

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
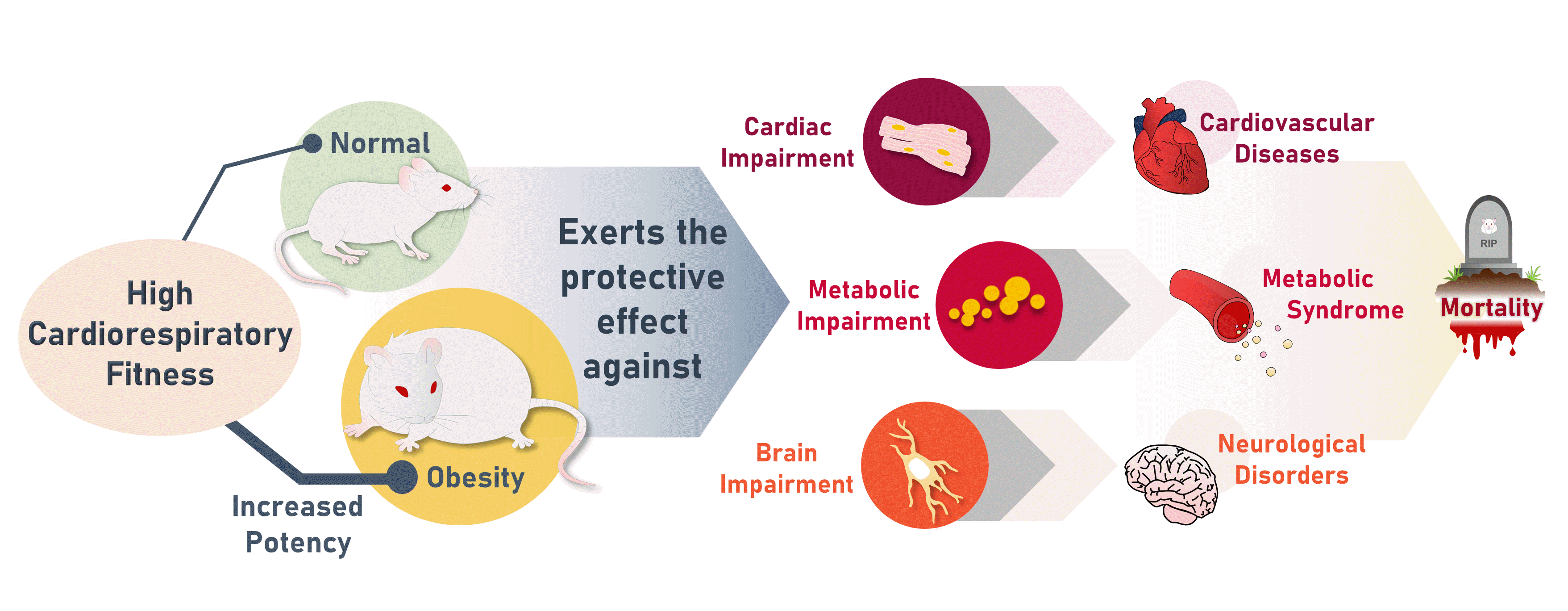
Cardiorespiratory fitness (CRF) represents the ability of the cardiovascular, respiratory, and muscular systems to supply oxygen during exercise [1]. It has been demonstrated that CRF is significantly genetically determined, with a heritability of 72% [2]. Previous longitudinal studies have shown that people with higher CRF levels had lower risks of metabolic syndrome, cardiovascular diseases, dementia, and all-cause mortality [3-5], suggesting that high CRF potentially protects against metabolic, cardiac, and brain impairments.
Previous studies investigating the mechanisms responsible for high CRF-induced delayed metabolic impairment were established using rats selectively bred for high and low intrinsic running capacity (HCR and LCR) [6]. It was observed that HCR rats had lower adiposity, higher insulin sensitivity, and a more favorable lipid profile [6-9]. Moreover, HCR rats had greater energy expenditure and heat production in skeletal muscle during exercise [10]. Metabolomic and proteomic analyses also revealed that skeletal muscle from HCR rats exhibited a higher capacity for utilization of fatty acids and branched-chain amino acids [11]. These findings may all be mechanistically linked to the protective effect of high CRF against metabolic syndrome. However, the effect of CRF has never been compared between non-obese and obese conditions, although obesity is known to accelerate aging [12]. The protective role of CRF in non-selectively bred animals remains unknown. The hearts of HCR rats have been shown to be characterized by higher respiration, greater antioxidative capacity, a lower degree of remodeling, and more favorable systolic and diastolic function [13,14]. Regarding the brain, HCR rats displayed a higher neuronal number, a greater hippocampal volume, a higher mitochondrial DNA number, higher respiration, lower expression of Alzheimer’s disease (AD)-related proteins, and better cognition [15,16]. The above findings imply that some molecular mechanisms potentially underlying the effect of high CRF on protection against cardiac and brain impairments have not yet been determined. As is the case with metabolic impairment, this effect of CRF has been neither compared between non-obese conditions and obesity-induced premature aging, nor evaluated in non-selectively bred animals.
