

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
As the aging population increases due to the recent increase in the world's population, interest in aging diseases has increased [1]. Accordingly, a healthy retirement life has become even more important. On the other hand, senile diseases caused by various problems, such as metabolism and homeostasis imbalance, threaten the quality of life of the elderly [2]. The initial symptoms of benign prostatic hyperplasia (BPH) are lower urinary tract symptoms (LUTS), including nocturia, intermittent, weak flow, urgency, frequency, and incomplete emptying [3]. Therefore, the progression of BPH can lead to the loss of bladder function and impaired kidney function [4]. In general, the prostate is an androgen-dependent organ, so androgens are essential for prostate growth and maintenance [5]. A sex hormone imbalance has been reported as one of the leading causes of BPH because androgens are relatively decreased, and estrogen is relatively increased in the elderly; however, the precise cause is unknown [6]. According to several studies, the dihydrotestosterone (DHT) levels of BPH patients are higher than in normal men [7]. In the prostate, 5α-reductase (5AR) (nuclear membrane-binding enzyme steroid) converts testosterone to DHT, which is a more potent androgen than testosterone because of its high affinity for the androgen receptor (AR) [8]. The activated and modified DHT-AR complex migrates to the nucleus and interacts with the androgen-responsive element (ARE) [9]. The DHT-AR complex promotes the recruitment of co-activators, such as steroid receptor co-activator 1 (SRC1) or core pressers, to regulate gene expression by binding to the specific DNA response elements. Finally, the DHT-AR complex increases the protein levels by increasing the mRNA expression of growth factors, such as vascular endothelial growth factor (VEGF), epidermal growth factor (EGF), and insulin-like growth factor-1 (IGF-1) and prostate specific antigen (PSA) [10]. AR and PI3K/AKT signaling influence each other's feedback on prostate cell proliferation and death. Androgens can activate the PI3K/AKT pathway to improve the protein synthesis levels and promote proliferation and cell growth in the prostate tissue [11]. One of the representative cell proliferation factors, cyclin D1, plays a vital role in the G1/S progression of the cell cycle. [12]. In addition, proliferative cell nuclear antigen (PCNA) is an acidic nuclear protein recognized as a histological marker in the cell cycle [13]. Therefore, the expression of cyclin D1 and PCNA can reflect the state of the prostate cell proliferation for BPH. In the process of exogenous pathways, caspase is an effector protein that activates the downstream of the apoptosis signal in the mitochondria. The Bcl-2 family proteins act on the mitochondria or upstream of the apoptosis signal in relation to the mitochondrial membrane [14]. Among the Bcl-2 family, pro-apoptotic proteins, such as Bax, form pores in cytochrome c and other mitochondrial membranes [15]. The anti-apoptotic proteins, Bcl-2 and Bcl-xL, have been reported to interfere with this action while promoting the release of pro-apoptosis factors. Therefore, the Bax/Bcl-2 ratio determines cell death. In the case of BPH prostate tissue, the Bax/Bcl-2 ratio appears significantly lower than in normal tissue [16]. Overall, BPH leads to excessive prostate growth by overexpressing cell proliferation factors and downregulating the pro-apoptosis factors.
In modern medicine, patients with BPH with symptoms of LUTS require surgical or pharmacological treatment [17]. Currently, the most widely used pharmacological treatment is the prescription of 5α-reductase inhibitors (5ARI) and α-blockers [18]. The α-blockers are α1-adrenergic receptor antagonists that are commonly used in the treatment of BPH because of their ability to help urine flow by relaxing the smooth muscles of the prostate and bladder [19]. On the other hand, α-blockers do not help reduce the prostate size, so they do not treat the underlying cause. 5ARI, such as finasteride (Fi), inhibits the conversion of testosterone to DHT because it specifically inhibits 5α-reductase type2 (5AR2) [20]. As a result, it lowers the DHT content in the prostate, preventing DHT-AR binding, thereby inhibiting prostate growth. On the other hand, 5ARI has been reported to cause various side effects, such as ejaculation disorder, erectile dysfunction, and loss of appetite [21]. Therefore, there is a need for alternative medicines derived from natural substances with fewer side effects to help relieve BPH [22].
Abeliophyllum distichum Nakai (A. distichum) is a rare plant in the world's first genus, which grows only in Korea [23]. Currently, A. distichum is designated as a protected wild plant by the Ministry of Environment in Korea and is designated as a natural monument in 3 areas of Goesan-gun, Yeongdong-gun, and Buan-gun [24]. Since ancient times, A. distichum has been used for the treatment and prevention of stomach ailments, pest control, and inflammation [24]. To date, various research results have been published using A. distichum as well as food materials and pharmaceutical materials [24]. Recently, studies on the optimal extraction conditions, polyphenol compounds, and physiologically active substances are actively underway. According to Kwon et al.'s research [25], the polyphenol content in the leaves and stems of A. distichum is 50.64 mg/g and 13.53 mg/g, respectively. The flavonoid content is 96.47 mg/g in leaves and 18.53 mg/g in stems. Overall, the bioactive substances are high in A. distichum leaves. Chang reported that A. distichum leaves exhibited high antioxidant activity in antioxidant experiments, such as DPPH, ABTS, and reducing power because they contain abundant polyphenols and flavonoids [26]. In addition, according to Kwon et al.'s study [25], the LD50 value was judged to be more than 2,000 mg/kg because A. distichum leaves did not show genotoxicity in mice. In a recently reported study, A. distichum has showed efficacy in anti-osteoporosis [27], anti-colorectal cancer [28], anti-melanoma [29], anti-DNA damage related oxidative stress [30], anti-inflammatory [31] and anti-obesity [32]. On the other hand, no research results have been reported on BPH using A. distichum. Therefore, this study examined the effect of A. distichum on BPH.
