

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The adaptive immune response is mediated by diverse receptors on the surface of T cells that allow for recognition of pathogens [8]. Accordingly, identification of the T cell receptor (TCR) repertoire can be used to monitor the host immune response. Characterization of TCR repertoires have proven useful in cancer and autoimmune disease patients who received immunotherapy [34]. Sims et al. [30] sequenced TCRs of glioma αand β-chains. They showed that more divergent VJ-independent repertoires are associated with tumors than non-neoplastic tissues. Yu et al. [34] evaluated the associations between TCR repertoire and symptom remission in systemic lupus erythematosus (SLE) patients treated with glucocorticoids. They suggested that monitoring of the TCR complimentary determining region (CDR) 3 is related to prognosis in SLE, suggesting that it mirrors therapeutic efficacy. T cells are also actively involved in atherosclerosis [14,22]. Jia et al. [14] reported epigenetic suppression of forkhead box that was related with a decrease in regulatory T cells, resulting in an increased risk of atherosclerosis. In addition, the T cell response is involved in the initiation of plaque disruption [10].
SAH occurs spontaneously via ruptured aneurysm, arteriovenous malformation (AVM), or traumatic brain injury. The mortality rate of SAH due to ruptured cerebral aneurysm exceeded 20% [21,33]. Although an aneurysm itself may be successfully occluded, neurologic complications can occur due to direct brain injury and vascular dysregulation [26]; such complications are associated with poor neurologic outcomes if not promptly treated. Consequently, SAH patients require multimodal monitoring in the neurointensive care unit for early detection of neurologic complications and subsequent optimal intervention [26]. In attempts to understand the pathophysiology of SAH, many researchers have focused on genomic and proteomic analyses; by comparison, the adaptive immune response has not been well studied [1]. Recently, we reported the TCR repertoire for the first time in SAH patients with and without delayed cerebral ischemia (DCI) [18]. In addition, we further evaluated the T cell immunology based on SAH severity to delineate SAH pathogenesis.
MATERIALS AND METHODS
Study population
This investigation was approved by the Institutional Review Board of the participating hospital (No. 2016-3, 2017-9, and 2018-6). Informed consent was received from all patients or their relatives. The study cohort was derived from the participating hospital’s stroke database, which is part of an ongoing prospective, observational project in the regional medical center of the district; all patients presented to the hospital between March 2016 and December 2019 [7,11,16,19,20,27,33]. We selected SAH patients based on the following conditions : 1) age over 18 years old; 2) spontaneous SAH due to ruptured aneurysms with saccular appearance; and 3) treatment with coil embolization. The exclusion criteria were : 1) fusiform, dissection, and infectious aneurysms; 2) angiogram-negative SAH; and 3) SAH concomitant with other cerebrovascular diseases including AVM and dural arteriovenous fistula [17].
We aimed to compare the composition and variation of the TCR β-chain (TCRB) CDR3 repertoire between the day of admission and at the time of follow-up, 7 days afterwards. Additionally, we divided patients into two groups based on SAH severity, good- and poor-grade, and further compared TCRB repertoire between these groups. Good-grade SAH was defined as initial Hunt and Hess (H-H) grade less than III. Clinical and radiologic medical information were reviewed. Clinical outcome was assessed using modified Rankin Scale (mRS) score 6 months after SAH.
DNA extraction and high-throughput sequencing
Peripheral blood samples were prepared on the day of admission and follow-up. Genomic DNA was extracted from whole blood using the QIAamp DNA Blood Midi Kit (Qiagen, Hilden, Germany), quantified with a DropSense96, and diluted for library preparation in buffer according to a standard. Then, sample data was generated using the immunoSEQ assay (Adaptive Biotechnologies, Seattle, WA, USA). The somatically rearranged locus CDR3 region was amplified using a biascontrolled multiplex polymerase chain reaction (PCR) [4,18,29]. Specifically, the first PCR was performed for every V and J gene segments using forward and reverse amplification primers, and the hypervariable CDR3 was amplified. Then, proprietary barcode sequence and Illumina adapter sequences (https://support.illumina.com) were performed in the second PCR [28]. CDR3 libraries were sequenced on an Illumina instrument according to the manufacturer’s instructions.