Therefore, the aim of this study was to investigate the mechanisms mediating the effect of high intrinsic CRF on protection against metabolic, cardiac, and neuronal dysfunction in non-obese versus obese untrained Wistar rats. We hypothesized that a high intrinsic CRF level would protect against metabolic, cardiac, and neuronal impairment by modulating several mechanisms, and importantly, that these effects of high intrinsic CRF would also be observed in obesity-induced premature aging.
METHODS
Animals
The experiments were performed at the laboratory animal center of Chiang Mai University, Thailand following a protocol approved by the Animal Care and Use Committee of Chiang Mai University, Chiang Mai, Thailand (2563/RT-0006). Male Wistar rats (n=16) were purchased from Nomura Siam International Co. Ltd. (Bangkok, Thailand).
Study protocol
The study protocol is illustrated in Supplemental Fig. S1. Male Wistar rats at the age of 7 weeks (n=16) were randomly assigned into two equal groups (n=8 per group). The first group received an ad libitum normal diet (ND) containing 19.7% energy from fat. The second group received an ad libitum high-fat diet (HFD) containing 59.28% energy from fat. The HFD was made by the laboratory animal center of Chiang Mai University, Chiang Mai, Thailand. At weeks 12 and 28, CRF and whole-body substrate oxidation during exercise were evaluated. Cardiac function was also investigated at weeks 12 and 28 using echocardiography and heart rate variability (HRV). Additionally, blood was collected by cutting the tip of the tail after 5 hours of fasting at weeks 12 and 28 to measure metabolic parameters, including the insulin sensitivity profile and lipid profile. At week 28, behavior tests including an open-field test (OFT) and modified novel object location (NOL) and novel object recognition (NOR) tests for cognitive function were performed. At the end of week 28, all rats were euthanized to enable the collection of heart and brain tissues for molecular studies. Regular insulin (10 U/kg; Actrapid HM, Novo Nordisk, Copenhagen, Denmark) was intraperitoneally injected into the rats 30 minutes before decapitation.
General methods
The details regarding the general methods are described in the Supplemental Methods. These methods include CRF level measurements, whole-body substrate oxidation during vigorous exercise, plasma insulin sensitivity and lipid profiles, echocardiography and HRV, behavior and cognitive tests (OFT, NOL, and NOR), measurement of malondialdehyde concentrations in cardiac and brain tissues, protein expression analyses, cardiac mitochondrial function and mitochondrial respiration assay, hippocampal reactive oxygen species levels, ex vivo brain incubation with regular insulin solution, and an apoptotic cell death assay.
Statistical analyses
For comparisons between the different time points (baseline at week 12 vs. follow-up at week 28) within the same group, data were analyzed using the paired Student’s t test. For comparisons between ND-versus HFD-fed rats, and between subgroups of rats with higher baseline CRF levels (ranked first to fourth in baseline running distance; n=4) versus those with lower baseline CRF levels (ranked fifth to eighth in baseline running distance; n=4) within each diet group, the unpaired two-tailed Student’s t test was performed. Correlations within each group of rats were determined using Pearson correlation coefficients. A P<0.05 was considered statistically significant.
