

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Cheonggukjang is one of the most popular fermented soybean foods consumed in Korea. The production process of Cheonggukjang is classified as modern or traditional. Fermented soybean products, including Cheonggukjang, are well-known healthcare food for inflammation, cardiovascular diseases, obesity, and constipation [12]. Although the efficacy of Cheonggukjang is well known, there are several limitations because of problems in the fermentation process. Harmful substances, such as hazardous (food poisoning) microorganisms and biogenic amines (BAs), can be generated during the fermentation process [3].
BAs are made during the process of spoiling or fermenting foods containing protein-rich materials. Several studies have suggested that the upper limits for histamine (100 mg/kg), tyramine (100–800 mg/kg), β-phenylethylamine (30 mg/kg), and 1,000 mg/kg for the total BAs in foods [456]. The excessive intake of BAs stimulates the nervous and vascular systems, but it has been argued that it causes food allergies [57]. Although high levels of BAs have been reported in some traditional Cheonggukjang [78], no significant toxicity of fermented soybean products has been reported in vitro or in vivo [9]. To the best of our knowledge, there are no clear results on whether the efficacy can be reduced by toxin and BAs in Cheonggukjang. Traditionally, Cheonggujkang has been produced by fermentation using Bacillus, which exists in rice straw. Bacillus improves gastrointestinal function. Functional materials, such as dietary fiber, phosphatide, isoflavone, flavonoids, phenolic acids, and poly-glutamic acid, were increased during fermentation [10].
Constipation is a prevalent chronic gastrointestinal disorder that is characterized by dry and hard stools, persistent, infrequent, or incomplete defecation [11]. Approximately 70% of people have gastrointestinal disorders caused by changes in the living environment and eating habits in modern society, and most of them have constipation. The beneficial microorganisms and dietary fiber in Cheonggukjang can improve the intestinal function and relieve constipation [2].
This study examined whether toxins and BAs in Cheonggukjang, HTBC and LTBC, affect their efficacy in a mice model of Lop-induced constipation. Their side effects were monitored by changes in the body weight, food intake, water consumption, and behavior in model mice. The laxative effects of HTBC and LTBC were compared using the stool number, fecal water content, intestinal charcoal transit, histological changes in colon tissue, and their underlying mechanisms.
MATERIALS AND METHODS
Chemicals and antibodies
The chemicals were obtained from the following sources: Loperamide hydrochloride (Lop), prucalopride, sodium chloride, activated charcoal, and aqueous methylcellulose from Sigma-Aldrich (St Louis, MO, USA). Rabbit monoclonal antibody against phospho-extracellular signal-regulated kinase (ERK) 1/2 (Thr202/Tyr204), rabbit polyclonal antibodies against phospho-p38 (Thr180/Tyr182) and phospho-SAPK/JNK (Thr183/Tyr185), horseradish peroxidase (HRP)-conjugated anti-rabbit, and anti-mouse immunoglobulin G (IgG) antibodies were purchased from Cell Signaling Technology Inc. (Beverly, MA, USA). Rabbit anti-c-Kit and anti-SCF antibodies were obtained from Abcam (Cambridge, MA, USA). The BCA protein assay kit was obtained from Thermo Scientific (Waltham, MA, USA). The polyvinylidene difluoride (PVDF) membrane and enhanced chemiluminescence substrate were obtained from Amersham Pharmacia Biotech. The Hybrid-R™ RNA extraction kit was purchased from GeneAll Biotechnology Co., Ltd (Seoul, Korea). The PrimeScript™ 1st strand cDNA Synthesis kit and TB Green® Premix Ex Taq™ II (Tli RNaseH Plus) were acquired from Takara (Takara, Japan). All other chemicals were of analytical grade or complied with the standards.