Go to : 
MATERIALS AND METHODS
Materials
Testosterone propionate (TP) was provided by Tokyo Chemical Industry Co. (Tokyo, Japan). Fi (≥ 97% pure) and DHT (≥ 99% pure) were purchased from Sigma-Aldrich Inc. (St. Louis, MO, USA). The β-actin (sc-1616), SRC1 (sc-32789), PSA (sc-7638), AKT (sc-8312), PI3K (sc-1637), AR (sc-816), EGF (sc-374255), VEGF (sc-152), transforming growth factor-beta 1 (TGF-β1; sc-52893), PCNA (sc-56), and anti-goat IgG (sc-2005) antibodies were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). ER-α (#8644), Bcl-2 (#2870), Bax (#5023), cyclin D1 (#2978), anti-mouse IgG (#7076), and anti-rabbit IgG (#7074) antibodies were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). 5AR2 (ab124877), IGF-1 (ab106836) antibodies were purchased from Abcam Inc. (Cambridge, MA, USA). The DHT ELISA kit was purchased from SunLong Biotech Co. (Hangzhou, China). Roswell Park Memorial Institute medium (RPMI), fetal bovine serum (FBS), and penicillin/streptomycin were purchased from Gibco (Big Cabin, OK, USA).
Distilled water (DW) extraction
A. distichum leaves (100 g) and DW (1,000 mL) were added to an Erlenmeyer flask, then extracted by boiling for1 h. The liquid extract was concentrated using a rotary vacuum concentrator at 80°C. The extract was then filtered under reduced pressure (0.2 μm pore size) and lyophilized in a freeze dryer for 5 days.
70% ethanol extraction
A. distichum leaves (100 g) and 70% ethanol (1,000 mL) were added to an Erlenmeyer flask. After extracting 70% ethanol for 2 h, the extract was transferred to a beaker. Subsequently, 1,000 mL of ethanol was added and extracted for 2 h, and the following process was repeated 3 times. The 70% ethanol extract was then concentrated at 60°C using a rotary vacuum concentrator. The extract was then filtered under reduced pressure (0.2 μm pore size) and lyophilized for 5 days in a freeze dryer.
95% hexane extraction
A. distichum leaves (100 g) and 95% hexane (1,000 mL) were added to an Erlenmeyer flask. The 95% hexane solution was extracted for 6 h, and the extract was transferred to a beaker. Subsequently, 1,000 mL of 95% hexane was added, and the following process was repeated 3 times. The extract was then concentrated at 60°C using a rotary vacuum concentrator. The extract was then filtered under reduced pressure (0.2 μm pore size).
Cell culture and treatments
LNCaP (KCLB numbers: 21740) cells were purchased from the Korean Cell Line Bank (Seoul, Korea). The LNCaP cells were cultured in RPMI containing 10% FBS and 100 mg/mL penicillin/streptomycin at 37°C in a 5% CO2 incubator. The cells were then co-incubated with TP (100 nmol) and A. distichum leaf distilled water extract (ADLD), A. distichum leaf ethanol extract (ADLE), and A. distichum leaf hexane extract (ADLH) for 24 h. As the positive control, LNCaP cells were treated with Fi (1 μg/mL).
Immunofluorescence (IF)
Expression of AR post-treatment with ADLD, ADLE, ADLH (100 μg/mL), or Fi (1 μg/mL) was visualized by IF. Briefly, LNCaP cells (5 × 104) were seeded in slides overnight. After the treatment, the cells were fixed in ice-cold methanol followed by permeabilization with 0.1% Triton X-100 for 1 h. After blocking in 5% goat serum, the slides were incubated overnight with the AR antibody (1:300 dilution) at 4°C followed by incubation with the corresponding AlexaFluor 588 secondary antibody (1:1,000 dilution) for 1 h. The mounting medium containing the 4′,6-diamidino-2-phenylindole (DAPI) was then added to the slides, and coverslips were mounted. The images were captured using a fluorescent microscope from a Zeiss fluorescence microscope (Oberkochen, Germany).