RESULTS
Deterioration in metabolic and cardiac function over time
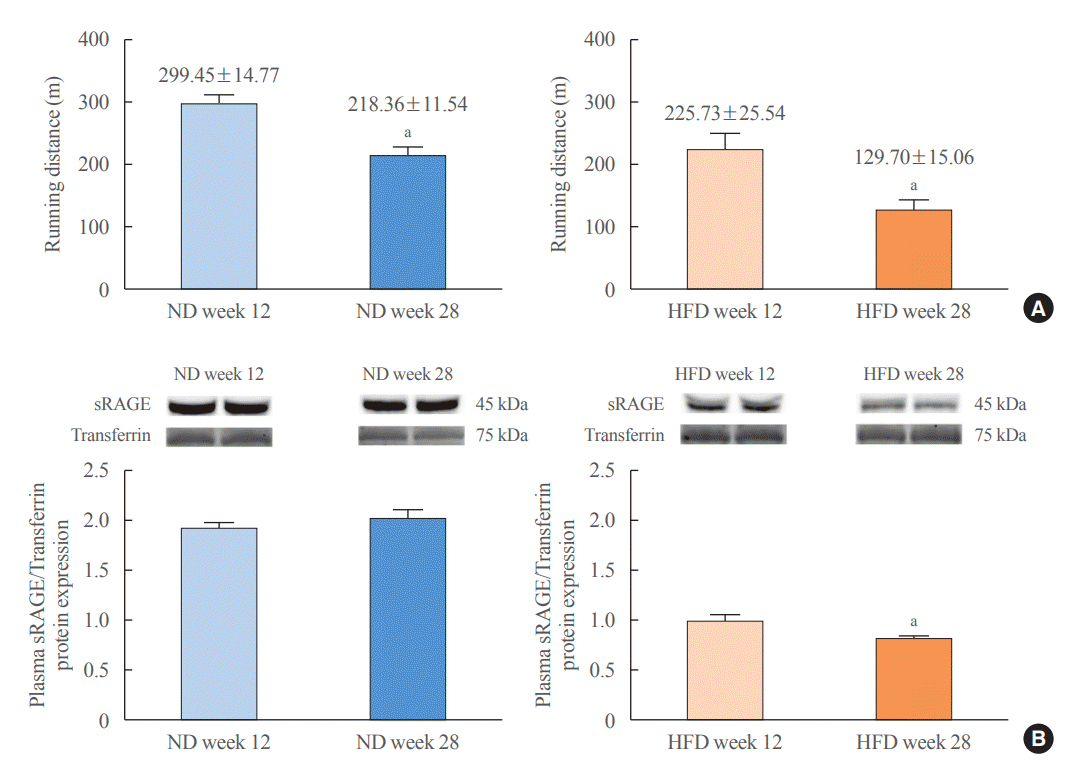
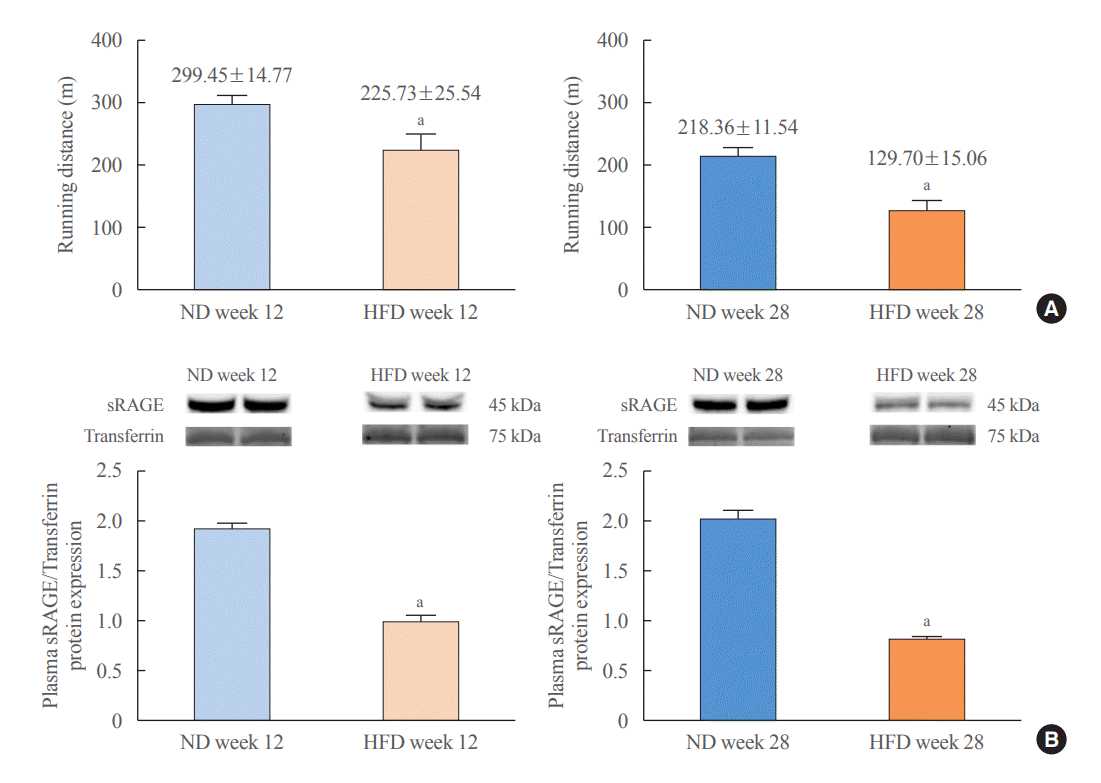
Both ND- and HFD-fed rats exhibited decreased CRF levels and a decrease in expression of soluble-receptor for advanced glycation end product (sRAGE), an anti-aging marker (Fig. 1). Moreover, metabolic parameters and cardiac function deteriorated from baseline to follow-up in both groups of rats (details shown in Supplemental Table S1). These changes were more pronounced in HFD-fed rats than in ND-fed rats, and some parameters were altered only in HFD-fed rats, while those of ND-fed rats remained unchanged from baseline to follow-up (details shown in Supplemental Table S1). These results collectively support the proposal that obesity induces premature aging [12].
