

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Colorectal cancer (CRC) was ranked the third for incidence rate and the second for mortality rate among all cancers worldwide in 2018 [1]. The cancer stem cells (CSCs) model accounts for the high recurrence, metastasis, and drug-resistance characteristics of CRC. CSCs are a small portion of cancer cells with stem cell characteristics, including self-renewal capacity, differentiation, and tumorigenicity [2]. Self-renewal allows stem cells to produce other stem cells that possess the same developmental and replication potential. Differentiation is the development of tissue-specific specialized cells, and tumorigenicity is the ability to form tumors. CSCs exhibit up-regulated self-renewal capacity and invasion properties and are resistant to existing therapeutic approaches [23]. Therefore, targeting CSC may be an effective strategy to develop efficient anticancer therapy.
CSCs can be discriminated from other cancer cells based on the expression of specific CSC markers. CSC markers, such as CD44, CD133, and aldehyde dehydrogenase 1 (ALDH1), are most commonly used for CSC isolation in CRC [4]. Notch homolog 1 (Notch1), a single transmembrane-receptor, plays a critical role in maintaining the self-renewal capacity and suppressing differentiation-related gene expression in colon CSCs [5]. CRC cells with a high level of Sox2 expression induced sphere formation and increased stem cell markers, such as CD24 and CD44 [6]. The Wnt/β-catenin pathway is fundamental in embryonic development and in maintaining the functional integrity and growth of CSCs [7]. The translocation of β-catenin to the nucleus leads to the transcriptional activation of Wnt target genes. Furthermore, the Wnt/β-catenin signaling pathway plays a critical role by regulating the self-renewal of epithelial stem cells, but aberrant Wnt/β-catenin signaling contributes to CRC development [8].
β-carotene (BC) is an active vitamin A precursor and well known-antioxidant found in abundance in many fruits and vegetables with deep yellow, orange, or green colors. Increased plasma concentrations of BC and due to high dietary intakes of BC have been shown to decrease the incidence of several cancers, such as lung cancer, non-Hodgkin lymphoma, and epithelial cell cancer [910]. Previously, the protective effect of BC against colon cancer has been reported in the 1,2-dimethylhydrazine (DMH)-induced colon cancer model in vivo [11]. BC also increases the expression of β,β-carotene 15,15′-monooxygenase (BCMO1). When BCMO1 expression was inhibited, cell invasion was increased in colon cancer cells [12]. In addition, BC inhibited cancer stemness and induced neuronal differentiation of neuroblastoma in vitro and in vivo, demonstrating an anti-CSC effect [1314]. However, there is a lack of evidence about the inhibitory effect of BC on colon cancer stemness.
Go to : 
MATERIALS AND METHODS
Cell culture
The human colon cancer cell lines, HCT116 and HT-29, were purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA) and were maintained in McCoy's 5A medium (Welgene, Daegu, Korea) supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA) and 1% penicillin/streptomycin (100 U/mL and 100 μg/mL; Invitrogen, Carlsbad, CA, USA). The cells were cultured in humidified air at 37°C with 5% CO2. BC was purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA).
Isolation of CSCs
Colon CSCs were isolated from HCT116 and HT-29 cells after double-staining for colon CSC markers, CD44 and CD133, as described previously [15]. Briefly, colon cancer cells were harvested and washed with FACS buffer (10% FBS and 0.5 mM ethylenediaminetetraacetic acid in phosphate-buffered saline [PBS]) and stained with CD44 and CD133 monoclonal antibodies (Miltenyi Biotec, Bergisch Gladbach, Germany) at 4°C for 30 min in the dark. Then, stained cells were sorted as colon CSCs by FACSAria (BD, Franklin Lakes, NJ, USA).
Clonogenic assay
To determine the self-renewal capacity of colon CSCs, the clonogenic assay was undertaken as described previously [16]. Briefly, both CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cells were seeded (250 or 400 cells/well, respectively) in 6-well plates and were incubated with BC (0, 20, and 40 μM) for 10–12 days. The colonies were then fixed with 0.9% sodium chloride (NaCl) solution and stained with crystal violet (Sigma-Aldrich). The stained colonies were counted, and the plating efficiency was calculated: (number of colonies/number of seeded cells) × 100%.
