

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Gastrointestinal organs and associated visceral organs, such as the pancreas, liver, and adipose tissue control energy balance (i.e., the balance between energy intake and energy expenditure) by regulating satiety or hunger signals. They communicate with the hypothalamus, a key appetite control center, through neural and endocrine pathways [1]. Neurons in the hypothalamus express appetite-stimulating or suppressing neuropeptides in response to signals from the periphery to maintain energy homeostasis. Dysregulation of these systems results in chronic positive energy balance, thus leading to obesity [2] and the development of chronic diseases such as dyslipidemia, hypertension, type 2 diabetes, and cardiovascular diseases [3]. Therefore, dietary bioactive compounds capable of improving the regulation of energy homeostasis could be used as effective and safe nutraceutical strategies to prevent obesity and obesity-related chronic diseases.
Pine nuts have been consumed for centuries around the world, especially in the Mediterranean and Asia, and approximately 60% of the weight of pine nuts is fat. The fatty acid composition of Korean pine nut (Pinus Koraiensis) oil (PNO) is 4% palmitic acid (16:0), 28% oleic acid (18:1, Δ9), 47% linoleic acid (18:2, Δ9,12), and 14% pinolenic acid (18:3, Δ5,9,12) [4]. Pinolenic acid, a unique fatty acid found in PNO, is an unsaturated polymethylene-interrupted fatty acid with a cis-5 ethylenic bond (Δ5-UPIFA) [5] and a positional isomer of γ-linolenic acid [6]. Korean PNO contains a much higher fraction of pinolenic acid than other PNOs including Italian stone PNO [7].
Many studies have confirmed the health benefits of Korean PNO. In a mouse model of diet-induced obesity, Korean PNO suppressed intestinal lipid uptake [8], increased fatty acid oxidation in skeletal muscle [9], and induced thermogenesis in brown adipose tissue (BAT) [9]. Korean PNO was also shown to attenuate high-fat diet (HFD)-induced hepatic lipid accumulation in mice [10], and blood pressure in spontaneously hypertensive rats [11]. In clinical studies, Korean PNO increased the release of satiety hormones and decreased appetite in overweight post-menopausal women [7] and decreased dietary intake in overweight women [12]. However, another study reported no effect of Korean PNO on appetite and energy intake [13]. Moreover, no previous studies have evaluated the chronic effects of Korean PNO on the expression of satiety hormone receptors and neuropeptides, which play a significant role in food intake, energy expenditure, and weight control [2]. Therefore, our study sought to examine the effects of Korean PNO on the long-term regulation of energy balance, including satiety hormones, neuropeptides, dietary lipid absorption, and adiposity, in mice fed with a HFD (45% kcal fat) or control diet (10% kcal fat) for 12 weeks.
Go to : 
MATERIALS AND METHODS
Animals and diets
Five-week-old male C57BL/6N mice were purchased from Central Laboratory Animal Inc. (Seoul, Korea) and maintained on a chow diet for 3 days before being assigned to 1 of 4 experimental diets: control diets containing 10% kcal fat from Korean PNO or soybean oil (SBO) (PC or SC) or HFDs containing 35% kcal fat from lard and 10% kcal fat from Korean PNO or SBO (PHFD or SHFD). The mice (n = 10-11 per group) were fed with the experimental diets ad libitum for 12 weeks. Table 1 shows the composition of the experimental diets. Korean PNO, generously provided by Dubio Co., Ltd. (Hwaseong, Korea), was sent to Dyets Inc. (Bethlehem, PA, USA) to prepare the experimental diets. All mice were housed individually under controlled temperature (23 ± 3°C) and humidity (55 ± 10%) conditions with a 12 hour-dark/light cycle. Body-weight was measured once a week, and food intake was measured 4 times a week. At the end of the experimental period, the mice were euthanized in a CO2 chamber after a 12-h fast. Brain, stomach, small intestine, and white adipose tissues (WAT) were dissected. All tissues were cleaned, flash-frozen in liquid nitrogen, and stored at −80°C until required for analyses. All animal procedures were approved and carried out in accordance with the Institutional Animal Care and Use Committee of Seoul National University (approval No. SNU-101029-1).