Aging-related unfavorable metabolic, cardiac, and brain profiles in HFD-fed rats, indicating obesity-induced premature metabolic, cardiac, and brain aging
HFD-fed rats exhibited lower CRF levels and plasma sRAGE protein expression than found in ND-fed rats (Fig. 2). More unfavorable metabolic, cardiac, and brain parameters were observed in HFD-fed rats than in ND-fed rats (details shown in Supplemental Tables S2-S4, Supplemental Figs. S2, S3). Since these unfavorable profiles are also associated with aging [17-19], our results support the proposal that obesity results in premature metabolic, cardiac, and brain aging [12,20,21].
The protective effect of high CRF against metabolic impairment
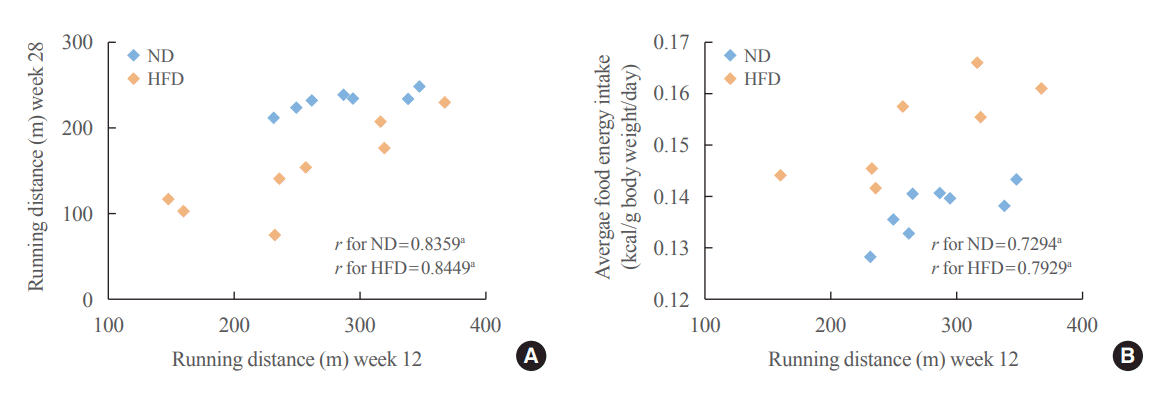
The relationship between CRF levels at baseline and at follow-up was investigated. There was a positive correlation between baseline CRF levels and CRF levels at follow-up in both ND-and HFD-fed rats (Fig. 3A), providing support that CRF is closely linked to genetics [2]. Interestingly, CRF showed a positive correlation with average food energy intake (Fig. 3B). This finding suggests that the effect of high CRF on protection against obesity is independent of calorie intake.
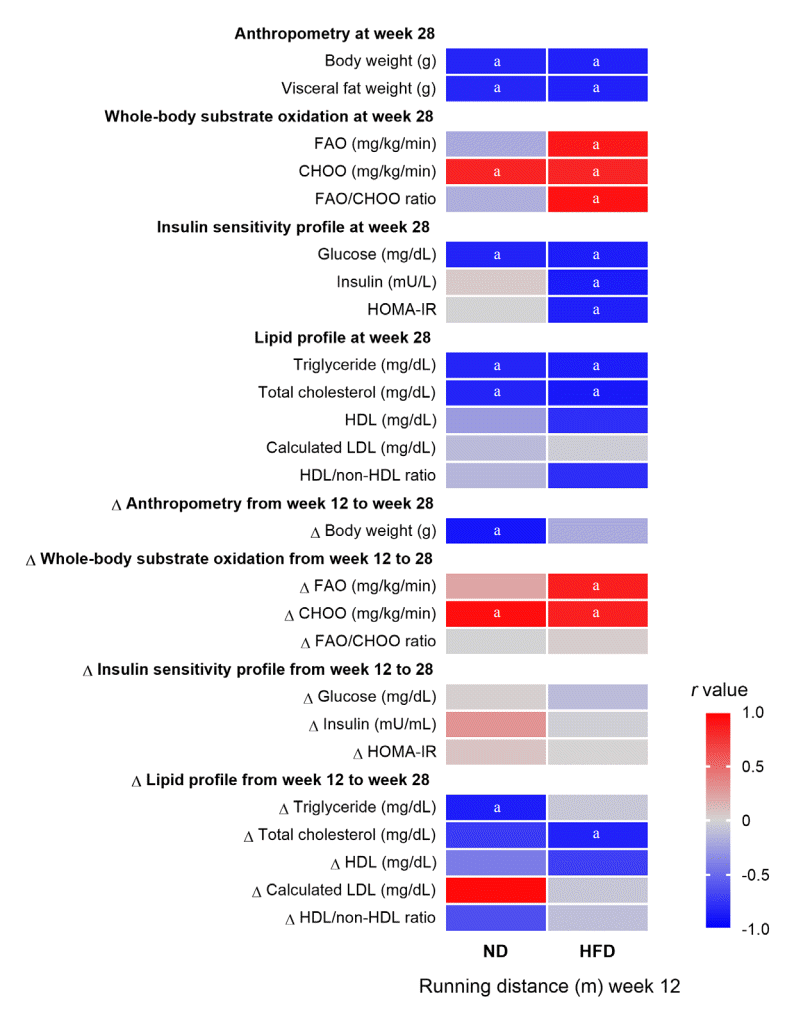
To determine the mechanisms underlying the protective effect of high CRF against metabolic dysfunction, we investigated the relationships of baseline CRF levels with metabolic parameters at follow-up and the absolute change of metabolic parameters from baseline to follow-up (value at week 28–value at week 12). We found that high baseline CRF levels were positively correlated with favorable metabolic parameters