Next-generation sequencing analysis
Cheonggukjang, containing a high and low level of toxins and biogenic amines (HTBC and LTBC), was obtained from Sunchang (Jeonbuk, Korea). The total genomic DNA was extracted from each sample using a FastDNA Spin Kit (MPBio, USA), according to the manufacturer’s instructions. The universal primer pairs 27F (5′-CCTATCCCCTGTGTGCCTTGGCAGTC (adapter)-TCAG (key)-AC linker-GAGTTTGATCMTGGCTCAG-3′) and 518R (3′, 5′-CCATCTCATCCCTGCGTGTCTCCGAC (adapter)-TCAG (key)-Barcode-AC linker-WTTACCGCGGCTGCTGG-3′) were conducted to 16S rDNA genes with the following conditions of polymerase chain reaction (PCR) amplification: initial denaturation at 94°C (5.5 min), 60°C (45 s), and 10 cycles of 72°C for 90 s. The procedure was followed by denaturation for 94°C (30 s), 55°C (45 s), and 20 cycles of 72°C for 90 s.
Determination of salt and BAs
The salt content was measured using a salinity meter (TM-30D, Takemura, Japan). The BAs levels were based on that described by García-García et al. [12]. Five grams of the paste containing 15 mL of 0.1 N-HCl, were homogenized and centrifuged (4,000 ×g, 4°C, 15 min). The supernatant was pooled and filtered, and 0.1 N-HCl was added to 50 mL. The standards (histamine and tryptamine) and samples were derivatized using saturated Na2CO3 and 1% trichloroacetic acid. The content of BAs was analyzed using a nanospace SI 2 HPLC system (Shiseido Co., Tokyo, Japan). The analysis was performed on a UG 120 column (4.6 × 250 mm, 5 μm) applying gradient elution, using water (A) and acetonitrile (B) (Thermo Scientific). The injection volume was 10 μL and the flow rate was 1 mL/min at 40°C. The gradient was performed as follows: 10 min 55% B, 15 min 65% B, 25 min 80% B, and 30 min 90% B. The BAs were detected at ultraviolet 254 nm.
Lop-induced constipation mice model
ICR mice (male, bodyweight 22–25 g) were purchased from Damul Science (Daejeon, Korea). During the experiment, the mice were randomized into five groups (n = 10) and housed under a strict light cycle (12 h light/dark cycles) at 22 ± 2°C and 55 ± 5% relative humidity. The mice were acclimatized to the laboratory conditions for 1 week before the experiments and allowed free access to potable water and food. All animals were strictly cared for according to the committee guidelines, and all experimental procedures were reviewed and approved by the Animal Care Committee of Jeonju AgroBio-Materials Institute (JAMI IACUC 2020002). As shown in Fig. 1, the ICR mice were divided randomly into a non–constipation or experimental groups.
Fig. 1
Schematic protocol for animal experiments.
Lop, loperamide; PC, positive control; HTBC, high level of toxins and biogenic amines containing Cheonggkujang; LTBC, low level of toxins and biogenic amines containing Cheonggkujang.
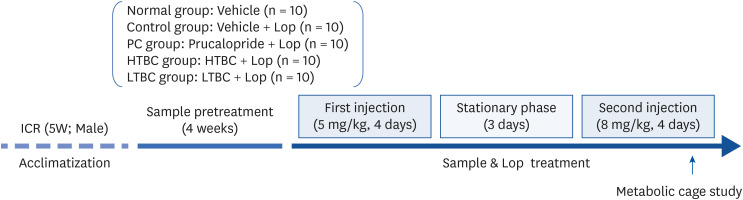
The Lop-induced constipation groups were divided further into 4 groups as follows: vehicle, prucalopride (PC; positive control, 5 mg/kg), HTBC (500 mg/kg), and LTBC (500 mg/kg) treated group. Constipation was induced, as described in previous reports, with some modification [13]. Briefly, Lop (0.9% in sterile saline) was administered orally twice a day for 4 days at a dosage of 5 mg/kg. After resting for 3 days, an increased dose of Lop (8 mg/kg) was administered in the same manner for 4 days. The normal group was treated with the same volume of saline. Before one hour, HTBC and LTBC were administered orally with final doses of 500 mg/kg. Each sample was orally administered daily for four weeks before inducing constipation. The normal and control groups received saline with the same volume, whereas the PC group received a prucalopride in saline solution. The body weights of the mice were recorded once a week.