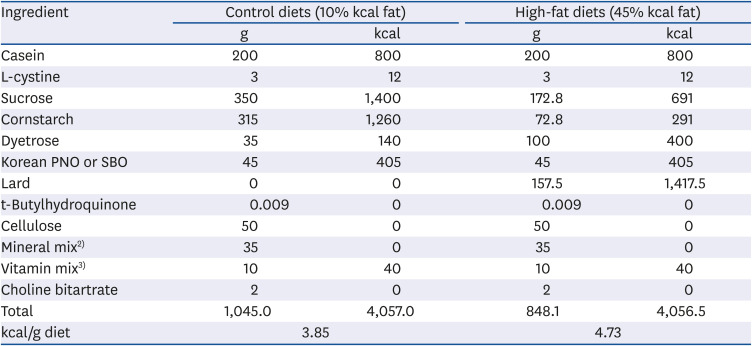
Table 1
Composition of the experimental diet1)
PNO, pine nut oil; SBO, soybean oil.
1)Resource: Dyets, Inc., Bethlehem, PA, USA.
2)Thirty-five g of mineral mix (Research Diets, Inc., New Brunswick, NJ, USA, S10026) provides 1.0 g sodium, 1.6 g chloride, 0.5 g magnesium, 0.33 g sulfur, 59 mg manganese, 45 mg iron, 29 mg zinc, 6 mg copper, 2 mg chromium, 1.6 mg molybdenum, 0.16 mg selenium, 0.9 mg fluoride, 0.2 mg, iodine and 3.99 g sucrose.
3)Ten g of vitamin mix (Research Diets, Inc., V10001) provides 4000 IU vitamin A, 1000 IU vitamin D3, 50 IU vitamin E, 30 mg niacin, 16 mg pantothenic acid, 7 mg vitamin B6, 6 mg vitamin B1, 6 mg vitamin B2, 2 mg folic acid, 0.5 mg menadione, 0.2 mg biotin, 10 μg vitamin B12 and 9.78 g sucrose.
![]()
Determination of fecal triacylglycerol (TAG), non-esterified fatty acids (NEFAs), and cholesterol concentrations
Feces were collected for 3 days during the 12th week of the feeding experiment, and fecal TAG, NEFA, and cholesterol concentrations were determined via an enzymatic assay using a commercial kit (Asan Pharmacy, Seoul, Korea) following the manufacturer's instructions.
RNA extraction and complementary DNA (cDNA) synthesis
Total RNA was extracted from the hypothalamus and the remaining brain tissue, as well as from the stomach, jejunum, and epididymal WAT using Trizol reagent (Invitrogen, Carlsbad, CA, USA). cDNA was synthesized from 2 μg of total RNA using the PrimeScript II 1st strand cDNA synthesis kit (Takara, Kusatsu, Japan).
Quantification of gene expression
To determine the effects of PNO on the appetite control pathway, the mRNA levels of cholecystokinin related genes (jejunal Cck, and cerebral Cckar and Cckbr), ghrelin related genes (gastric Ghrl and Goat, and hypothalamic Ghsr), and neuropeptides (hypothalamic Npy, Agrp, Pomc, and Cartpt) were quantified. To examine the influence of PNO on lipid metabolism, the mRNA levels of genes associated with lipid absorption and metabolism in the small intestine (jejunal Cd36, Fabp2, Dgat2, and Apoa4) and those in WAT (epididymal Lpl, Plin1, Ucp2, Adrb3, Pparg, Ppargc1a, and Ppard) were quantified via real-time polymerase chain reaction (PCR) using SYBR Premix Ex Taq (Takara) and a StepOne Real-time PCR System (Applied Biosystems, Foster City, CA, USA). Each reaction was performed in duplicate. The results were analyzed with the StepOne™ software version 2.1 (Applied Biosystems) and all values were normalized to the levels of the house-keeping gene Gapdh. Table 2 summarizes the specific primer sequences used to conduct the aforementioned experiments.