at follow-up, including favorable body weight and composition, increased wholebody substrate oxidation during exercise, decreased insulin resistance, and favorable lipid profiles (details shown in Fig. 4). The subgroup analyses also showed statistically significant differences in these parameters between rats with higher baseline CRF levels (ranked first to fourth in baseline running distance; n=4) and those with lower baseline CRF levels (ranked fifth to eighth in baseline running distance; n=4) within each diet group (details shown in Supplemental Table S5). Interestingly, the correlations between baseline CRF levels and metabolic parameters at follow-up were more obvious in HFD-fed rats than in ND-fed rats. Specifically, some metabolic parameters showed correlations with CRF levels at baseline only in HFD-fed rats, but not in ND-fed rats (details shown in Fig. 4). However, there were no significant correlations between changes in CRF levels from week 12 to 28 with metabolic parameters in the follow-up data. Importantly, the relationships between baseline CRF level and metabolic parameters were exhibited when the rats were only 35 weeks old, suggesting that high CRF protects against metabolic impairment starting in early life.
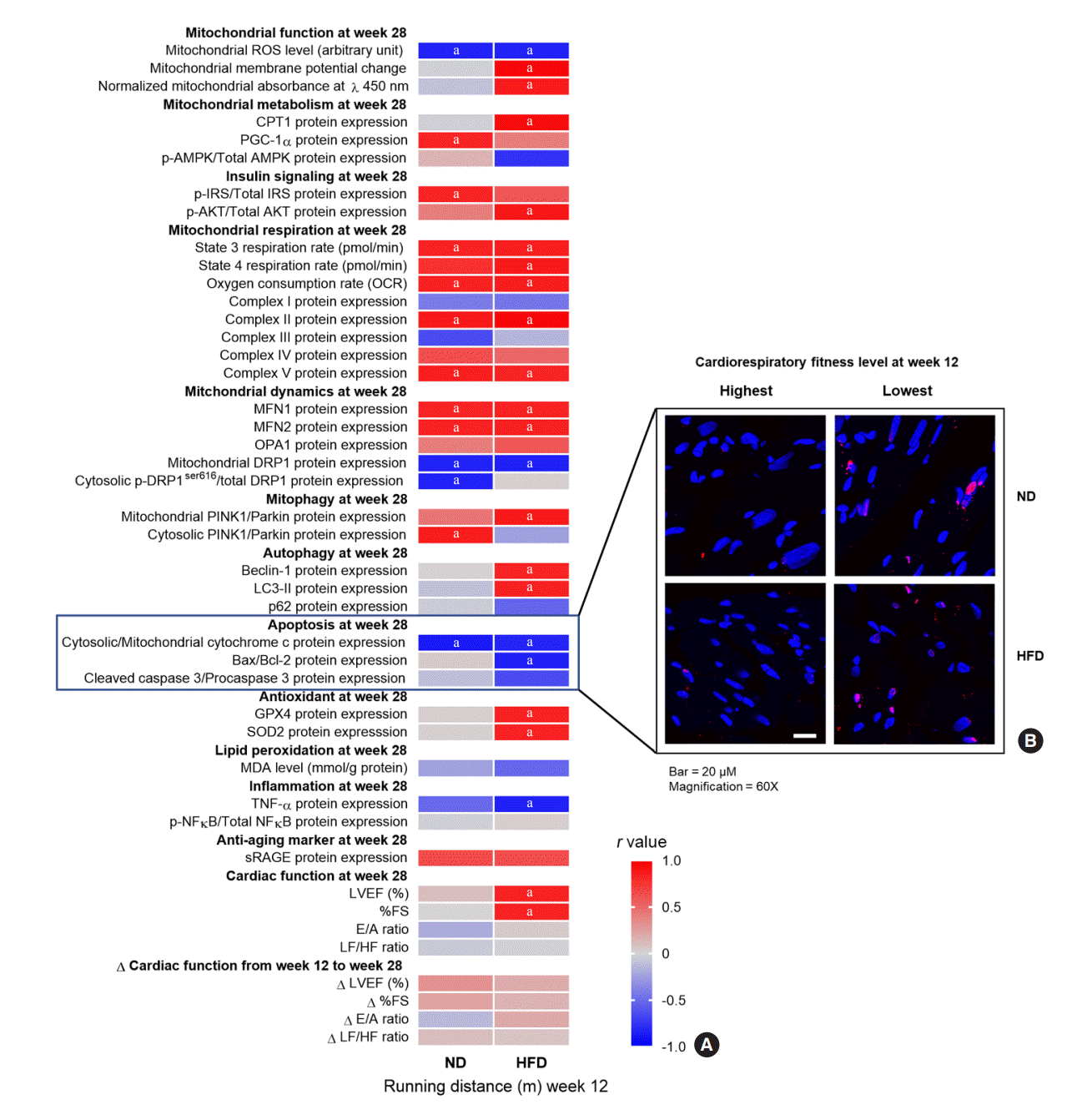
Mechanisms of the protective effect of CRF against cardiac impairment
To determine the mechanisms responsible for the protective effect of high CRF against cardiac impairment, we identified the relationships between baseline CRF levels and cardiac parameters at follow-up. We also evaluated the relationships between baseline CRF levels and changes in cardiac function from baseline