Measurement of food intake, water intake, fecal, and urine parameters
The food intake, water consumption, fecal and urine parameters in the individual mice were measured on the 10th day (D10) after the Lop treatment using a metabolic cage (Fig. 1). The mice were housed individually in metabolic cages (Jeung Do Bio & Plant Co. LTD., Seoul, Korea) to analyze the food and water intake and fecal and urine output over 24 h. The feces weight and number were weighed and counted 3 times using an electric balance and manual counter. The fecal water content was determined by drying the collected fecal pellets to a constant weight for 24 h at 60°C, and the fecal dry weights were determined. The fecal water content was analyzed using the following formula:
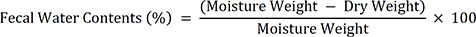
Analysis of intestinal charcoal transit ratio
At the end of the experiment, the mice were fasted for 18 h and then administered a charcoal meal orally (5% aqueous suspension of activated charcoal and 10% gum arable) to analyze the intestinal charcoal transit ratio. A 60 min after the charcoal meal administration, the mice were sacrificed humanely via cervical dislocation, and the small intestine between the pyloric and the cecum was quickly removed. The charcoal transit ratio was analyzed using the following equation by measuring the charcoal meal transit distance and the total length of the small intestine:
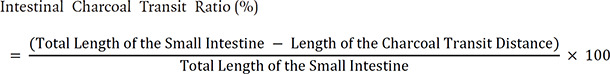
Western blot analysis
The total homogenate proteins were extracted from the colons using the ice-cold radioimmunoprecipitation (RIPA) buffer containing PMSF, and protein concentrations were performed with a BCA protein assay kit according to the manufacturer’s instructions. The proteins were separated by electrophoresis on 10% or 12% sodium dodecyl sulphate–polyacrylamide gel electrophoresis gel and transferred onto PVDF (GE Healthcare, Little Chalfont, Buckinghamshire, UK) membranes. After blocking with 5% non-fat milk in TBST (50 mmol/L Tris-HCl, pH 7.5, 150 mmol/L NaCl, and 0.1% Tween 20) for 1 h at room temperature, the blots were incubated with the primary antibodies at 1:2,500 dilutions overnight at 4°C. The blots were rinsed 4 times with TBST and incubated with HRP-conjugated anti-rabbit IgG or anti-mouse IgG antibodies at dilutions of 1:5,000 for 1 h at room temperature. The blot was developed using an enhanced chemiluminescence (ECL) system (Bio-Rad, Munich, Germany). Quantitative analysis was visualized by densitometric scanning (Amersham imager 600; GE Healthcare).
Quantitative PCR assay
Gene expression of the colons was then assessed by quantitative real time-PCR (qPCR) using the total RNA. The total RNA from the chopped colon tissue was isolated using a Hybrid-R™ (GeneAll Biotechnology Co., Ltd) according to the manufacturer's instructions. Complementary DNA (cDNA) was synthesized using a Prime Script™ 1st strand cDNA Synthesis kit and amplified for PCR. Table 1 lists the sequences for the primers used in qPCR. The real-time PCR reaction was performed with the TB Green® Premix Ex Taq™ II (Tli RNaseH Plus). qPCR was performed in triplicate using β-Actin as a standard.

Histological analysis
Mouse colons tissues were fixed with 4% paraformaldehyde for 24 h, and dehydrated with alcohol using routine procedures. The sections were cut into 4 μm thick slices that were stained with hematoxylin and eosin (H&E) or alcian blue to general histopathological changes or mucin staining, respectively.
The morphological features of the stained sections were observed by optical microscopy (Olympus, Tokyo, Japan) and photographed. The thickness of the muscle and mucosal layer and inflammatory cell infiltration index were analyzed quantitatively using Image J software (National Institutes of Health, Bethesda, MD, USA).
Statistical analyses
The data are expressed as the means ± SD and all statistical analyses were performed with Sigmaplot v14.0 software (Systat Software Inc., San Jose, CA, USA). Statistical analysis was applied to identify the differences, followed by one-way analysis of variance (ANOVA) and Tukey’s honestly significant difference (HSD) post-hoc tests. A value of P < 0.05 was considered significant.