Analysis of TCRB repertoire
The raw Illumina sequence reads were demultiplexed and then further processed in the following steps : removal of the adapter and primer sequences, identification and correction of technical errors, and removal of primer dimer, germline, and other contaminant sequences. Then, the data were filtered and clustered to merge closely-related sequences using relative frequency ratio and modified nearest-neighbor algorithm. In the variable region of the chains, antibody is generated by the combination of the gene fragments, V, D, and J genes. The V, D, and J regions are separated and encoded by single or multiple axons [3]. Accordingly, the V, D, and J genes were defined based on annotation in compliance with the IMGT database (www.imgt.org). The biological CDR3 locus sequences were normalized and quantified [4,18]. The immunoSEQ Analyzer toolset was used for data analysis. The results of comparisons are described as median (25th–75th percentile) using the Mann-Whitney U test. p values less than 0.05 are considered statistically significant. Analyses were conducted with SPSS ver. 19 (SPSS, Chicago, IL, USA) [13].
RESULTS
Baseline characteristics of the study
TCRB CDR3 repertoires from six SAH patients were analyzed and compared at two different time points, admission and 7-day follow-up. Three patients presented with goodgrade SAH, while the three others had poor-grade SAH. The median age was 53.5 years (range, 41–77). Four of the patients were female (66.7%). Most aneurysms (n=5, 83.3%) were located in the anterior circulation. Two patients experienced severe DCI and recovered well after chemical angioplasty. Three patients with good-grade SAH recovered well without significant disability. However, one out of three patients (33.3%) exhibited poor neurologic outcome defined by mRS score of 3 (Table 1).
Table 1.
Clinical characteristics of the enrolled patients
| Pt No. | Age (years) | Sex | Hunt and Hess grade* | Fisher grade | Aneurysm site | DCI | 6-month mRS |
|---|---|---|---|---|---|---|---|
| 1 | 61 | F | II | 2 | MCA | No | 0 |
| 2 | 50 | F | II | 3 | MCA | Yes | 0 |
| 3 | 77 | F | III | 3 | A-com | No | 0 |
| 4 | 47 | M | IV | 4 | PICA | No | 3 |
| 5 | 57 | M | IV | 4 | A-com | No | 0 |
| 6 | 41 | F | IV | 3 | MCA | Yes | 1 |
![]()
Sequence, clonality, and CDR3 lengths of TCRB
Variables such as total and unique sequences, clonality, and frequency of the TCRB CDR3 repertoire were measured (Table 2). Clonality using the Shannon index at admission and follow-up were 0.059 (0.037–0.038) and 0.027 (0.014–0.082) (p=0.699), respectively. Clonality of poor-grade (0.025 [0.011–0.038]) SAH was significantly lower than that of good-grade SAH (0.095 [0.079–0.101]; p=0.026; Fig. 1A).
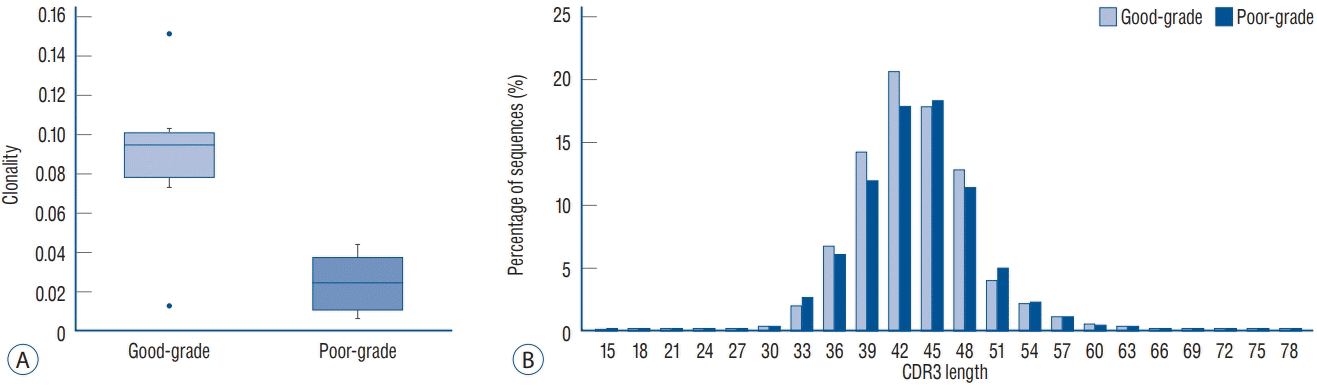
Fig. 1.