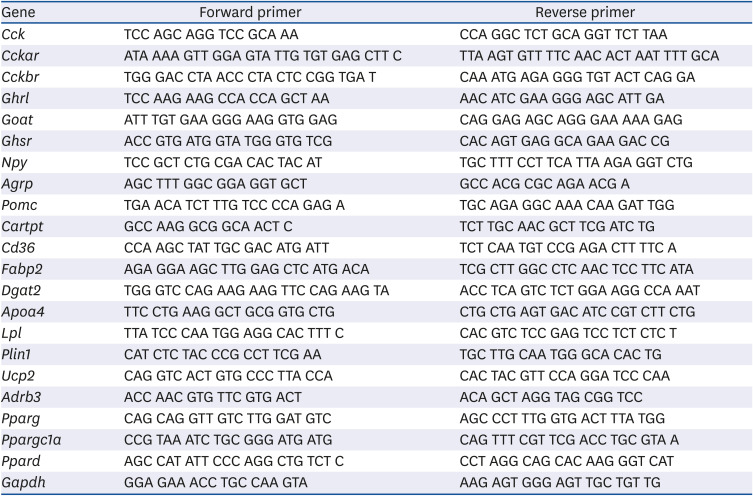
Table 2
The primer sequences used for real-time polymerase chain reaction
Cck, cholecystokinin; Cckar, cholecystokinin A receptor; Cckbr, cholecystokinin B receptor; Ghrl, ghrelin; Goat, ghrelin O-acyltransferase; Ghsr, growth hormone secretagogue receptor; Npy, neuropeptide Y; Agrp, agouti-related peptide; Pomc, pro-opiomelanocortin-alpha; Cartpt, cocaine- and amphetamine-regulated transcript prepropeptide; Cd36, fatty acid translocase; Fabp2, fatty acid binding protein 2, intestinal; Dgat2, diacylglycerol O-acyltransferase 2; Apoa4, apolipoprotein A-IV; Lpl, lipoprotein lipase; Plin1, perilipin 1; Ucp2, mitochondrial uncoupling protein 2; Adrb3, beta-3 adrenergic receptor; Pparg, peroxisome proliferator-activated receptor gamma; Ppargc1a, peroxisome proliferator-activated receptor gamma coactivator 1 alpha; Ppard, peroxisome proliferator-activated receptor delta; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
![]()
Statistical analysis
All statistical analyses were conducted using SPSS version 19.0 (SPSS Inc., Chicago, IL, USA). Two-way analysis of variance was conducted to identify the overall effects of dietary fat amount, oil type, and the interaction between these 2 factors, followed by Fisher's LSD multiple comparison test for individual group comparisons. Pearson's correlation was used to determine the linear relationship between variables. Differences were considered statistically significant at P < 0.05.
Go to :
RESULTS
Body weight change, food intake, feed efficiency, and WAT weight
HFD-fed mice exhibited significantly higher weight gain (P < 0.001) and WAT amount (P < 0.001) than the mice fed with the control diet. Mice fed with diets containing Korean PNO gained less weight gain (P = 0.014) and exhibited less white fat mass (P = 0.004) than mice fed with the SBO-containing diets. There were no significant differences in daily food intake and daily energy intake between the mice fed with the diets containing SBO or PNO; however, feed efficiency, a ratio of weight gain to food intake, was lower (P = 0.007) in the mice fed with the PNO-containing diets (Table 3).
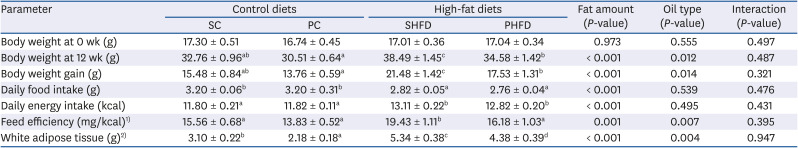
Table 3
Body weight, weight gain, food intake, feed efficiency, white adipose tissue weight, and serum leptin level
Data are presented as means ± SEM, n = 10–11 for each group. Different superscripts indicate significant differences, P < 0.05.
SC, 10% soybean oil; PC, 10% pine nut oil; SHFD, 10% soybean oil + 35% lard; PHFD, 10% pine nut oil + 35% lard; WAT, white adipose tissue.
1)Feed efficiency (mg/kcal) = weight gain (mg)/total food intake (kcal).
2)WAT includes inguinal subcutaneous, epididymal, perirenal, and retroperitoneal WAT.
![]()
The PHFD group exhibited significantly lower body weight (10% less, P = 0.022), less weight gain (18% less, P = 0.015), and lower feed efficiency (17% less, P = 0.012) than the SHFD group. The PC and PHFD groups had significantly less WAT compared to the SC and SHFD groups (30% less, P = 0.046; 18% less, P = 0.033; respectively). Some of the results in Table 3 have been reported in our previous studies (10, 14), in which the effects of Korean PNO on immune response and hepatic triglyceride lipid accumulation were investigated using the same animals.
Fecal lipid concentrations
HFD-fed mice had higher fecal TAG (6.8 times higher, P < 0.001) and NEFA excretion (3.2 times higher, P < 0.001) than control diet-fed mice, whereas cholesterol levels did not differ. Further, fecal NEFA levels were significantly higher in PHFD-fed mice than SHFD-fed mice (58% higher, P = 0.044; Fig. 1).