Sphere formation assay
Another analysis to assess self-renewal capacity for CSCs is the sphere formation assay, which was performed as described previously [14]. Briefly, CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cells (1 × 104 or 2 × 104 cells/well, respectively) were cultured in 6-well plates coated with 10% poly-(2-hydroxyethyl methacrylate) (polyHEMA; Sigma-Aldrich). DMEM-F12 (1:1; Welgene) containing 20 ng/mL epidermal growth factor (Pepro Tech, London, UK), 40 ng/mL basic fibroblast growth factor 2 (Pepro Tech), and 2% B27 (Invitrogen) were used for the sphere medium. After 24 h, cells were treated with BC (0, 20, and 40 μM) for 8–10 days, and the number of spheres was counted and photographed using a phase contrast microscope (Olympus, Tokyo, Japan).
Human primary cell isolation and culture
Primary CRC tissues were obtained from patients at the Seoul National University-Seoul Metropolitan Government Boramae Medical Center (Seoul, Korea). The tumor specimens were placed in DMEM/F12 medium with 6% penicillin/streptomycin and 3 μg/mL amphotericin B (Sigma-Aldrich) for transport to the laboratory for process and analysis. Tumors were washed several times with cold PBS containing 1% penicillin/streptomycin, then finely minced, followed by enzymatic digestion using 1.5 mg/mL collagenase I (Thermo Fisher Scientific, Waltham, MA, USA) and 20 μg/mL hyaluronidase (Sigma-Aldrich) in McCoy's 5A medium at 37°C for 1 h in a shaking incubator. The digested cancer tissues were filtered through a 40-μm-pore size nylon cell strainer and centrifuged at 1,250 × g for 10 min. The supernatant was removed, and red blood cell lysis buffer (Sigma-Aldrich) was added. The cells were subsequently cultured in McCoy's 5A medium with 10% FBS, 6% penicillin /streptomycin, and 3 μg/mL amphotericin. This study was approved by the ethical review board of the Boramae Medical Center (IRB No. 10-2017-5).
In vivo xenograft model
Male 5-week-old Balb/c nu/nu mice were purchased from Central Lab Animal Inc. (Seoul, Korea). After 1-week of adaptation, the mice were randomly assigned to 3 groups (n = 6/group) as follows: (a) mice were fed control diet (TC); (b) mice were orally supplemented with 6 mg/kg body weight (BW) of BC (BC 6); (c) mice were orally supplemented with 12 mg/kg BW of BC (BC 12). Mice were fed vehicle (corn oil) for the TC group or BC dissolved in corn oil for BC groups twice a week for 3 weeks. After 3 weeks, mice were subcutaneously injected with 1 × 105 CD133+CD44+ HCT116 cells mixed with growth factor-reduced matrigel (1:1) (BD Bioscience Laboratory, Bedford, MA, USA) into both sides of the back of the animal. Tumor volume was measured twice a week using digital calipers: volume = length (mm) × width2 (mm2) × 0.5. Mice were fed BC for 7 weeks more and then sacrificed. Animal care and the experimental procedure for this study were approved by the Animal Care and Use Committee of Ewha Womans University (IACUC No. 17-025).
Real-time polymerase chain reaction (RT-PCR) analysis
RT-PCR analysis was performed as previously described [17]. RNA was extracted by TRIzol Reagent (Invitrogen) and a RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific) was used. Samples were mixed with 2X SYBR Green PCR Master Mix (Qiagen, Hilden, Germany), followed by RT-PCR. The reaction was carried out using a Rotor-Gene Q real-time cycler (Qiagen) under the following conditions: initiation at 95°C for 5 min, denaturation at 95°C for 15 s, and annealing and extension at 60°C for 10 s. Glyceraldehyde 3 phosphate dehydrogenase (GAPDH) was used as an internal control. The PCR primers used were specific for human CD44: 5′- CCA ATG CCT TTG ATG GAC C -3′ (forward) and 5′- TCT GTC TGT GCT GTC GGT GAT -3′ (reverse); CD133: 5′- ATA CCT GCT ACG ACA GTC GT -3′ (forward) and 5′- TGG ATG CAG AAC TTG ACA AC -3′ (reverse); ALDH1A1: 5′- TGT TAG CTG ATG CCG ACT TG -3′ (forward) and 5′- TTC TTA GCC CGC TCA ACA CT -3′ (reverse); NOTCH1: 5′- AGAG GCG TGG CAG ACT ATG C -3′ (forward) and 5′- CTT GTA CTC CGT CAG CGT GA -3′ (reverse); GAPDH: 5′- AGA AGG CTG GGG CTC ATT TG -3′ (forward) and 5′- AGG GGC CAT CCA CAG TCT TC -3′ (reverse).