Comparison of TCRB clonality (A) and CDR3 length repertoire (B) according to SAH severity, good- vs. poor-grade. Good-grade SAH patients had higher clonality and shorter CDR3 length than did poor-grade SAH patients. CDR : complimentary determining regions, TCRB : T cell receptor β-chain, SAH : subarachnoid hemorrhage.
![]()
Table 2.
TCRB CDR3 repertoire – total sequences, unique sequences, and clonality – in patients with subarachnoid hemorrhage at two times, admission and follow-up
| Pt No. |
Nucleic acid* |
Total |
Frame |
Sequences (amino acid)† |
|||
|---|---|---|---|---|---|---|---|
| Group | Total | Unique | Unique | Clonality | Max frequency (%) | ||
| 1 | Adm | 6762 | 5721 | 4961 | 4069 | 0.094 | 5.74 |
| F/U | 8187 | 6858 | 5919 | 4803 | 0.096 | 6.44 | |
| 2 | Adm | 23275 | 19296 | 13403 | 10858 | 0.150 | 4.74 |
| F/U | 3431 | 2826 | 3023 | 2476 | 0.013 | 0.61 | |
| 3 | Adm | 5213 | 4222 | 4698 | 3788 | 0.007 | 0.35 |
| F/U | 4171 | 3334 | 3565 | 2837 | 0.015 | 0.48 | |
| 4 | Adm | 1797 | 1509 | 1318 | 1082 | 0.073 | 5.56 |
| F/U | 4019 | 3360 | 2816 | 2291 | 0.102 | 9.16 | |
| 5 | Adm | 3743 | 2901 | 2808 | 2229 | 0.044 | 2.99 |
| F/U | 8032 | 6244 | 5981 | 4743 | 0.039 | 1.97 | |
| 6 | Adm | 2261 | 1721 | 1785 | 1380 | 0.035 | 3.05 |
| F/U | 4648 | 3657 | 4174 | 3271 | 0.009 | 0.47 | |
![]()
Read frequency of ≤0.005% to <0.05% dominated the clonal expansion, but the changes between admission and follow-up did not show a definite association (Supplementary Table 1). We ranked TCRB clonotype frequencies and compared repertoire diversity using the Simpson diversity index (D) [5], where a score close to one indicates high diversity (Supplementary Fig. 1). Overall, the diversity indices ranged from 0.993 to 0.999 at admission and from 0.987 to 0.999 at follow-up. Poor-grade SAH patients had diversity scores of 0.998 and 0.999, which were higher than the scores of good-grade SAH patients.
CDR3 lengths ranged from 15 to 78 nucleotides, with a peak at 42 in good-grade and at 45 in poor-grade SAH patients. The five most common CDR3 lengths were 36, 39, 42, 45, and 48 nucleotides. The results are calculated as the average of five individuals. Good-grade SAH cases tended to have shorter CDR lengths compared to poor-grade SAH (Fig. 1B). The ten most expanded sequences were presented and compared in terms of amino acid levels (Fig. 2) [32]. The distribution of the top ten most common sequences was different in each patient. Overall, poor-grade SAH patients had a lower proportion of the top ten most common sequences than did good-grade SAH patients.