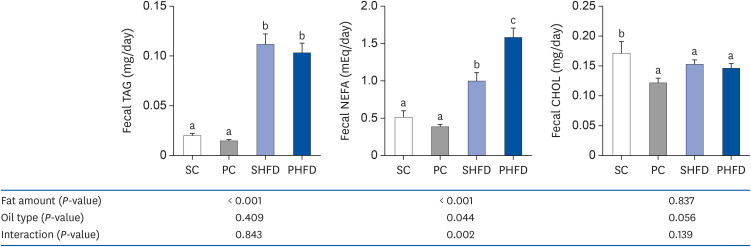 | Fig. 1Fecal triacylglycerol, non-esterified fatty acid, and cholesterol levels. Data are presented as means ± SEM, n= 7–14 for each group. Two-way analysis of variance was used to determine the significant effect of fat amount and oil type. Different letters indicate significant difference at P < 0.05 by Fisher's LSD multiple comparison test.SC, 10% soybean oil; PC, 10% pine nut oil; SHFD, 10% soybean oil + 35% lard; PHFD, 10% pine nut oil + 35% lard; TAG, triacylglycerol; NEFA, non-esterified fatty acid; CHOL, cholesterol.
|
Expression of genes involved in appetite control
The expression of hypothalamic Ghsr, a ghrelin receptor, was significantly affected by both fat amount and oil type (P = 0.028, P = 0.027, respectively). The Ghsr mRNA level of the PC group was 1.23-fold higher (P = 0.024) than that of the SC group. Ghsr mRNA levels showed negative correlation with body weight (r = −0.467, P = 0.025) and white fat mass (r = −0.460, P = 0.027) at 12 week. However, the mRNA levels of Cck (a major satiating hormone) and its receptors Cckar and Cckbr in the brain, gastric Ghrl, (an appetite inducer), and gastric Goat (ghrelin O-acyltransferase; a ghrelin activating enzyme) were not influenced by fat amount or oil type (Table 4), and none of them showed significant correlation with body mass or fat mass (data not shown). The Ghrl mRNA level was not correlated with hypothalamic Npy or hypothalamic Agrp mRNA levels.
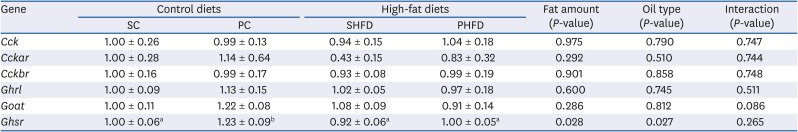
Table 4
The mRNA expression levels of cholecystokinin- and ghrelin-related genes
Data are presented as means ± SEM, n = 5–6 for each group. All values are normalized to the levels of house-keeping gene Gapdh and expressed as relative mRNA level compared to the average expression level of SC group. Different superscripts indicate significant differences, P < 0.05.
mRNA, messenger RNA; SC, 10% soybean oil; PC, 10% pine nut oil; SHFD, 10% soybean oil + 35% lard; PHFD, 10% pine nut oil + 35% lard; Cck, cholecystokinin; Cckar, cholecystokinin A receptor; Cckbr, cholecystokinin B receptor; Ghrl, ghrelin; Goat, ghrelin O-acyltransferase; Ghsr, growth hormone secretagogue receptor; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
![]()
The mRNA level of Agrp, a neuropeptide that increases appetite and reduces energy expenditure, was significantly higher in mice fed with Korean PNO-containing diets (P = 0.022). The PHFD group had significantly higher Agrp expression (2.16-fold, P = 0.023) than the SHFD group. However, the mRNA level of Npy, another neuropeptide that acts similarly to Agrp, was not significantly influenced by fat amount or by oil type. The mRNA levels of hypothalamic Pomc and Cartpt, leptin-responsive neuropeptides that suppress appetite and increase energy expenditure, tended to be higher in mice fed with Korean PNO-containing diets (Pomc, P = 0.079; Cartpt, P = 0.056) (Fig. 2). Further, Npy and Agrp mRNA levels were negatively correlated with the weight (Npy, r = −0.420, P = 0.046; Agrp, r = −0.465, P = 0.025), whereas Pomc and Cartpt mRNA levels showed no correlation with the weight at 12 week (Fig. 3).