Western blot analysis
Western blot was performed as previously described [18]. To extract whole proteins, RIPA lysis buffer (150 mM NaCl, 50 mM Tris-hydrochloride [pH 7.5], 1% Nonidet, P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS], 1 mM phenylmethylsulfonyl fluoride, 1 mM Na3VO4, and 1 mM sodium fluoride) were used. Protein concentrations of the samples were determined by Bradford protein assay (Bio-Rad, Hercules, CA, USA). An equal concentration of proteins was loaded and separated on SDS polyacrylamide gels by electrophoresis, then transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA) and blocked with 5% bovine serum albumin or skim milk in TBS-T. The blocked membranes were incubated with primary antibodies against Notch1 (Novus Biologicals, Littleton, CO, USA), Sox 2 (Abcam, Cambridge, UK), β-catenin (Santa Cruz Biotechnology, Santa Cruz, CA, USA), CD44 (Cell Signaling Technology, Danvers, MA, USA), CD 133 (MACS, Bergisch Gladbach, Germany), and β-actin (Abcam) in a cold room overnight. After washing, membranes were incubated with secondary antibodies (Santa Cruz Biotechnology) for 1 h and visualized using an enhanced chemiluminescence reagent (Animal Genetics, Inc., Suwon, Korea).
Statistical analyses
All data are expressed as the mean ± standard error of the mean of at least 3 independent experiments. Analysis of variance and the Newman-Keuls post hoc test were performed using GraphPad PRISM software (GraphPad Software, Inc., San Diego, CA, USA). Statistical significance was set at P-value < 0.05.
Go to :
RESULTS
BC suppressed the self-renewal capacity of colon CSCs
The clonogenic assays is designed to evaluate the differences in the reproductive ability of cells by measuring colonies formed from single cell cultures and assess the self-renewal capacity of CSCs [19]. CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cells were treated with BC (20 or 40 μM), and the number of colonies were compared to the untreated control group (Ctrl) (Fig. 1A). The ability to generate colonies from CD133+CD44+ HCT116 cells was decreased by 39.1% and 86.3% following treatment with 20 and 40 μM BC, respectively, compared to the Ctrl (both P < 0.001; Fig. 1B). Similarly, the proliferative ability was decreased by 24.4% and 54.4% for CD133+CD44+ HT-29 cells treated with 20 and 40 μM BC, respectively compared to the Ctrl (both P < 0.001; Fig. 1C).
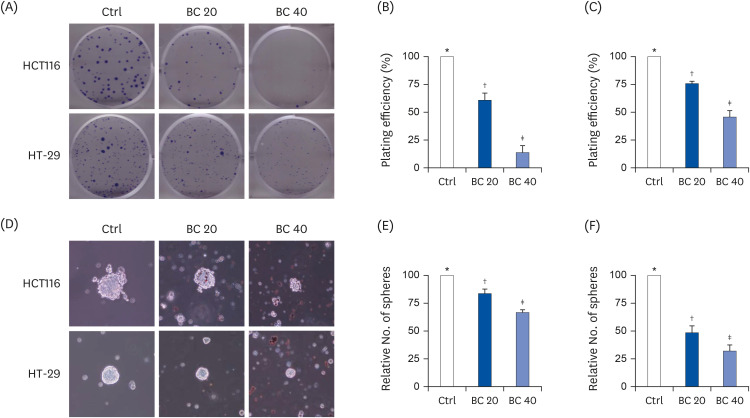 | Fig. 1BC suppressed self-renewal capacity of colon cancer stem cells by inhibiting clonogenicity and sphere formation. CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cells were treated with BC (0, 20, and 40 μM) for 8–14 days. Clonogenic assay (A-C) and sphere formation assay (D-F) were performed. (A, D) Representative images (magnification, 100 ×) and number of colonies or spheres are shown in (B, E) CD133+CD44+ HCT116 and (C, F) CD133+CD44+ HT-29 cells.Ctrl, control group; BC, β-carotene.