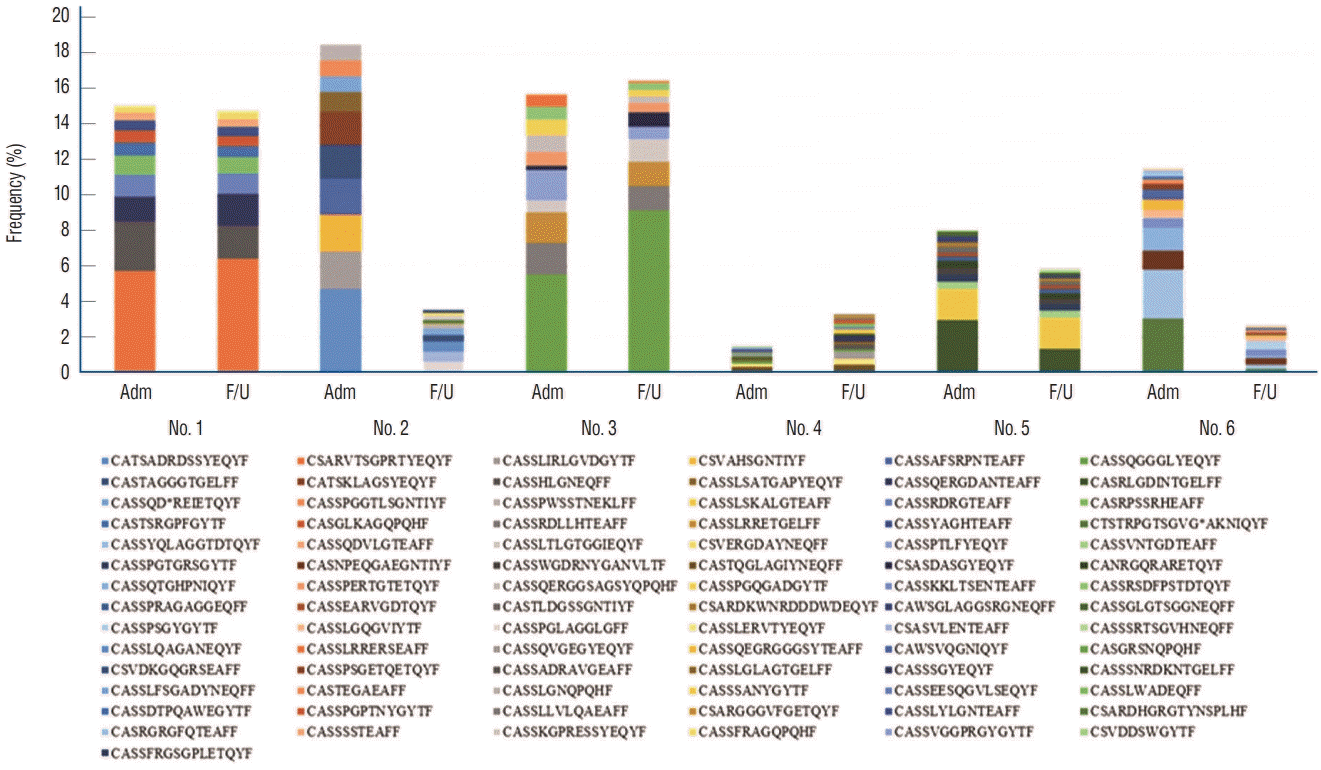
Fig. 2.
Quantification of TCRB CDR3 sequences in each patient between the day of admission and the 7-day follow-up. The 10 most common sequences are presented in different colors. Patients who presented with poor-grade SAH tended to show a low proportion of the top 10 most common sequences compared to those with good-grade SAH at admission. Adm : admission, F/U : follow-up, TCRB : T cell receptor β-chain, CDR : complimentary determining regions, SAH : subarachnoid hemorrhage.
![]()
Distribution of TCRBV and TCRBJ
We analyzed the distribution of the unique clonotypes in both the TCRBV and TCRBJ gene segments. Overall, TCRBV gene segments from the follow-up day had different clonotypes than did those from the admission day, but the distribution of TCRBJ gene segments was similar between days. Compared to good-grade SAH, poor-grade SAH tended to exhibit a great number of different clonotypes of the TCRBV gene segments (Fig. 3). We analyzed the relative frequencies of V-J combinations using different colors representing disease severity. We obtained 754 annotated V-J pairs. TCRBV21-01/TCRBJ01-05, TCRBV21-01/TCRBJ01-04, and TCRBV21-01/TCRBJ02-04 were the most commonly increased V-J pairs in poor-grade SAH compared to good-grade SAH patients (Supplementary Fig. 2). We further compared the V-J combinations at two different time points (admission and 7-day follow-up) in good-grade and poor-grade SAH patients, respectively. The results showed that TCRBV19-01/TCRBJ02-04 was the most increased V-J pair and TCRBV28-01/TCRBJ02-04 was the most decreased V-J pair in good-grade SAH. Regarding poorgrade SAH patients, the most increased and decreased V-J pairs were TCRBV28-01/TCRBJ02-06 and TCRBV30-01/TCRBJ02-04, respectively (Fig. 4).