Animal treatments and experimental design
Male Sprague-Dawley rats (Nara Biotech, Co., Ltd., Korea), weighing 200–220 g, were purchased and maintained at 40–60% humidity in an environment with an alternate 12-h day/night cycle and reared in cages for one week to undergo adaptation. Water and feed were provided ad libitum. All animal care procedures and experiments were approved by the Institutional Animal Care and Use Committee of the Konkuk University (KU20020). After an adaptation period, the rats were anesthetized by an intraperitoneal injection with phenobarbital (50 mg/kg). Subsequently, the rat's scrotum skin was incised. The vas deferens was tied with sutures, and the testes and epididymis were excised. The BPH-induced groups were then injected with TP (3 mg/kg/d) dissolved in corn oil 3 days after castration. The BPH model was induced by TP administration for 28 consecutive days. ADLE at a dietary concentration of 100 mg/kg/d was administered orally to rats. Saw palmetto extract (Saw) is generally known as a functional food with a beneficial effect on BPH among natural ingredients, so it was used as a positive control [3334]. Saw (100 mg/kg/d) and Fi (1 mg/kg/d) dissolved in water was administered orally. This experiment was conducted for 4 weeks. Blood samples were collected after the final injection. The prostate tissue was weighed after removing the surrounding fat and foreign substances. One part of the ventral prostate was fixed with 10% formalin and used for histological analysis. The other part of the ventral prostate was stored immediately in liquid nitrogen and used for the western blot experiment.
Hematoxylin and eosin (H&E) staining
H&E staining was performed as described previously [35]. The slides were examined using a Leica DMi8 Research Inverted Phase microscope (Leica, Werzlar, Germany), and the staining density was measured using ImageJ 1.47v software (National Institute of Health, Bethesda, MD, USA).
Immunohistochemistry
Sections with a thickness of 4 μm were deparaffinized and hydrated using a common xylene-alcohol series. For antigen retrieval, the sections were treated in a microwave oven for 10 min with 0.01 M citrate buffer (pH, 6.0), left to stand at room temperature for 10 min, and washed with DW. The endogenous peroxidase was quenched by a treatment with 3% H2O2. Subsequently, it was treated with normal serum from a secondary antibody host animal to block the non-specific binding. The sections were then incubated overnight at 4°C with anti-PCNA or anti-AR (Abcam, Cambridge, UK). After 1 h of treatment with the secondary antibody, the immunostained sections were counter-stained with Mayer's hematoxylin.
Western blotting assay
The Western blotting assay was performed as described previously [35]. Immunodetection was performed using an enhanced chemiluminescent horseradish peroxidase substrate (Advansta Inc., San Jose, CA, USA). Subsequently, the membranes were photographed using the Azure c300 imaging system (Azure Biosystems, Dublin, CA, USA). The chemiluminescent intensities of the protein signals were quantified using ImageJ 1.47v software (National Institute of Health).
Serum Concentrations of DHT
The blood collected from rats was centrifuged at 3,000 × g for 20 min at 4°C to obtain serum and stored at −80°C until the enzyme-linked immunosorbent assay (ELISA). The concentration of DHT in serum was measured using the DHT ELISA kit according to the manufacturer's instructions.
Statistical analysis
Statistical evaluations of the data were expressed as the mean ± SE of mean. The data were analyzed by SPSS version 11.5 for Windows (SPSS Inc., Chicago, IL, USA). The mean of 2 continuous normally distributed variables was compared using an independent samples Student's test. Dunnett's multiple range tests were used to compare the means of 2 and 3 or more groups of variables not normally distributed. P-values < 0.05 and < 0.01 were considered significant.
Go to :
RESULTS
Comparison of AR signal inhibitory effect by extraction methods of A. distichum
In this experiment, we compared the effects of 3 solvent extracts of A. distichum (DW, 70% ethanol, 95% hexane) on AR signaling in prostate cells (Fig. 1A). ADLE reduced the levels of AR, 5AR2, and PSA significantly, which are overexpressed by the TP treatment in prostate cells. IF confirmed that AR was overexpressed by the TP treatment. The IF experiment again confirmed that ADLE inhibits AR (Fig. 1B).