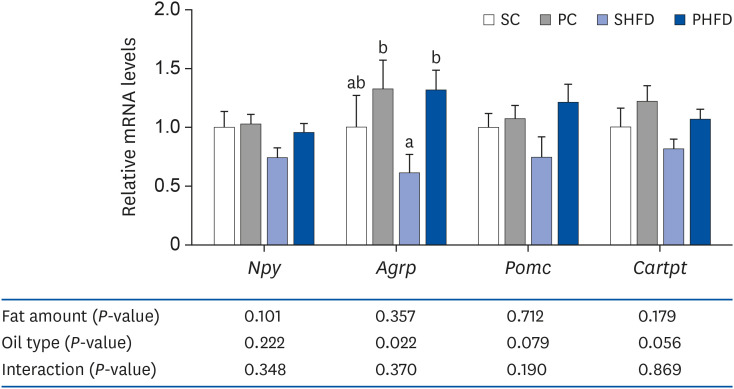 | Fig. 2The mRNA expression levels of neuropeptides (hypothalamic Npy, Agrp, Pomc, and Cartpt). Data are presented as means ± SEM, n= 5-6 for each group. Two-way analysis of variance was used to determine the significant effect of fat amount and oil type. Different letters indicate significant difference at P < 0.05 by Fisher's LSD multiple comparison test. All values are normalized to the levels of house-keeping gene Gapdh and expressed as relative mRNA level compared to the average expression level of SC group.mRNA, messenger RNA; SC, 10% soybean oil; PC, 10% pine nut oil; SHFD, 10% soybean oil + 35% lard; PHFD, 10% pine nut oil + 35% lard; Npy, neuropeptide Y; Agrp, agouti-related peptide; Pomc, pro-opiomelanocortin-alpha; Cartpt, cocaine- and amphetamine-regulated transcript prepropeptide; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
|
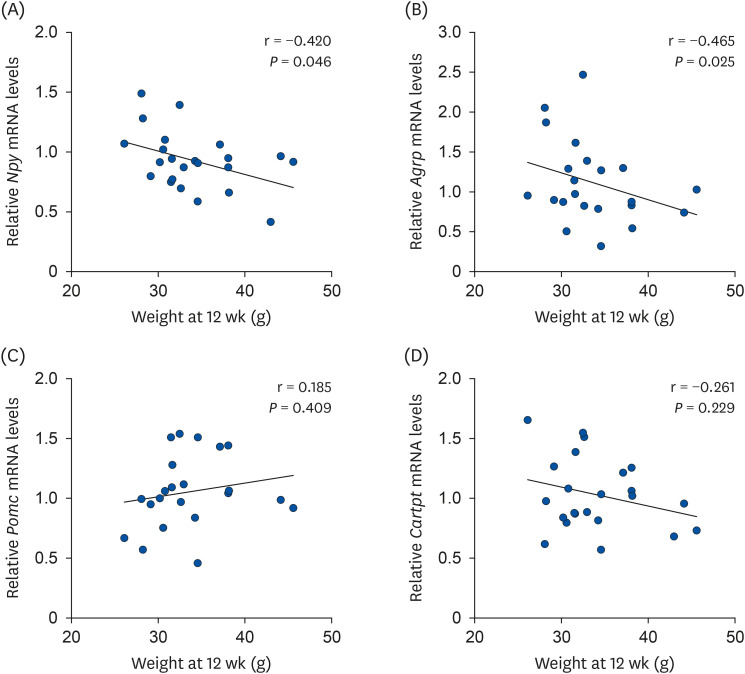 | Fig. 3Correlation between body weight and hypothalamic neuropeptide mRNA levels. Correlation between body weight and (A) Npy, (B) Agrp, (C) Pomc, and (D) Cartpt mRNA levels. Pearson's correlation was used to determine the linear relationship between variables.mRNA, messenger RNA; Npy, neuropeptide Y; Agrp, agouti-related peptide; Pomc, pro-opiomelanocortin-alpha; Cartpt, cocaine- and amphetamine-regulated transcript prepropeptide.
|
Expression of genes involved in lipid absorption
To investigate if the lower body mass, fat mass, and feed efficiency in Korean PNO-fed mice were caused by an alteration in intestinal fat absorption, the expression of genes involved in jejunal lipid metabolism was quantified (Fig. 4).