to follow-up. High CRF levels at baseline were positively correlated with favorable cardiac parameters at follow-up, including improved mitochondrial function, favorable mitochondrial metabolism, increased insulin signaling, increased mitochondrial respiration, decreased mitochondrial fission, increased mitochondrial fusion increased mitophagy, increased autophagy, decreased apoptosis, increased antioxidative capacity, decreased lipid peroxidation, decreased inflammation, increased anti-aging markers, and favorable cardiac function (details shown in Fig. 5, Supplemental Figs. S4, S5). The subgroup analyses also demonstrated significant or nearly-significant (P=0.05 to 0.09) differences in these parameters between rats with higher baseline CRF levels (ranked first to fourth in baseline running distance; n=4) and those with lower baseline CRF levels (ranked fifth to eighth in baseline running distance; n=4) within each diet group (details as shown in Supplemental Table S6). Specifically, the relationships between baseline CRF levels and cardiac parameters at follow-up were more obvious in HFD-fed rats than in ND-fed rats, since some cardiac parameters were only correlated with baseline CRF levels in HFD-fed rats, but not in ND-fed rats (details shown in Fig. 5, Supplemental Fig. S5). However, no significant correlations between changes in CRF levels from week 12 to 28 and cardiac parameters at follow-up were observed. Interestingly, the relationships between baseline CRF levels and cardiac parameters were displayed when the rats were only 35 weeks old, indicating that high CRF protects against cardiac impairment starting in an early period of life.