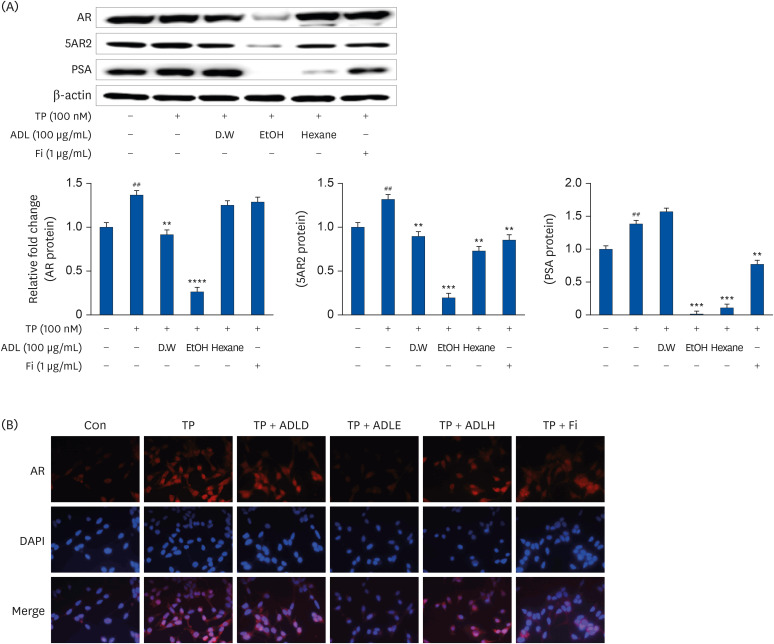 | Fig. 1Comparison of the AR signal inhibitory effect by extraction methods of A. distichum. (A) Representative western blot showing the bands of AR, 5AR2, and PSA. LNCaP cells were incubated for 24 h in a culture medium containing TP (100 nmol), ADL (A. distichum leaves, 100 μg/mL; DW, 70% EtOH or 95% hexane), or Fi (1 μg/mL). (B) IF detection of AR in LNCaP cells. LNCaP cells were plated on glass slides and processed for IF. The cells were stained with the anti-AR antibody and then with AlexaFluor 588 secondary antibody. Nuclear staining was done by DAPI staining. Image merging of AR and DAPI was analyzed with a Zeiss fluorescence microscope program.AR, androgen receptor; TP, testosterone propionate; 5AR2, 5α-reductase type2; PSA, prostate specific antigen; ADL, A. distichum leaf; DW, distilled water; EtOH, ethanol; Fi, finasteride; IF, immunofluorescence; ADLD, A. distichum leaf distilled water extract; ADLE, A. distichum leaf ethanol extract; ADLH, A. distichum leaf hexane extract; DAPI, 4′,6-diamidino-2-phenylindole.
##P < 0.01 compared to untreated LNCaP cell; **P < 0.01, ***P < 0.001 and ****P < 0.0001 vs. TP-treated LNCaP cell.
|
Effect of ADLE on the inhibition of AR signaling according to harvest season
The activity of ADLE was compared according to the harvest time. As a result of the previous experiment, the 70% ethanol extract had the highest androgen signal inhibitory ability. Accordingly, the activity was compared by extracting the leaves harvested in spring and autumn with 70% ethanol. ADLE extracted from the autumn leaves showed a higher inhibitory effect on PSA and 5AR2 than the ADLE extracts harvested in spring (Fig. 2).
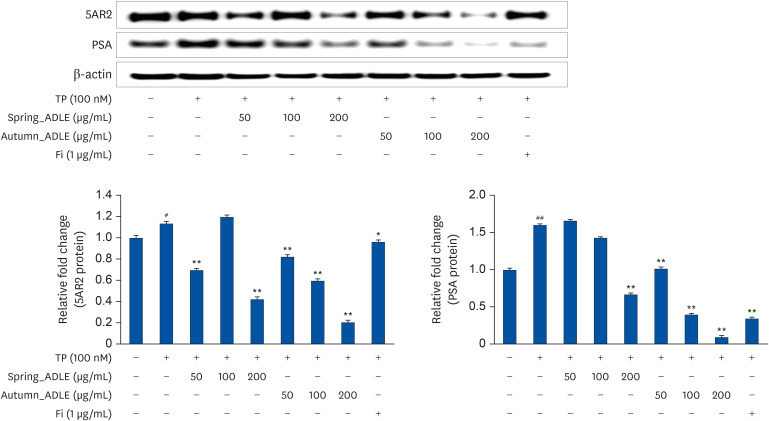 | Fig. 2Effect of ADLE on inhibiting androgen receptor signaling according to harvest season. LNCaP cells were incubated for 24 h in culture medium containing TP (100 nmol), spring_ADLE (50, 100, or 200 μg/mL), autumn_ADLE (50, 100, or 200 μg/mL) or Fi (1 μg/mL). Thereafter, cell lysates (30 μg) were analyzed for the expression of 5AR2 and PSA. The relative protein expression levels were normalized to those of β-actin.ADLE, A. distichum leaf ethanol extract; TP, testosterone propionate; Fi, finasteride; 5AR2, 5α-reductase type2; PSA, prostate specific antigen.
#P < 0.05 and ##P < 0.01 vs. untreated cells; *P < 0.05 and **P < 0.01 vs. TP-treated cells.
|
Effect of ADLE on prostate tissue weight in BPH rats
In the previous experiment, the 70% ethanol extract of A. distichum leaves harvested in autumn inhibited androgen signaling, a key signal of BPH in LNCaP cells. Therefore, the BPH alleviation efficacy of ADLE was evaluated using a TP-induced BPH model. The BPH group showed a significantly higher prostate weight compared to the Con group (Fig. 3B). The BPH group had a 1.62 times larger prostate index than the Con group (Fig. 3C). In contrast, the BPH + ADLE group decreased the prostate index significantly compared to the BPH group. In addition, the BPH + ADLE group showed an interesting result because it showed a superior prostate size-reduction effect than Saw, which is currently sold as a functional food for BPH, and showed similar efficacy to the currently used Fi. The prostate was fixed using 10% formalin, and a histological examination was then performed with paraffin. In the group in which prostatic hyperplasia was induced, the thickness of the prostate tissue epithelial cells became thicker. The BPH group had significantly thicker prostate epithelial cells than the Con group (Fig. 3D and E). The BPH + ADLE group showed a significant reduction in the thickness of epithelial cells compared to the BPH group.
