Interestingly, blocking mast cell degranulation decreased scratching behaviors in Tmem79−/− mice by inhibiting histaminergic itching via the histamine receptor 1 and 4-dependent pathways [33]. These findings suggest that the reduction in mast cells may be related to the decrease in scratching behaviors. In our study, PPE remarkably reduced IgE levels and mast cell infiltration (Figs. 1C and 2B). In addition, PPE effectively inhibited scratching behavior and improved clinical symptoms (Fig. 1D and E). Increased epidermal thickness by DNCB treatment was suppressed in the PPE-treated groups (Figs. 1G and 2A). These protective effects against the AD symptom suggest that PPE treatment may be useful for the treatment or management of AD.
RAGE is a multiligand receptor in the immunoglobulin superfamily on the cell surface and has been implicated as an amplifier of pro-inflammatory responses in inflammatory diseases [3637]. The activation of RAGE engages key signaling pathways including ERK1/2 phosphorylation and the downstream NF-κB pathway to mediate the expression of molecules that are involved in inflammation [36]. NF-κB, composed of the p50/p65 heterodimer and the inhibitory protein IkBα, is a ubiquitous transcription factor that resides in the cytoplasm of resting cell. Once activated, phosphorylated IκBα underwent degradation and further disassociation. The remaining p50/p65 heterodimer translocate to the nucleus, where it induces the transcription of more than 400 genes of which are associated with cellular activities such as apoptosis, proliferation, metastasis, chemo resistance, and inflammation. [3839]. Therefore, the regulation of mediators via the NF-κB pathway is a critical mechanism in the inflammatory process and a potential target for inflammation intervention. Consistent with the literature, our results indicated that PPE treatment reduced the expression of RAGE that was increased by the application of DNCB (Fig. 3). In addition, PPE treatment inhibited NF-κB expressions in the dorsal skin of NC/Nga mice (Fig. 3) and in RAW 264.7 cells (Fig. 4). Moreover, iNOS, one of the types of NOSs, is attributed to the redundant or overproduced NO during inflammation or infection [40]. Our present findings clearly displayed that iNOS expression was effectively diminished through PPE treatment both in vivo and in vitro (Figs. 3 and 4C). The down-regulated iNOS expression may, in part, explain the PPE-decreased NO production in RAW 264.7 cells (Fig. 4B). Since, when NF-κB is activated, the p65/p50 heterodimer disassociates with the phosphorylation of IκBα and translocate from the cytosol to the nucleus, PPE may block the translocation of NF-κB to the nucleus and thus, down-regulate iNOS expression. However, whether PPE also interferes with the phosphorylation of IκBα remains unknown and further studies are needed.
The main active ingredients in pears are considered to be phenolic and flavonoids compounds, among which, leucocyanidin, catechin, epicatechin, chlorogenic acid, arbutin, ursolic acid, quercitrin, and quercetin account for a large proportion [41]. The unique combination of polyphenols and flavonoids may be related to the potential activity for treating allergic inflammatory diseases [33]. In addition, remarkable progress has been achieved in recent years to elucidate the anti-AD properties of quercetin [4243]. Moreover, chlorogenic acid has been widely studied as an antioxidant in mouse RAW 264.7 macrophages [4445] and human THP-1 macrophages [46] which can mitigate inflammatory response. However, since there has been no experimental report on the screening of the bioactive fractions and components from PPE, an arbitrary decision about which component is mainly responsible for the anti-inflammatory properties could not be made, and further research is required to clarify those issues.
In conclusion, we demonstrated that PPE was effective in decreasing AD-like lesions in DNCB-induced NC/Nga mice. The inflammatory response including NF-κB expression in the dorsal skin was inhibited by PPE treatment. PPE was also shown to ameliorate LPS-induced inflammation in RAW 264.7 cells by decreasing NO production and NF-κB expression. Taken together, PPE might represent an anti-inflammatory candidate to treat inflammatory diseases like AD.
Go to :
 XML Download
XML Download