RESULTS
Microorganisms, BAs, and salt contents in Cheonggkukjang, HTBC, and LTBC
The microbiota compositions in each Chonggkukjang were analyzed by 16S rRNA sequencing on genomic DNA isolated from the samples. Fig. 2 shows the microbial community in 2 Cheonggkujang products, HTBC (Fig. 2A) and LTBC (Fig. 2B). The most common species in HTBC and LTBC were Bacillus species. While the hazardous species, Acinetobacter baumannii 17.4%, Cronobacter sakazakii 13.9%, and Enterobacter cloacae 3.7%, were presented in HTBC (Fig. 2A). The BAs, histamine (His) and tyramine (Tyr), were analyzed in both Cheonggukjang. The level of His was 0.8 and 3.9 mg/kg. Otherwise, Tyr was 494.6 and 240.5 mg/kg in HTBC and LTBC, respectively. The salt content was 0% and 0.3% in HTBC and LTBC, respectively (Table 2).
Fig. 2
Relative abundance of microorganisms in (A) HTBC or (B) LTBC.
HTBC, high level of toxins and biogenic amines containing Cheonggkujang; LTBC, low level of toxins and biogenic amines containing Cheonggkujang.


Effects of HTBC and LTBC on the feeding behavior on Lop-induced constipation mice
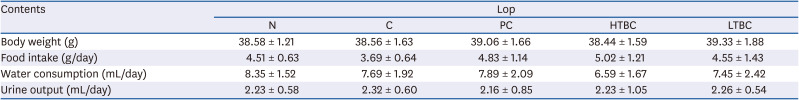
The influence of HTBC and LTBC on constipation was examined by establishing a mouse model using Lop after HTBC and LTBC administration for four weeks at a dose of 500 mg/kg (Fig. 1). Prucalopride (5 mg/kg) was used as a positive control. The body weight, food intake, water consumption, and urine output showed no significant differences among the experimental groups (Table 3). HTBC and LTBC did not show toxicity or cause other major changes in the general feeding behaviors and mortality.
Table 3
Effect of HTBC and LTBC on the body weight, food intake, and urine output in constipation mice
Effects of HTBC and LTBC on fecal parameters
The effects of HTBC and LTBC on Lop-induced constipation were assessed using the fecal number and water content. As shown in Fig. 3A, the fecal matter in the Lop-induced constipation group was small, thin, dry, and hard compared to the stool in the normal group (N). On the other hand, the fecal matter in the PC, HTBC, and LTBC administered group was similar to that of the N group. After collecting the total number of fecal pellets at D10 for 24 h, Lop administration reduced the number and water content of the fecal pellets significantly compared to the N group (P < 0.001, Fig. 3). HTBC or LTBC counteracted the Lop-induced decrease in feces number and water content (Fig. 3B and C). The total number of fecal pellets was reduced by 55.2% in the Lop group (C, 43.44 ± 8.59) compared to the N group (78.70 ± 9.01), whereas the PC (72.10 ± 15.08), HTBC (63.11 ± 15.03), and LTBC (61.70 ± 12.75) treatment groups increased by 166.0%, 145.2%, and 142.0%, respectively, compared to the C group (Fig. 3B). The total water content of the fecal pellets decreased by 55.1% in the C group (24.20 ± 11.55) compared to the N group (43.90 ± 10.94). On the other hand, the water content increased by 195.6%, 182.5%, and 203.1% in the PC (47.34 ±8.33), HTBC (44.16 ± 19.98), and LTBC (49.16 ± 5.44) treated groups, respectively, compared to the C group (Fig. 3C). Cheonggkukjang products had a similar effect on the fecal parameters. These results suggest that the BAs of Cheonggukjang did not affect the constipation relief effect and had no side effects.
Fig. 3
Effects of HTBC and LTBC administration on the fecal parameters in mice with Lop-induced constipation. (A) Representative fecal pellet images collected over 24 h at D10 after Lop administration. (B) Total fecal number. (C) Total water content of fecal pellets.
Results were expressed as mean ± SD (n = 10 mice per group).