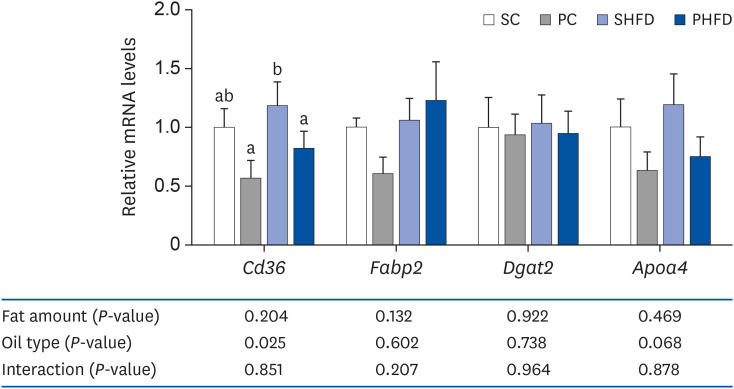 | Fig. 4The mRNA expression levels of genes associated with intestinal lipid metabolism (jejunal Cd36, Fabp2, Dgat2, and Apoa4). Data are presented as means ± SEM, n= 5–6 for each group. Two-way analysis of variance was used to determine the significant effect of fat amount and oil type. Different letters indicate significant difference, P < 0.05. All values are normalized to the levels of house-keeping gene Gapdh and expressed as relative mRNA level compared to the average expression level of SC group.mRNA, messenger RNA; SC, 10% soybean oil; PC, 10% pine nut oil; SHFD, 10% soybean oil + 35% lard; PHFD, 10% pine nut oil + 35% lard; Cd36, fatty acid translocase; Fabp2, fatty acid binding protein 2, intestinal; Dgat2, diacylglycerol O-acyltransferase 2; Apoa4, apolipoprotein A-IV; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
|
Overall, mice fed with diets containing Korean PNO had significantly lower mRNA levels of jejunal Cd36, which transports fatty acids from the gut lumen to enterocytes (P = 0.025). The PC group exhibited a lower yet non-significant Cd36 mRNA expression trend (0.57-fold, P = 0.085) compared to the SC group. Mice fed with Korean PNO-containing diets also tended to have lower expression levels of jejunal Apoa4, which facilitates intestinal lipoprotein production (P = 0.068). Fat amount and oil type did not affect the expression of jejunal Fabp2, which transports and metabolizes fatty acids in enterocytes, and jejunal Dgat2, which synthesizes TAG from diacylglycerol (DAG).
Expression of genes involved in body fat accumulation
To examine whether Korean PNO reduced body fat mass by suppressing lipid storage or by enhancing lipid utilization, the expression of genes involved in lipid metabolism was measured in epididymal WAT (Fig. 5).
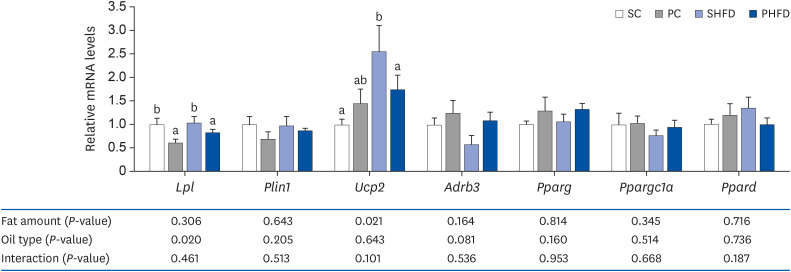 | Fig. 5The mRNA expression levels of genes associated with lipid metabolism in the white adipose tissue (epididymal Lpl, Plin1, Udp2, Adrb3, Pparg, Ppargc1a, and Ppard). Data are presented as means ± SEM, n = 5–6 for each group. Different letters indicate significant difference, P < 0.05. All values are normalized to the levels of house-keeping gene Gapdh and expressed as relative mRNA level compared to the average expression level of SC group.mRNA, messenger RNA; SC, 10% soybean oil; PC, 10% pine nut oil; SHFD, 10% soybean oil + 35% lard; PHFD, 10% pine nut oil + 35% lard; Lpl, lipoprotein lipase; Plin1, perilipin 1; Ucp2, mitochondrial uncoupling protein 2; Adrb3, β3 adrenergic receptor; Pparg, peroxisome proliferator-activated receptor gamma; Ppargc1a, peroxisome proliferator-activated receptor gamma coactivator 1 alpha; Ppard, peroxisome proliferator-activated receptor delta; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
|
Mice fed with Korean PNO-containing diets had significantly lower expression levels of epididymal Lpl (P = 0.020), which hydrolyzes TAG within lipoproteins to DAG and fatty acids to facilitate fatty acid uptake by adipocytes. The Lpl mRNA level of the PC group was 38% lower than that of the SC group (P = 0.035). The mRNA level of epididymal Adrb3, which enhances lipolysis and thermogenesis in WAT, tended to be higher in mice fed with Korean PNO containing diets (P = 0.081). The mRNA level of Ucp2, which plays a role in thermogenesis, was significantly higher in HFD-fed mice (P = 0.021). The mRNA levels of Plin1, Pparg, Ppargc1a, and Ppard were not affected by fat amount and oil type.