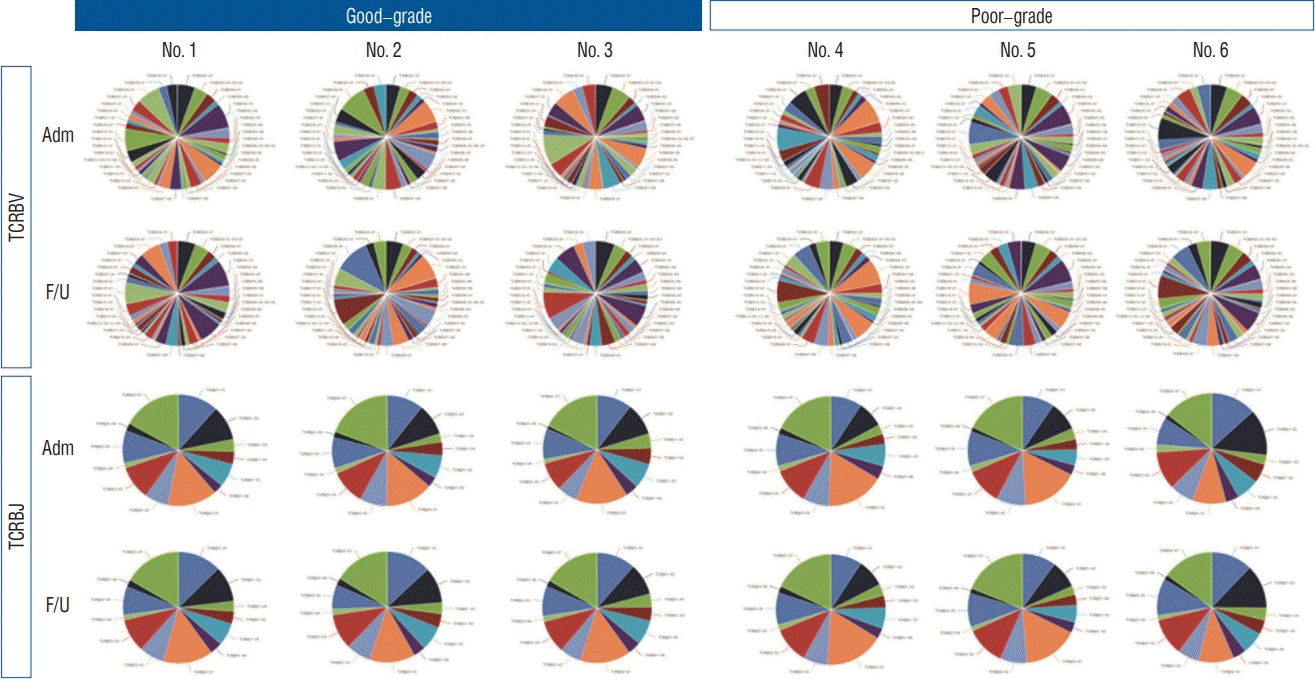
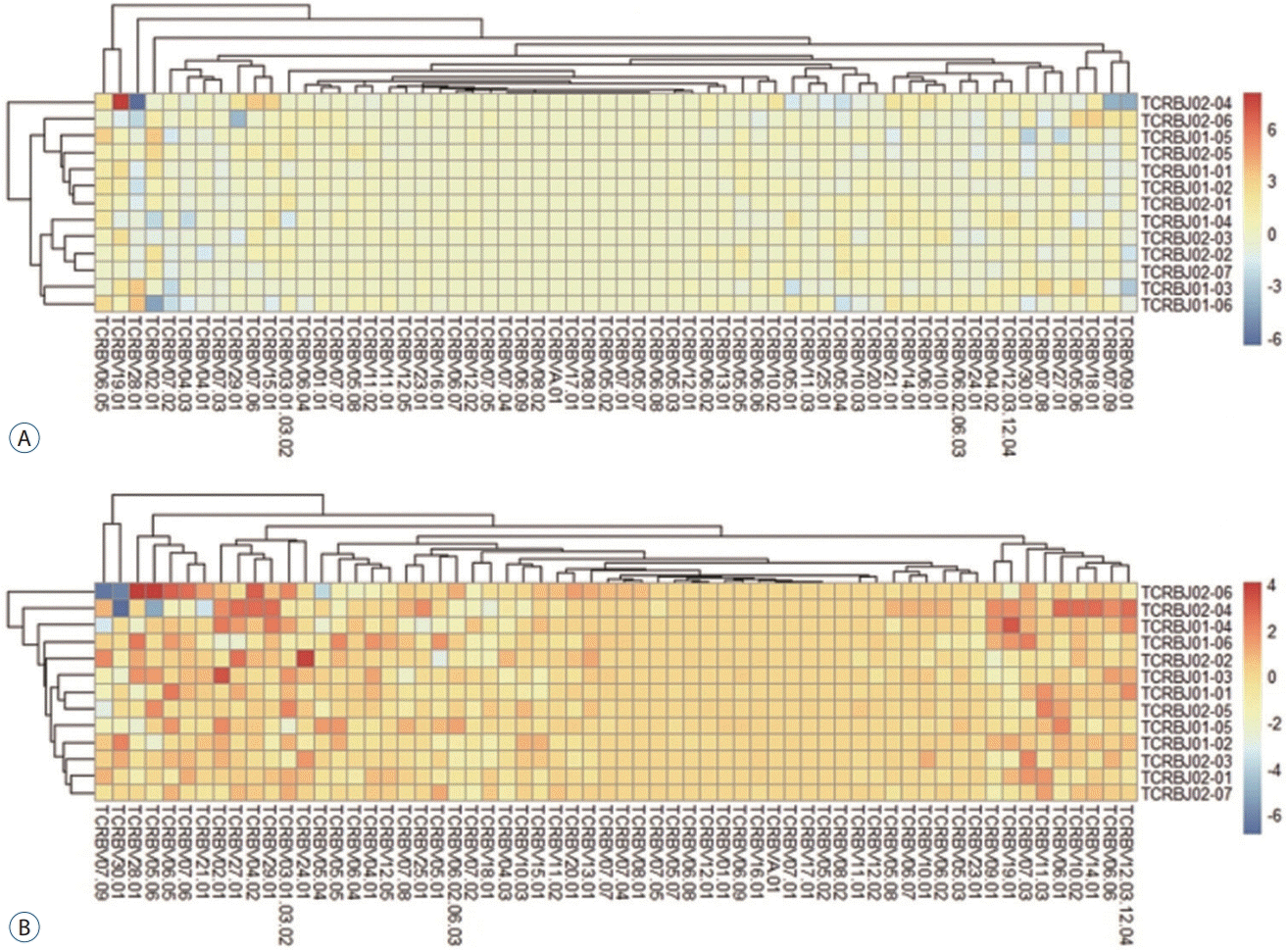
Fig. 3.
Distribution and change in the unique clonotypes of the TCRBV and TCRBJ genes in patients with SAH. Compared to TCRBJ, TCRBV showed different distributions between the day of admission and the 7-day follow-up. Different colors are used to represent gene frequencies in each patient. TCRBJ : T cell receptor β chain J segment, TCRBV : T cell receptor β chain V segment, Adm : admission, F/U : follow-up, SAH : subarachnoid hemorrhage.
![]()
Fig. 4.
Comparison of average TCRB V-J gene utilization at admission and during the 7-day follow-up according to SAH severity of good (A) and poor (B)-grade. V gene and J gene segments are arranged on the x- and y-axis. Different colors indicate the differences in gene frequencies of poor-grade SAH patients compared with good-grade SAH patients. TCRB : T cell receptor β-chain, SAH : subarachnoid hemorrhage.
![]()
DISCUSSION
T cells recognize antigens with the major histocompatibility complex-TCR complex. The TCR repertoire mainly relies on diversity in the α- and β-chains [8]. Among the hypervariable regions of CDRs chains, CDR3 has been targeted for high-throughput immune sequencing due to its diversity [8,32]. Chang et al. [5] compared TCRB repertoires in rheumatoid arthritis patients who received different medications. In their study, the diversity of the TCRB repertoire varied according to different biological medications and was inversely correlated with disease activity [5]. Sun et al. [31] reported that TCRB diversity was lower overall in neonatal sepsis than in controls, with different clonotypes and V-J gene combinations in the two. In addition, shortening of CDR3 was suggested to increase autoimmune disease risk by increased self-recognition during thymic T cell development in type 1 diabetus mellitus [9]. Therefore, the TCR repertoire can function as a mirror of the health of the host, a marker of disease severity, or a marker of biological response for immunotherapy [2].