Lop, loperamide; N, normal control group; C, Lop control group; PC, positive control group; HTBC, high level of toxins and biogenic amines containing Cheonggkujang; LTBC, low level of toxins and biogenic amines containing Cheonggkujang.
***P < 0.001 vs. N group; #P < 0.05, ##P < 0.01, and ###P < 0.001, vs. C group.

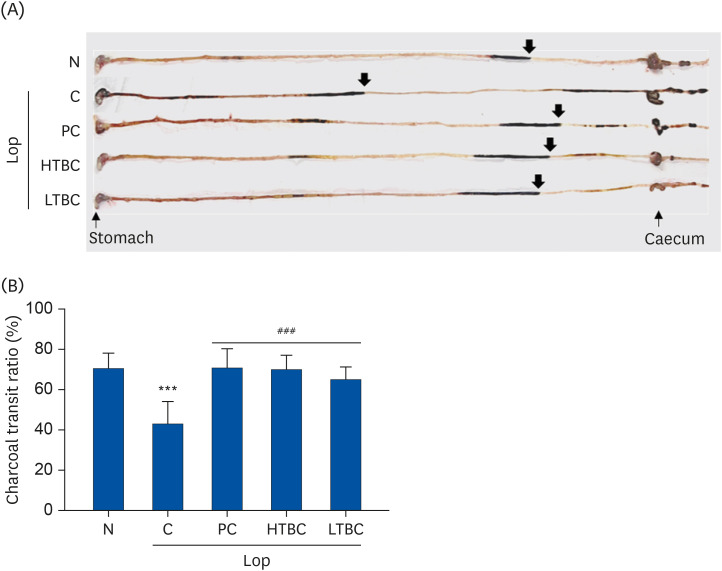
Effects of HTBC and LTBC on intestinal charcoal transit
A charcoal transit test was performed at D11 after the Lop treatment to determine if the increase in fecal excretion by HTBC and LTBC was accompanied by changes in intestinal mobility. As shown in Fig. 4, the impellent of the charcoal mobility decreased remarkably in the Lop-induced C group than those in the N group (P < 0.01). The propulsion of the charcoal meal was decreased by 43.3% in the C mice group compared to the N mice group (70.6%). On the other hand, the intestinal charcoal transit ratio increased significantly in the PC, HTBC, and LTBC groups (P < 0.001, Fig. 4B), compared to the C group. Although the charcoal transit ratio was low, the length of the intestine was similar (Fig. 4A). These results suggest that HTBC and LTBC administration can promote gastrointestinal motility in constipated mice.
Fig. 4
Effects of HTBC or LTBC administration on the intestinal transit ratio in constipated mice induced by Lop. (A) The representative image of charcoal transit in the intestine. The covered distances were measured after 60 min of administration with charcoal. The arrows indicate that the charcoal meal has migrated. (B) The transit ratio of charcoal was calculated using the total length of the intestine and the distance of the charcoal diet.
Results were expressed as mean ± SD (n = 10 mice per group).
Lop, loperamide; N, normal control group; C, Lop control group; PC, positive control group; HTBC, high level of toxins and biogenic amines containing Cheonggkujang; LTBC, low level of toxins and biogenic amines containing Cheonggkujang.