Go to :
DISCUSSION
The HFD containing 10% kcal derived from Korean PNO fat and 35% from lard resulted in lower body mass increases and less white fat mass in mice compared to the HFD containing 10% kcal derived from SBO fat and 35% from lard even though the mice exhibited no differences in calorie consumption. Previously, mice fed with a HFD prepared with 30% kcal from Korean PNO fat and 15% from lard reportedly consumed less food (7.27% less g/day, 7.46% fewer calories/day) than those fed with a HFD made with 30% kcal from SBO fat and 15% from lard [14]; however, the substitution of 10% kcal Korean PNO fat for SBO in the control diet or the HFD did not induce any difference in food intake. This indicates that the substitution of 10% kcal fat with Korean PNO may not be enough to exert significant effects on appetite control, and the differences in body fat mass between SBO- or Korean PNO-fed mice may be caused by other characteristics of Korean PNO. Therefore, in this study, we investigated the chronic effects of dietary Korean PNO on molecular mechanisms of energy balance regulation, including the expression of satiety hormone receptors and neuropeptides.
We did not observe differences in the mRNA levels of Cck, a major satiety hormone, and its receptors Cckar and Cckbr among the experimental groups. This appeared to contradict a previous study that reported that Korean PNO promoted CCK release both in STC-1 enterocytes and in post-menopausal overweight women [7]. This discrepancy may have been caused by the 12-hour fasting period to which the mice were submitted prior to their euthanasia in this study. Given that CCK signals are induced when food is present in the gut lumen [15], fasting could have masked the satiating effect of PNO. However, it is also plausible that the expression of satiating peptides was not affected simply because the amounts of dietary Korean PNO were too low, which could also explain why no differences in food intake were observed between the SBO- and Korean PNO-containing diet groups.
The mRNA levels of Ghrl, an appetite-stimulating hormone, and Goat, a ghrelin activating enzyme, were not different among the experimental groups. This result was consistent with earlier studies that reported no difference in Ghrl and Goat expression between lean and diet-induced obese mice after fasting [1617]. However, the mRNA level of ghrelin receptor Ghsr was significantly higher in the PC group than in other groups, and the Ghsr mRNA levels were negatively correlated with body weight and white fat mass (data not shown). Ghsr is known to increase appetite and decrease energy expenditure and is thus upregulated when more energy intake is needed [18]. Therefore, these data suggest that mice in the PC group, which had the lowest body fat amount among all groups, received a strong appetite-stimulating signal to promote energy consumption. Similarly, Korean PNO consumption was reported to mimic calorie restriction responses by reversing HFD-induced downregulation of SIRT3 (sirtuin 3; i.e., a protein that mediates calorie restriction) [10]. In contrast, Ghrl and Goat mRNA levels showed no correlation with body weight and fat mass (data not shown). This may indicate that the expression of gastrointestinal hormones and their associated enzymes is regulated by short-term satiation signals, whereas that of hormone receptors is regulated by long-term energy status signals.
Given that Korean PNO-fed mice had lower amounts of WAT, we speculated that they would also have lower expression levels of Npy and Agrp (neuropeptides that increase energy intake and reduce energy expenditure). However, contrary to our expectations, Agrp mRNA expression was significantly lower in the SHFD group compared to the PHFD group. A compensatory mechanism to decrease food intake and maintain energy balance was likely induced in the SHFD group mice, as they exhibited higher body weight and more WAT than the PHFD mice. This was further supported by the negative correlation of Npy and Agrp mRNA levels with body weight and white fat amount.
Pomc and Cartpt mRNA levels tended to be higher in Korean PNO-fed mice, whereas Pomc and Cartpt levels in the SHFD group were generally lower. Given that leptin stimulates Pomc and Cartpt expression [1], the SHFD group, which had the highest leptin level [10], was expected to have higher Pomc and Cartpt expression than the other groups. However, the SHFD group had lower Pomc and Cartpt mRNA levels, suggesting that leptin failed to induce an increase in Pomc and Cartpt expression. This, in turn, may have been caused by leptin resistance. Similar results were reported in previous animal studies. High-fat and high-sugar diet lowered Pomc mRNA levels in rats [19], and obese rats had lower Pomc expression than obesity-resistant rats [20]. In these 2 studies, Pomc downregulation induced hyperphagia and obesity [1920]. Another study reported that HFD feeding for 8 weeks did not upregulate Pomc mRNA expression, and Pomc mRNA expression in HFD-fed mice was even lower than that of lean mice after 19 weeks. This dysregulation of the POMC/CART pathway was accompanied by severe leptin resistance, suggesting that leptin resistance was a possible cause [21]. Therefore, the lower fat mass of the PHFD group may have prevented the development of leptin resistance and POMC/CART pathway dysregulation.