It has been postulated that SAH can spark innate and adaptive immune responses [25]. The study of immune response in SAH has generally focused on pro-inflammatory cytokines, which are responsible for brain edema after hemorrhagic insult, and treatments to alleviate the condition [6]. Hemorrhage leak into the subarachnoid space can trigger recruitment of leukocytes and inflammatory responses [15]. SAH patients exhibited increased levels of interleukin (IL)-6 and soluble IL-2 receptor in the cerebrospinal fluid [24]. Chen et al. [6] reported a beneficial effect of melatonin on preservation of tight junction proteins in the blood-brain barrier, resulting in decreased brain edema in an experimental SAH model in rats. In their study, the levels of pro-inflammatory cytokines such as IL-6, IL-1β, and tumor necrosis factor-α were suppressed after melatonin injection. Moraes et al. [25] also reported higher levels of innate immune cells such as CD14++, CD16+, and CD69+ following SAH. However, the adaptive immune response, and TCRB repertoire in particular, has not been well studied in SAH patients. In this study, we explored the composition and variation of the TCRB CDR3 repertoire to understand T cell-mediated adaptive immunity after SAH. TCRB CDR3 repertoires are dynamic in nature according to disease severity. Poor-grade SAH patients showed significantly decreased clonality and increased diversity as compared with good-grade SAH patients, as well as longer CDR3 amino acid lengths. Interestingly, disease severity was associated with differences in VJ gene usage of TCRB and CDR3 lengths, which can affect TCR rearrangement. Therefore, further studies on the role of individual clonotypes in the development of clinical complications such as DCI or SAH-induced immunocompromised states are necessary.
In this study, we only included SAH patients who received endovascular coil embolization though the transfemoral approach, not those with craniotomy and clipping. The host immune response to tissue injury can affect systemic inflammatory and anti-inflammatory responses [23]. Marik and Flemmer [23] reported increased T-helper 2 cells which can result in impaired cell-mediated immunity after physical injury. Compared to coil embolization, clipping is associated with concerns about infection of the surgical site and tissue injury. Accordingly, we only sequenced and analyzed the TCRB repertoire in SAH patients with coiling to reduce inherent bias from different treatment methods. Therefore, patients who underwent craniotomy and clipping may have different TCRB CDR3 repertoires following SAH, which requires additional investigation.
The study limitations are as follows. First, the small sample size may be a concern, although this is the largest series to date involving SAH patients. Most previous studies enrolled less than 10 patients due to economic concerns [5,12]. Our previous study analyzing the TCRB repertoire in DCI also included five SAH patients [18]. Nevertheless, economic concerns cannot justify the results of the study. Accordingly, further studies are needed to investigate the adaptive immune response in a large number of the SAH patients to validate our findings. Second, we did not evaluate the TCR α-chain. Although most research has been done on either the α- or β-chain, one at a time [8], quantification of both the α- and β-chains can more accurately determine the TCR repertoire. Third, we have focused on the clinical grading system of H-H grade, but not Fisher grade. T cell adaptive immune response via extravasation of blood can trigger additional inflammatory response. We have analyzed and compared the average TCRB V-J gene utilization according to Fisher grades, II and III (A) and IV (B) (Supplementary Fig. 3). TCRBV19-01/TCRBJ02-04 was the most increased V-J pair in Fisher grades II and III in the 7-day follow-up and TCRBV28-01/TCRBJ02-04 was the most decreased V-J pair. In case of Fisher grade IV, TCRBV28-01/TCRBJ02-06 and TCRBV30-01/TCRBJ02-04 were the most increased and decreased V-J pairs, respectively. Nevertheless, the small number of enrolled patients may be a concern for the interpretation, suggesting the need for further TCRB studies focusing on Fisher grade. Fourth, different drug regimens, which possibly affect immunity, cannot accurately reflect the adaptive immune response. Accordingly, additional multicenter studies based on drug regimen are needed to evaluate TCRB repertoire.
CONCLUSION
The TCRB repertoire has a dynamic nature following SAH. Poor-grade SAH patients displayed a reduction in clonality and an increase in diversity and CDR3 length. Monitoring TCRB CDR3 repertoires can provide a better understanding of the adaptive immune response and could be useful for identification of therapeutic targets in SAH patients.
 XML Download
XML Download