***P < 0.001 vs. N group; ###P < 0.001, vs. C group.
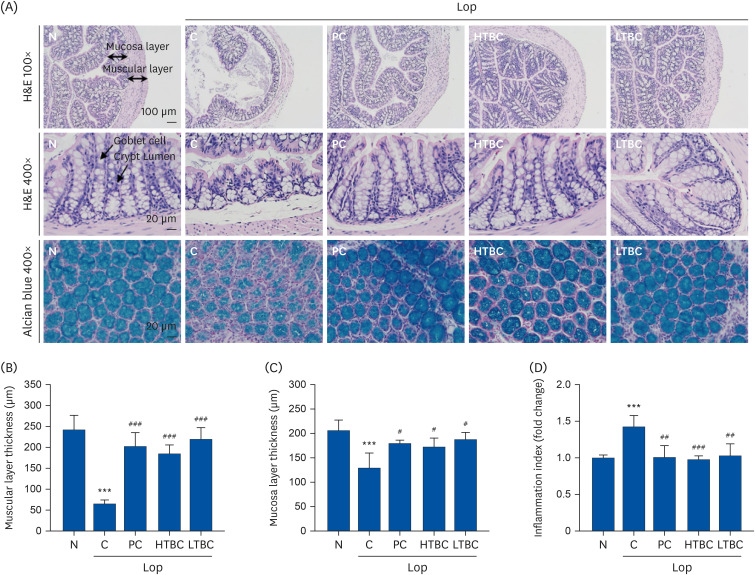
Effects of HTBC and LTBC on histological alterations of the transverse colon in Lop-induced constipation mice
The histopathological changes in the transverse colon tissue in the Lop-induced constipation model were analyzed by H&E and alcian blue staining. Fig. 5A (upper panel) shows the effects of HTBC and LTBC administration on the mucosal layer thickness in the Lop-induced constipation mouse tissue. The intestinal mucosa and muscular layer thicknesses were significantly lower in the Lop-induced C group than the N group (P < 0.001). On the other hand, HTBC and LTBC administration prevented the decrease in Lop-induced intestinal mucosa and muscular layer thickness. Inflammatory cells have infiltrated into Lop-induced constipated mice (Fig. 5D). In the Lop-induced constipation mice, the structure of the crypts was remarkably changed from open form to luminal face. In addition, an irregular shape, distribution, and size of the crypts goblet cells were observed in the Lop-induced constipation mice (Fig. 5A, middle panel). These changes were accompanied by the loss of mucus secretion in transverse colon sections analyzed by alcian blue staining (Fig. 5A, lower panel). The crypts of the mucosal layer in the colon were stained dark blue in the untreated group and light blue in the constipation group. On the other hand, the colons were stained similarly in the PC, HTBC, and LTBC treated groups. In conclusion, the mucous membrane and muscle layer thicknesses and mucus secretion were positively correlated with HTBC and LTBC administration.
Fig. 5
Histological structures and mucin secretion of the transverse colon in Lop-induced constipation mice after HTBC and LTBC administrations. (A) The mice were evaluated by hematoxylin and eosin (H&E), and alcian blue stain, and the observed sections were photographed at 100× and 400× magnification using a microscope. (B) Muscular and (C) Mucosa layer thickness and (D) Inflammation indices were calculated using the Image J image software.
Results were expressed as mean ± SD (n = 10 mice per group).
Lop, loperamide; N, normal control group; C, Lop control group; PC, positive control group; HTBC, high level of toxins and biogenic amines containing Cheonggkujang; LTBC, low level of toxins and biogenic amines containing Cheonggkujang.
***P < 0.001 vs. N group; #P < 0.05, ##P < 0.01, ###P < 0.001, vs. C group.
Effects of HTBC and LTBC on c-Kit and SCF expression in the colons of Lop-induced constipation mice
The interstitial cells of Cajal (ICCs) have been described as mesenchymal cells located between the intestinal nervous system and the smooth muscle, which play a central role in mediating the intestinal nerve signals to smooth muscle contractions [1415]. The major role of the ICCs in the gastrointestinal (GI) tract was functional activity by c-Kit and stem cell factor (SCF) signaling [16]. Therefore, quantitative PCR and immunoblotting for measuring the mRNA and protein levels of c-Kit and SCF were performed to examine the effects of HTBC and LTBC in the colon of Lop-induced constipation mice. As shown in Fig. 6, compared to the N group, the expression of c-Kit and SCF mRNA and the protein levels were decreased remarkably in the mouse colon after the Lop treatment. In contrast, the PC, HTBC, and LTBC administered group showed increased expression levels than the Lop group (P < 0.05). These results suggest that HTBC and LTBC inhibit Lop-induced c-Kit and SCF downregulation in mice.
Fig. 6
Effect of HTBC and LTBC administrations on the expression levels of Cajal markers in Lop-induced constipation mice. Relative mRNA levels of (A) c-Kit and (B) SCF were examined by quantitative PCR in colons of Lop-induced constipation mice. (C) The protein expression levels of the cajal markers (c-Kit and SCF) were analyzed by western blot analysis in the colons of Lop-induced constipation mice. The protein levels of (D) c-Kit and (E) SCF (relative density normalized to β-Actin) were measured using an imaging densitometer and followed by statistical analysis.