Korean PNO upregulated fecal NEFA excretion, and fecal NEFA levels were significantly higher in mice fed with the Korean PNO-containing HFD than in mice fed with the SBO-containing HFD. Consistent with these observations, Korean PNO-fed mice exhibited lower Cd36 expression and tended to have lower Apoa4 expression in the jejunum. Dietary fatty acids in the gut lumen are transported by CD36 into enterocytes [22], and Apo A-IV facilitates the chylomicron packaging process [23]. Therefore, the lipids in Korean PNO-containing diets may not be efficiently absorbed and packaged into chylomicron particles compared to SBO-containing diets, which could have led to the lower fat mass and body mass of mice fed with the Korean PNO-containing diets. This was supported by one of our previous studies in which mice were fed with a HFD with 30% kcal fat from Korean PNO and 15% kcal fat from lard. The mice fed with the Korean PNO-containing diet tended to exhibit lower mRNA expression of Mttp, which is involved in TAG incorporation into chylomicrons [8].
Compared to SBO-fed mice, Korean PNO-fed mice had lower Lpl mRNA expression and tended to have higher Adrb3 mRNA expression in epididymal WAT. These data suggest the potential benefits of Korean PNO for attenuating body fat accumulation by downregulating fatty acid uptake by adipocytes and upregulating lipolysis and thermogenesis in adipocytes. This is because LPL (lipoprotein lipase) in adipocytes cleaves fatty acids from TAG in chylomicrons [24], and activation of the β3-adrenergic receptor, encoded by the Adrb3 gene, increases lipolysis and thermogenesis [25]. This is supported by another study in which substitution of 30% kcal fat with Korean PNO in a 45% kcal fat diet upregulated Ucp1 expression in BAT at both the mRNA and protein levels [9]. Pinolenic acid, the key fatty acid in Korean PNO, upregulated thermogenic gene expression, including Ucp1 and Ppargc1a, and augmented the norepinephrine-induced upregulation of the genes in mouse primary adipocytes [26]. The higher Ucp2 mRNA levels in the SHFD group were interpreted as a defense mechanism to maintain energy balance by increasing energy expenditure, which was supported by previous studies in which HFD upregulated Ucp2 expression [2728]. Although 45% kcal fat diets containing 30% kcal fat from Korean PNO upregulated Ppargc1a and Ppard mRNA expression in skeletal muscle [9], the expression of these genes in epididymal WAT was not different among the groups evaluated in the present study. This inconsistency may be due to the differences in Korean PNO content in the experimental diets or the differences in tissue characteristics. For instance, skeletal muscle is richer in mitochondria compared to other tissue types and could thus regulate fatty acid catabolism more sensitively.
In summary, dietary Korean PNO consumption reduced weight gain in HFD-induced obese mice and also decreased the amount of WAT in both control diet- and HFD-fed mice without affecting food intake. Further, the SHFD group exhibited potential POMC/CART pathway dysregulation, whereas the PHFD group did not, suggesting that the appetite-suppressing pathway may not be impaired in the PHFD group. The higher fecal NEFA excretion, the lower Cd36 mRNA expression, and lower Apoa4 mRNA expression tendency in Korean PNO-fed mice indicated that Korean PNO may have lower intestinal lipid absorption efficiency. The lower Lpl mRNA expression and higher Adrb3 mRNA expression trend may suggest that Korean PNO attenuated fat accumulation in WAT by decreasing lipid uptake by adipocytes and increasing lipolysis in adipocytes.
One of the limitations of this study was that expression of gastrointestinal hormone receptors, hypothalamic neuropeptides, and genes related to lipid metabolism were exclusively determined at the gene expression level. However, the genes we measured herein have also been assessed at the protein [2930] and phenotype [3132] levels in other studies.
In conclusion, the gene expression results suggest that dietary Korean PNO consumption may decrease body fat accumulation and could prevent the dysregulation of the appetite-suppressing pathway induced by HFD consumption. Nevertheless, additional phenotypic analyses are needed to confirm the overall effects of Korean PNO on whole-body lipid and energy metabolism.
Go to :
 XML Download
XML Download