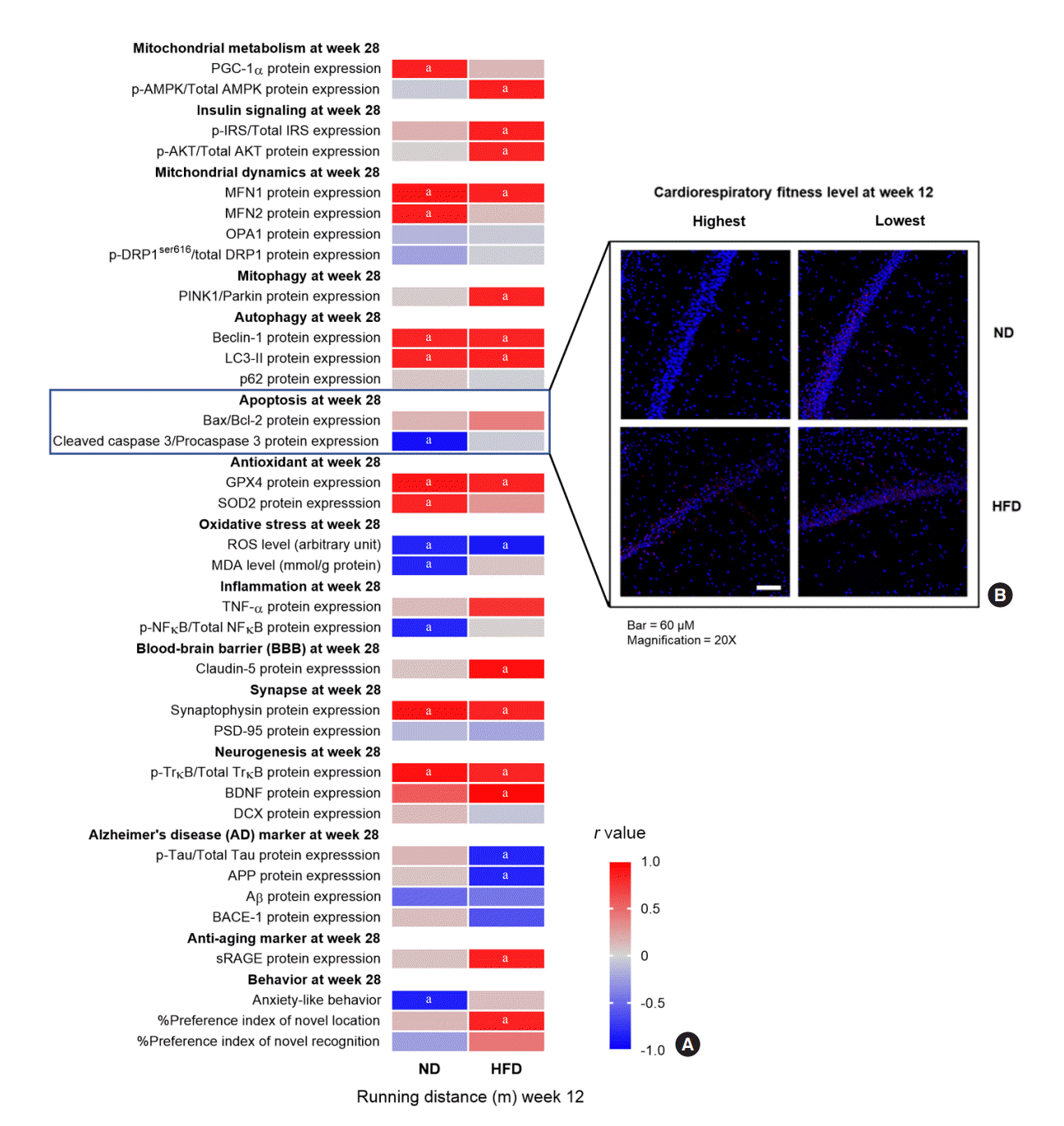
Mechanisms of the protective effect of high CRF against brain impairment
To determine the mechanisms underlying the protective effect of high CRF against brain impairment, we investigated the relationships between baseline CRF levels and brain parameters at follow-up. There were positive relationships between high baseline CRF levels and favorable brain parameters, including favorable metabolism, increased insulin signaling, decreased mitochondrial fission, increased mitochondrial fusion, increased mitophagy, increased autophagy, decreased apoptosis, increased antioxidative capacity, decreased oxidative stress, improved blood-brain barrier (BBB) function, improved synaptic function, increased neurogenesis, decreased AD markers, increased levels of an anti-aging marker, and improved cognition (details shown in Fig. 6, Supplemental Figs. S6, S7). The subgroup analyses also revealed a significant difference in these parameters between rats with higher baseline CRF levels (ranked first to fourth in baseline running distance; n=4) and those with lower baseline CRF levels (ranked fifth to eighth in baseline running distance; n=4) within each diet group (details shown in Supplemental Table S7). The relationships between baseline CRF levels and brain parameters at follow-up were more pronounced in HFD-fed rats than in ND-fed rats, as indicated by the findings that some brain parameters showed correlations with baseline CRF levels only in HFD-fed rats, but not in ND-fed rats (details shown in Fig. 6, Supplemental Fig. S7). Nevertheless, no significant correlations were found between changes in CRF levels from week 12 to 28 and brain parameters at follow-up. Importantly, the relationships between baseline CRF levels and brain parameters were exhibited when the rats were only 35 weeks old, suggesting that high CRF protects against brain impairment starting at an early stage of life.
DISCUSSION
In the present study, baseline CRF levels showed correlations with metabolic, cardiac, and brain parameters at the point of follow-up in both ND- and HFD-fed rats. We also found that HFD-fed rats exhibited aging-related unfavorable metabolic, cardiac, and brain profiles at follow-up, when compared with those of ND-fed rats. Taken together, it can be postulated that high CRF protects against metabolic, cardiac, and brain impairments both in normal conditions and in obesity-induced premature aging. Interestingly, the effect of high CRF as a protective mechanism against these impairments remained significant even though the follow-up duration was short (16 weeks) and the age of rats at follow-up was only 35 weeks (equivalent to less than 30 years in humans) [22], suggesting that the impact of CRF begins early in life.