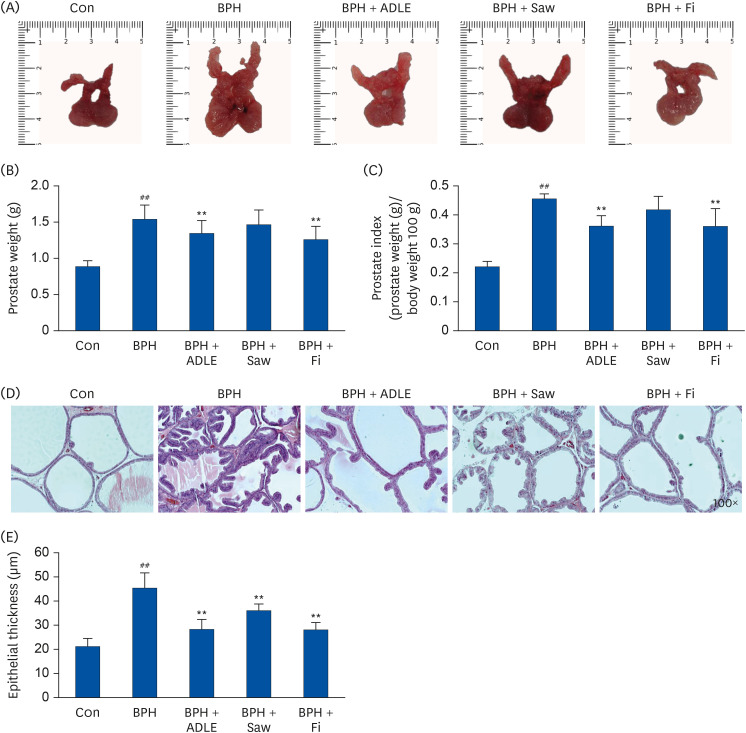 | Fig. 3Effect of ADLE on prostate tissue in TP-induced BPH rats. (A) Photographs of the prostate tissues (VP, DLP, AP). (B) Total prostate tissue weight and (C) prostate indices of the rats. (D) H&E-stained prostate tissues (magnification 100×). (E) Epithelial thickness of the prostate tissues.ADLE, A. distichum leaf ethanol extract; TP, testosterone propionate; BPH, benign prostatic hyperplasia; VP, ventral prostate; DLP, dorsolateral prostate; AP, anterior prostate; Fi, finasteride; H&E, hematoxylin and eosin; DW, distilled water; Con, corn oil subcutaneous injection and DW oral intake; BPH, TP subcutaneous injection (3 mg/kg) and oral intake; BPH + ADLE, TP subcutaneous injection (3 mg/kg) and A. distichum leaves 70% ethanol extract (100 mg/kg) oral intake; BPH + Saw, TP subcutaneous injection (3 mg/kg) and saw palmetto extract (100 mg/kg) oral intake; BPH + Fi, subcutaneous injection and finasteride (1 mg/kg) oral intake.
##P < 0.01 vs. the Con group; **P < 0.01 vs. the BPH group (n = 8 per group).
|
Effect of ADLE on AR-signaling-related factors in prostate tissues of BPH rats
Fig. 4A shows the expression of 5AR-2, AR, ER, SRC-1, and PSA by Western blotting analysis. The protein levels of 5AR-2, AR, SRC-1, and PSA were higher in the BPH group (P < 0.01) than the control. On the other hand, the expression of the 5AR-2, AR, SRC-1, and PSA proteins in the BPH + ADLE group was significantly downregulated compared to the BPH group (Fig. 4A and B). In addition, the BPH group had an increased DHT level compared to the Con group, and the BPH + ADLE group and Saw or Fi group had significantly lower DHT levels than the BPH group (Fig. 4C). IHC was performed to confirm the expression of AR in the tissue. AR was overexpressed in the BPH group compared to the Con group. On the other hand, the BPH + ADLE group showed significantly reduced AR expression than the BPH group (Fig. 4D). Overall ADLE had an inhibitory effect on AR signaling in the BPH rat model.