*,†,‡The labels on the bar graph indicate the values that significantly differ from each other (P < 0.05) by one-way analysis of variance for multiple comparisons.
|
A previous study demonstrated that treatment with 20 and 40 μM BC suppressed sphere formation in CD133+CD44+ HCT116 [20]. Therefore, the effect of BC on sphere formation ability of CD133+CD44+ HT-29 and CD133+CD44+ HCT116 cells was confirmed in the present study (Fig. 1D). Treatment of 20 and 40 μM BC to CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cells decreased the number and size of spheres. Sphere formation was suppressed by up to 16.2% (P < 0.01) and 33.2% (P < 0.001) with 20 and 40 μM BC treatment, respectively, compared to the Ctrl for CD133+CD44+ HCT116 cells (Fig. 1E). Likewise, sphere formation was suppressed by 51.7% and 68.3% for CD133+CD44+ HT-29 cells treated with 20 and 40 μM BC, respectively, compared to the Ctrl (both P < 0.001) (Fig. 1F). These results suggested that BC suppressed the self-renewal capacity of colon CSCs.
BC suppressed mRNA expressions of CSC markers and Wnt/β-catenin signaling in human colon CSCs and primary cells from CRC patients' tumor tissues
To determine whether BC regulates the expression of colon CSC markers, mRNA levels of major CSC markers, including CD44, CD133, ALDH1A1, and NOTCH1, were analyzed using RT-PCR. BC treatment down-regulated all CSC markers in CD133+CD44+ HCT116 (Fig. 2). In particular, 40 μM BC treatment down-regulated CD44, CD133, ALDH1A1, and NOTCH1 by 53.0% (P < 0.001), 51.1% (P < 0.001), 65.7% (P < 0.001), and 57.6% (P < 0.001) in CD133+CD44+ HCT116 cells (Fig. 2A) and by 85.8% (P < 0.001), 85.2% (P < 0.001), 86.9% (P < 0.001), and 80.7% (P < 0.001) in CD133+CD44+ HT-29 cells (Fig. 2B), respectively, compared to the Ctrl.
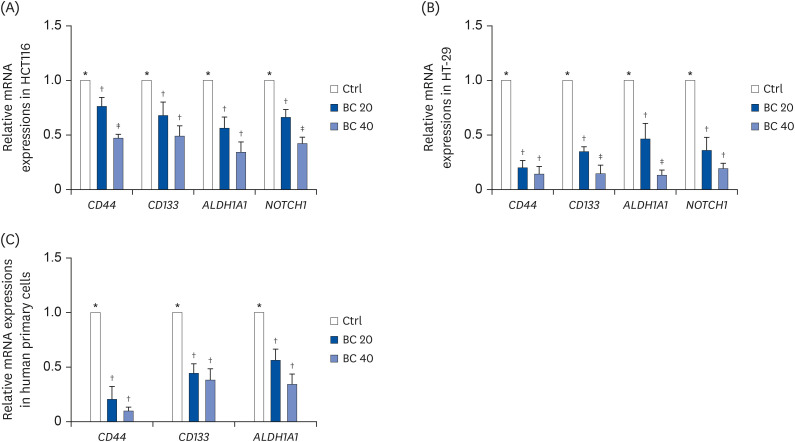 | Fig. 2BC suppressed mRNA expression of colon CSC markers in human colon CSCs and primary cells from CRC patients' tissues. After BC treatment (0, 20, and 40 μM) for 6 or 8 days, mRNA levels of colon CSC markers, CD44, CD133, ALDH1A1, and NOTCH1, were evaluated in (A) CD133+CD44+ HCT116 and (B) CD133+CD44+ HT-29 cells. (C) mRNA levels of colon CSC markers, CD44, CD133, and ALDH1A1, were analyzed in human primary cells from CRC patients by real-time polymerase chain reaction. GAPDH was used as a loading control.CSC, cancer stem cell; Ctrl, control group; BC, β-carotene; CRC, colorectal cancer.