We found significant correlations between baseline CRF levels and a variety of metabolic parameters at the time of follow-up, suggesting that high CRF delays metabolic impairment via the amelioration of obesity, impaired carbohydrate and fatty acid oxidation during vigorous exercise, reduced insulin resistance, and dyslipidemia. All of these findings have been reported in previous studies using HCR and LCR rats [7,8,11]. However, in our study, using non-selectively bred rats with a short duration of follow-up, it was found that metabolic parameters were significantly affected by CRF and the consequences of high CRF occurred early in life. The parameters that CRF appeared to have significantly impacted were body weight, visceral fat weight, fasting glucose, carbohydrate oxidation, triglyceride levels, and total cholesterol levels, as the correlations between baseline CRF levels and these parameters at follow-up were present in both ND and HFD-fed rats. Additionally, a comparison of the effect of high CRF against metabolic impairment between ND and HFD-fed rats demonstrated that this effect was stronger in obesity-induced premature aging. These findings support the proposal that high CRF confers protection against HFD-induced metabolic syndrome [23,24].
Our results showed relationships between baseline CRF levels and several cardiac parameters at the time of follow-up. These parameters could reflect the potential for several mechanisms to underlie the protective effect of high CRF against cardiac impairment, including mitochondrial function, metabolism, mitochondrial respiration, insulin signaling, mitochondrial dynamics, mitophagy, autophagy, apoptosis, redox reaction, inflammation, and systolic function. Interestingly, the effect of high CRF on protection against cardiac impairment was stronger in obesity-induced premature aging. Unlike some previous studies in HCR and LCR rats [13,14], we did not observe a relationship between baseline CRF levels and diastolic function at follow-up in normal and obese rats, and a relationship between baseline CRF levels and systolic function at follow-up was also not exhibited in normal rats. The contradictory results may be explained by the short follow-up in our study, for which reason the rats did not reach the age classified as elderly, when these correlations may have been exhibited. Nonetheless, our study, using non-selectively bred rats with a short follow-up, identified the cardiac parameters that were significantly affected by CRF levels early in life. These parameters were mitochondrial aspects, insulin signaling, and apoptosis, for which correlations with baseline CRF levels were observed in both ND- and HFD-fed rats.
We also observed relationships between baseline CRF levels and several brain parameters at follow-up, which may be mechanistically related to the protective effect of high CRF against brain impairment. These mechanisms included metabolism, insulin signaling, mitochondrial fusion, mitophagy, autophagy, apoptosis, redox reactions, inflammation, BBB, synaptic function, neurogenesis, AD markers, an anti-aging marker, anxiety-like behavior, and learning behavior. Consistent with metabolic and cardiac parameters, the protective effect of high CRF against brain impairment was stronger in obesity-induced premature aging. Since there were relationships between baseline CRF levels and mitochondrial aspects, autophagy, redox reactions, synaptic function, and neurogenesis in both ND and HFD-fed rats at follow-up, these brain parameters are considered to be closely affected by the level of CRF, particularly in early life.
First of all, this study cannot explain why the protective effect of high CRF against metabolic, cardiac, and brain impairment was stronger in obesity-induced premature aging. To further develop this line of inquiry, an additional molecular study comparing metabolic, cardiac, and brain profiles between normal and obese animals with the same level of baseline CRF would be needed. In addition, this study used an obesity-induced premature aging model that may not be representative of other premature aging models; hence, the findings may not be transferable. For that reason, a further study with various models of premature aging would be useful to determine the protective effect of high CRF against different pathological conditions. Additionally, we included only male rats in this study to control for sex; however, it is widely accepted that sex hormones are critical in determining lean body mass [25,26], and consequently CRF levels [27]. Hence, the findings of our study may not be transferable to clinical practice for female patients.
In conclusion, the protective effect of high CRF against metabolic, cardiac, and brain impairments was stronger in obesity-induced premature aging than in normal conditions. These findings imply that high CRF is a crucial protective factor against metabolic syndrome, cardiovascular diseases, and neurodegenerative disorders, especially in conditions of obesity. Moreover, because our molecular study indicated that these effects of CRF began at a relatively young age, high CRF might exhibit early protection against metabolic, cardiac, and brain impairments via various molecular mechanisms. Taken together, any interventions that help increase CRF levels starting in an early period of life would be considered highly beneficial, specifically in obese individuals. Several studies have reported that CRF was increased after exercise training [28-31]. Therefore, our findings emphasize the benefits of exercise training on metabolic, cardiac and brain health conditions.
 XML Download
XML Download