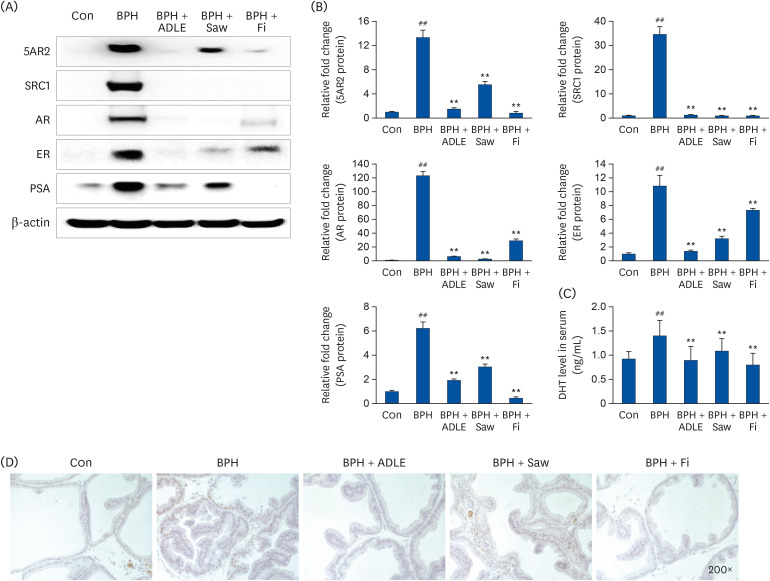 | Fig. 4Effect of ADLE on the AR-signaling-related factors in prostate tissues of BPH rats. (A) Representative western blot showing the bands of 5AR2, SRC1, AR, ER, and PSA. (B) densitometer of the protein expression level. (C) Concentration of DHT in the serum. (D) Immunochemical staining of AR in prostate tissue.ADLE, A. distichum leaf ethanol extract; TP, testosterone propionate; AR, androgen receptor; BPH, benign prostatic hyperplasia; 5AR2, 5α-reductase type2; SRC1, steroid receptor co-activator 1; ER, estrogen receptor; PSA, prostate specific antigen; DHT, dihydrotestosterone; DW, distilled water; Con, corn oil subcutaneous injection and DW oral intake; BPH, TP subcutaneous injection (3 mg/kg) and oral intake; BPH + ADLE, TP subcutaneous injection (3 mg/kg) and A. distichum leaves 70% ethanol extract (100 mg/kg) oral intake; BPH + Saw, TP subcutaneous injection (3 mg/kg) and saw palmetto extract (100 mg/kg) oral intake; BPH + Fi, subcutaneous injection and finasteride (1 mg/kg) oral intake.
##P < 0.01 vs. the Con group; **P < 0.01 vs. the BPH group (n = 8 per group).
|
Effect of ADLE on the growth factors and PI3K/AKT pathway in prostate tissues of BPH rats
BPH increases the expression of growth factors caused by overexpression of the androgen signal. Through this experiment, the level of growth factor expression of BPH and the ability of ADLE to inhibit growth factor expression was confirmed. The protein levels of EGF, IGF-1, TGF-β1, and VEGF in the prostate tissue were measured by Western blotting (Fig. 5A). The BPH group showed a significant increase in growth factor protein levels than the Con group (P < 0.01). On the other hand, the BPH + ADLE group (P < 0.01) showed significantly reduced levels of these proteins compared to the BPH group. Increasing the growth factor leads to PI3K/AKT overexpression, which increases prostate cell proliferation and decreases apoptosis, resulting in prostate growth. PI3K and p-AKT expression were significantly higher in the BPH group than in the Con group. The administration of ADLE reduced the expression of PI3K and p-AKT significantly compared to the BPH group (Fig. 5B).
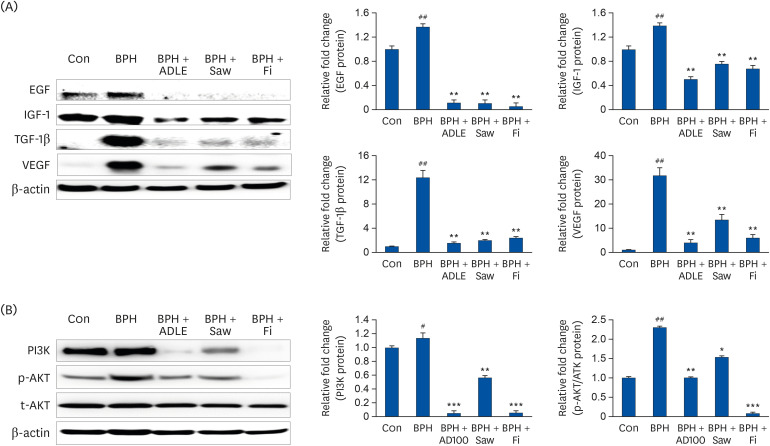 | Fig. 5Expression of growth factors and PI3K/AKT in prostate tissue after the administration of ADLE in BPH. (A) Representative western blot showing the bands of EGF, IGF-1, TGF-β1, and VEGF. (B) Representative western blot showing the bands of PI3K, p-AKT and t-AKT.TP, testosterone propionate; ADLE, A. distichum leaf ethanol extract; BPH, benign prostatic hyperplasia; EGF, epidermal growth factor; IGF-1, insulin-like growth factor-1; VEGF, vascular endothelial growth factor; TGF-β1, transforming growth factor-beta 1; DW, distilled water; Con, corn oil subcutaneous injection and DW oral intake; BPH, TP subcutaneous injection (3 mg/kg) and oral intake; BPH + ADLE, TP subcutaneous injection (3 mg/kg) and A. distichum leaves 70% ethanol extract (100 mg/kg) oral intake; BPH + Saw, TP subcutaneous injection (3 mg/kg) and saw palmetto extract (100 mg/kg) oral intake; BPH + Fi, subcutaneous injection and finasteride (1 mg/kg) oral intake.