Results were expressed as mean ± SD (n = 10 mice per group). Differences between the groups were determined as statistical significance by one-way ANOVA followed by Duncan's multiple range test.
Lop, loperamide; N, normal control group; C, Lop control group; PC, positive control group; HTBC, high level of toxins and biogenic amines containing Cheonggkujang; LTBC, low level of toxins and biogenic amines containing Cheonggkujang; ANOVA, analysis of variance.
*P < 0.05, **P < 0.01, and ***P < 0.001 vs. N group; #P < 0.05, ##P < 0.01, and ###P < 0.001 vs. C group.

Effects of HTBC and LTBC on mitogen-activated kinase (MAPK) signaling in the colons of Lop-induced constipation mice
The activation of MAPKs was examined to elucidate the effects of HTBC and LTBC on the mechanism for transcriptional signaling protein expression. MAPKs, including ERK1/2, c-Jun N-terminal kinase (JNK), and p38, play a key role in the visceral smooth muscle contraction and regulating the cellular responses [17]. As shown in Fig. 7, the levels of ERK1/2, JNK, and p38 phosphorylation were increased in colon tissue after Lop-administration compared to the N group. In contrast, PC, HTBC, and LTBC had opposite effects on their expression levels compared to the Lop group. These results suggest that the MAPKs signaling pathways could be attributed to the effects of HTBC and LTBC after the Lop treatment.
Fig. 7
Effect of HTBC and LTBC administrations on MAPK signaling in Lop-induced constipation mice. The protein levels of the phospho and total ERK1/2, JNK, and p38 were analyzed in the Lop-induced constipation mice. The relative protein levels were quantified using an image analyzer. The band density was normalized using each total protein.
The results were expressed as mean ± SD (n = 10 mice per group). The differences between the groups were determined as statistical significance by one-way ANOVA followed by Duncan's multiple range test.
Lop, loperamide; N, normal control group; C, Lop control group; PC, positive control group; HTBC, high level of toxins and biogenic amines containing Cheonggkujang; LTBC, low level of toxins and biogenic amines containing Cheonggkujang; MAPK, mitogen-activated kinase; ERK1/2, extracellular signal-regulated kinase 1/2; JNK, c-Jun N-terminal kinase; ANOVA, analysis of variance.
***P < 0.001 vs. N group; #P < 0.05, ##P < 0.01, and ###P < 0.001 vs. C group.

DISCUSSION
Chonggkukjang is a food made by fermenting soybean with Bacillus species, which has existed in Korea for more than 2,000 years [18]. Many studies have reported that Chonggkukjang has anti-inflammatory, anti-obesity [19], anti-diabetic [20], anti-cancer, stroke, hypertension, osteoporosis prevention [18], and constipation relief effects [1821].
Despite the health benefits of Chonggkukjang, Bacillus contamination, BAs, or mycotoxins are major safety issues. As mentioned in the introduction, the upper limit of BAs in food is as follows: histamine (100 mg/kg), tyramine (100–800 mg/kg), and total BAs 1,000 mg/kg [47]. The range of BAs in Cheonggukjang was reported to be histamine 0.2–70.3 mg/kg and tyramine 0.7–2,467.8 mg/kg [722]. The method of reducing the BAs content in Cheonggukjang has been reported in several groups [37]. To the best of our knowledge, few studies have reported the physiological activity, side effects, and safety of Cheonggukjang containing BAs. This study examined whether the consumption of Cheonggukjang in the Korean market can alter the physiological activity and cause health risks depending on the histamine and tyramine content. This was achieved by investigating the efficacy and safety of 2 types of Cheonggukjang with different levels of microorganisms, BAs, and salt in a Lop-induced constipation mouse model. The microorganisms in HTBC and LTBC mainly consist of Bacillus, but HTBC contained hazardous microorganisms (35.1%), such as A. baumannii, C. sakazakii, and E. cloacae (Fig. 2). The tyramine and salt levels were also higher approximately 2–3 fold in HTBC than LTBC (Table 2), but they did not exceed the safety range [7]. A Lop-induced constipation model was designed, as shown in Fig. 1. The samples were administered orally for four weeks before Lop administration to analyze the side effects of HTBC and LTBC. No noticeable side effects were noted, including abnormal behavior, food intake, water consumption, and body weight changes (Table 3).