#P < 0.05 and ##P < 0.01 vs. the Con group; *P < 0.05, **P < 0.01 and ***P < 0.001 vs. the BPH group (n = 8 per group).
|
Effect of ADLE on the proliferation and apoptosis-related factor in prostate tissues of BPH rats
In BPH, PCNA and cyclin D1, which are critical factors related to prostate cell proliferation, are substantially overexpressed. As a result of the experiment, PCNA and cyclin D1 were overexpressed in the BPH group compared to the Con group (Fig. 6A). In addition, the administration of ADLE produced a significant reduction effect compared to the BPH group. IHC confirmed that the BPH group showed increased PCNA expression than the Con group (Fig. 6B). The administration of ADLE inhibited prostate cell proliferation by reducing the overexpressed PCNA.
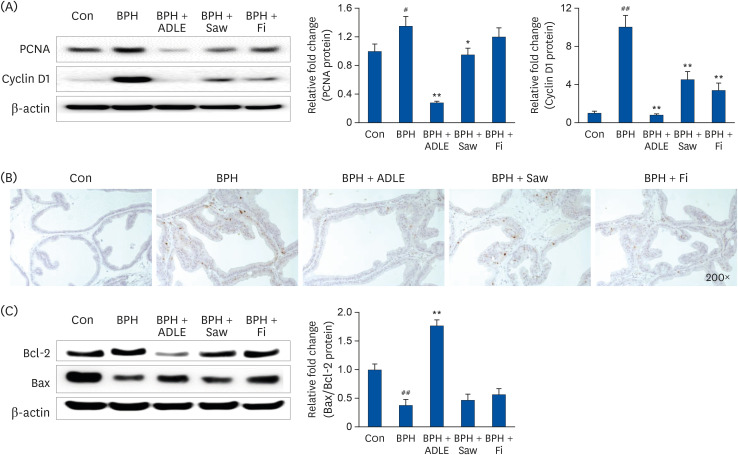 | Fig. 6Expression of proliferation and apoptosis-related factor in the prostate tissue after the administration of ADLE in BPH. (A) Representative western blot showing the bands of PCNA and cyclin D1. (B) Immunochemical staining of AR in prostate tissue. (C) Representative western blot showing the bands of Bcl-2 and Bax.ADLE, A. distichum leaf ethanol extract; BPH, benign prostatic hyperplasia; PCNA, proliferative cell nuclear antigen; AR, androgen receptor; DW, distilled water; Con, corn oil subcutaneous injection and DW oral intake; BPH, TP subcutaneous injection (3 mg/kg) and oral intake; BPH + ADLE, TP subcutaneous injection (3 mg/kg) and A. distichum leaves 70% ethanol extract (100 mg/kg) oral intake; BPH + Saw, TP subcutaneous injection (3 mg/kg) and saw palmetto extract (100 mg/kg) oral intake; BPH + Fi, subcutaneous injection and finasteride (1 mg/kg) oral intake.
#P < 0.05 and ##P < 0.01 vs. the Con group; *P< 0.05 and **P < 0.01 vs. the BPH group (n = 8 per group).
|
The apoptosis of prostate epithelial cells will occur more frequently than in the prostate from a normal state BPH. Bcl-2 to suppress apoptosis is detected in the prostate epithelium. This leads to prostate growth. Although the expression of the anti-apoptotic protein Bcl-2 decreased, the level of the pro-apoptotic protein Bax increased in the BPH + ADLE group. The BPH + ADLE group showed a significantly reduced Bax/Bcl-2 ratio than the BPH group (Fig. 6C).
Go to :
DISCUSSION
The prostate is a representative organ that relies on male hormones to maintain proliferation and function. Therefore, testosterone and DHT play an essential role in maintaining prostate homeostasis [36]. DHT binds to AR, a member of the nuclear receptor family, and acts biologically as a ligand-responsive transducer [37]. AR co-activators, such as SRC-1, interact with AR to regulate AR transactivation [38]. On the other hand, in men with a hormonal imbalance due to decreased testosterone levels caused by aging, the testosterone levels decreased, but the levels of 5AR2 and AR expression increased to regulate endocrine homeostasis and maintain a constant level of DHT in the prostate [39]. As a result, the combination of DHT and AR increases, which can lead to an enlarged prostate. 5ARI, a currently used treatment for prostatic hyperplasia, blocks 5AR2 and reduces the level of DHT in the prostate, thereby inhibiting excessive prostate growth [40]. On the other hand, several side effects have been reported, and replacements are urgently needed. As a result, research is actively underway on alternative substances derived from natural products that are safe for consumption and have no side effects. A literature review showed that A. distichum contains a large amount of flavonoids and polyphenols. There are numerous reports that several flavonoids and polyphenols may help relieve BPH [414243]. Therefore, this study examined whether the A. distichum were subjected to water extraction, 70% ethanol extraction, and 95% hexane extraction and then treated in LNCaP cells to see if they inhibit the expression of androgen-signaling related factors. As a result, ADLE inhibited the expression of AR, as shown in the western blot and IF results (Fig. 1). Similar to the study published by Wu et al. [44], in this experiment, Fi inhibited the binding and activity of DHT and AR without inducing a decrease in the amount of AR protein. Therefore, Fi did not alter the amount of AR protein, but ADLE exhibited efficacy in reducing the amount of AR itself in prostate cells (Fig. 1). The mechanism through which ADLE reduced AR protein expression cannot be known only from the current study. Hence, further studies will be needed.
In the case of Forsythia koreana, which is similar to A. distichum, the metabolite content differs according to the season. Hence, A. distichum leaves harvested in spring or autumn were extracted with 70% ethanol. The LNCaP cells were then treated with the extract. The western blot experiments showed that the A. distichum leaves harvested in autumn inhibited the expression of 5AR2 and PSA significantly compared to A. distichum leaves harvested in spring (Fig. 2). Therefore, ADLE harvested in autumn was used for the later TP-induced BPH rat model. The TP treatment-induced prostatic hyperplasia model is a commonly used BPH model because it shows similar trends clinically to patients with prostatic hyperplasia. The BPH-inducing group was castrated before the TP treatment to block endogenous testosterone. The BPH group showed a significantly higher prostate weight than the Con group. Compared to previously published papers, the weight of the prostate increased to a similar level in this experiment [35]. The ADLE treatment group showed a significantly lower prostate weight than the BPH group, highlighting its efficacy in alleviating prostatic hyperplasia. Histological analysis showed that the thickness of epithelial cells in the ADLE group was significantly lower than that of the BPH group. ADLE alleviated TP-induced prostate overgrowth in phenomenological and histological analysis. In addition, the effect of ADLE on the molecular biological mechanisms in prostatic hyperplasia was investigated. An analysis of the androgen signaling-related factors, which are essential for the development of prostatic hyperplasia, by Western blot analysis, showed that ADLE inhibited the expression of the androgen signaling-related factors. AR is a DNA-binding transcription factor that binds DHT and regulates AR target gene expression, and is critical for prostate growth. AR signaling acts as a key factor promoting the growth of stromal cells and epithelial cells in BPH, contributing to the development of BPH [7]. ADLE effectively reduced the expression of the AR signaling-related factors. By reducing the expression of 5AR2, it showed similar efficacy to Fi, confirming that it has the function of 5ARI. An analysis of AR expression in the tissues through Western blot and IHC staining showed that ADLE suppressed the excessively increased AR expression in the BPH group (Fig. 4). ADLE decreased the expression of SRC-1, a major factor regulating the binding of DHT and AR, and PSA, a representative AR signaling target gene (Fig. 4A). Therefore, ADLE inhibits the binding of AR and DHT, suggesting a pharmacological effect of alleviating the development of BPH.
AR signaling contributes to BPH development by increasing the expression of growth factors, such as EGF, IGF, and VEGF [45]. When AR signaling is decreased by ADLE, it is expected that growth factor expression in the prostate will also be down-regulated, which was confirmed. As a result, ADLE effectively reduced the growth factor in the prostate (Fig. 5A). This can explain why ADLE shows efficacy of inhibiting AR signaling. The PI3K/AKT pathway is expressed by growth factors that determine cell proliferation and apoptosis. In addition, the extract of A. distichum also suppresses overexpressed PI3K/AKT, thereby controlling cell death and cell proliferation [31]. BPH develops due to the indiscriminate proliferation of stromal cells and epithelial cells caused by AR signaling [46]. This suggests that the expression of proliferative factors is increased, and several studies show that BPH overexpresses PCNA and cyclin D1. PCNA acts as an essential DNA clamp for replication and is used extensively to assess cell proliferation in conjunction with the benign cell growth regulator cyclin [4748]. These results showed that ADLE had an antiproliferative effect on BPH by decreasing the expression of cyclin D1 and PCNA. ADLE then showed the effect of increasing apoptosis in BPH. In this experiment, the expression of Bax and Bcl-2, which are key factors determining apoptosis in the endogenous apoptosis pathway, was confirmed. An examination of the Bax/Bcl-2 ratio confirmed that ADLE induces apoptosis in prostate cells. Therefore, ADLE normally regulates the proliferation and death of prostate cells by suppressing the expression of excessively increased AR expression in BPH, demonstrating its potential as a therapeutic agent for BPH.
This experiment showed that ADLE had an alleviating effect on BPH by regulating the ARs in BPH. According to published research, A. distichum leaves contain rutin, taxifolin, narirutin, isorhamnetin, gallic acid, chlorogenic acid, and verbascoside. According to several studies, verbascoside is treated as an indicator of A. distichum [49]. Studies have shown that verbascoside effectively alleviates BPH by promoting the apoptosis of prostate cells in BPH-induced rats. In addition, in a previously reported study, chlorogenic acid reduced significantly the prostate index and serum 5AR2 levels in TP-induced BPH mice [50]. On the other hand, it is unclear which functional component in A. distichum affected the downregulation of AR signaling in BPH. Hence, further study will be needed. In the future, it will be necessary to use UPLC to identify metabolites in ADLE and then treat prostate cells to select active substances.
Go to :
 XML Download
XML Download