Constipation is a typical bowel disease in humans, due mainly to irregular and difficult bowel movements and dry feces [23]. Lop is a μ-opioid receptor agonist that prevents the release of acetylcholine and prostaglandins, inhibiting intestinal peristalsis and the excretion of intestinal fecal pellets [24]. The leading cause of constipation is decreased amount of water in the stool due to the long retention of stool in the intestine. This study used prucalopride, a serotonin (5-hydroxytryptamine receptor 4, 5-HT4) receptor agonist, as a positive control. 5-HT4 receptors play an important role in regulating the gastrointestinal motility, intestinal nerve signals, and visceral pain [25]. The results showed that both samples, HTBC and LTBC, had a significant effect on Lop-induced constipation in mice by reserving the fecal number, water content, and intestinal charcoal transition. The changes in pathological structures, such as muscle and mucous thickness, as well as mucin secretion in the colon, are important factors in assessing and determining the effectiveness of the constipation treatment. These results showed that the intestinal muscle and mucous thickness, including mucin secretion, was reduced significantly by the Lop treatment, which was recovered by HTBC and LTBC administration (Fig. 5). The interstitial cells of Cajal (ICCs) are involved in smooth muscle contraction and regulate gastrointestinal peristalsis. c-Kit is a specific marker for the normal function, which is the gastrointestinal motility and gastrointestinal movement of ICCs. The SCF is a ligand for the c-Kit receptor that maintains normal physiological function [152627]. HTBC and LTBC suppressed the downregulation of c-Kit and SCF in the colon of constipated mice (Fig. 6). Lop increased inflammatory cell infiltration in colon tissues, which was also reduced by HTBC and LTBC administration (Fig. 5).
The opioid-induced activation of MAPKs, which is associated with decreased gastrointestinal transition resulting in constipation [2829]. The subfamilies of MAPKs, ERK1/2, JNK, and p38, are important in the contractile response in inflamed intestinal smooth muscle [17]. The opioid receptors are a class of membrane proteins belonging to the G protein-coupled receptor (GPCR) bound to the heterotrimeric Gi/o protein. Studies showed that the opioid receptors activate MAPKs through cyclic adenosine 3,5-monophosphate, phosphatidylinositol-4,5-bisphosphate-3 kinase, protein kinase C, and intracellular calcium signaling cascade. Furthermore, activation of the opioid receptors triggers arrestin-dependent receptor desensitization that recruits alternative signaling cascades, including MAPK [3031].
Patients suffering from symptoms associated with altered gastrointestinal motility experience a decreased quality of life. Although several medications, including anti-muscarinic agents, acetylcholine-releasing drugs, 5-HT3 antagonists, 5-HT4 agonists, and dopamine D2 antagonists are currently available in clinical practice for GI motility disorders, there are still cases where their therapeutic efficacy is unsatisfactory. Several kinase inhibitors for MAPK, such as ERK1/2 or p38, are already used in clinical trials [17]. Therefore, blocking the MAPK signaling pathway with selective kinase inhibitors could be a target for developing therapeutics for GI motility disorders. The present study showed that HTBC and LTBC inhibited the MAPKs phosphorylation induced by Lop in mice.
In conclusion, toxins and BAs of Cheonggukjang did not have side effects by confirming that there was no weight change, decreased food intake, or abnormal behavior in the Lop-induced constipation mouse model. This study showed that both types of Cheonggukjang with different toxin and BAs contents improved the gastrointestinal transition, fecal number, and fecal water content in Lop-treated mice. In a histopathological evaluation, HTBC and LTBC administration increased muscle and mucus thickness in the colon of Lop-induced constipated mice. These results showed that BAs and toxins did not alter the effects of Lop-induced constipation relief.
 XML Download
XML Download