*,†,‡The labels on the bar graph indicate the values that significantly differed from each other (P < 0.05) according to one-way analysis of variance for multiple comparisons.
|
In human primary cells from CRC tumor tissue, CSC markers, including CD44, CD133, and ALDH1A1 were down-regulated by BC treatment (Fig. 2C). In particular, 40 μM BC treatment suppressed mRNA expression of CD44, CD133, and ALDH1A1 by 90.3% (P < 0.001), 62.0% (P < 0.01), and 53.4% (P < 0.01), respectively compared to the Ctrl. These data confirmed the in vitro anti-CSC effect of BC.
BC suppressed protein expression of CSC markers and Wnt/β-catenin signaling in human colon CSCs
To confirm these down-regulated protein levels and investigate the effect of BC on Wnt/β-catenin signaling, the expressions of major CSC markers, including CD44, CD133, Notch1, Sox2, and β-catenin, were investigated using western blot analysis. BC treatment significantly suppressed the expressions of CSC markers and β-catenin in CD133+CD44+ HCT116 (Fig. 3A and B) and HT-29 cells (Fig. 3C and D). In particular, 40 μM BC treatment down-regulated the expressions of CD133, Notch1, and Sox2 by 72.0% (P < 0.001), 62.7% (P < 0.05), and 47.0% (P < 0.05) in CD133+CD44+ HCT116 cells and by 53.1% (P < 0.05), 50.7% (P < 0.001), and 59.0% (P < 0.01), in CD133+CD44+ HT-29, respectively, compared to the Ctrl. Although BC treatment tended to down-regulate CD44 expression in CD133+CD44+ HCT116 cells, the expression was significantly down-regulated in CD133+CD44+ HT-29 cells treated with 40 μM BC compared to the Ctrl (P < 0.01). Furthermore, we analyzed the protein level of the Wnt/β-catenin pathway, to identify if the anti-CSCs effect of BC was related to this CSC signaling pathway. β-catenin expression was down-regulated in the BC 40 group by 54.4% (P < 0.05) and 66.1% (P < 0.01) in CD133+CD44+ HCT116 cells and HT-29 cells, respectively.
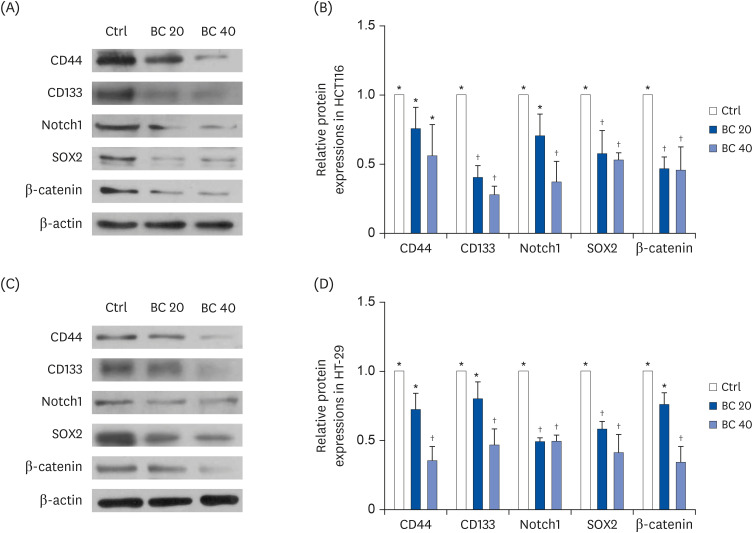 | Fig. 3BC suppressed protein expression of CSC markers and the Wnt/β-catenin signaling pathway in human colon CSCs. After BC treatment (0, 20, and 40 μM) for 6 or 8 days, protein levels of CD44, CD133, Notch1, SOX2, and β-catenin in (A, B) CD133+CD44+ HCT116 and (C, D) CD133+CD44+ HT-29 cells were detected by western blot analysis. β-actin was used as a loading control.Ctrl, control group; BC, β-carotene; CSC, cancer stem cell.
*,†The labels on the bar graph indicate the values that significantly differ from each other (P < 0.05) by one-way analysis of variance for multiple comparisons.
|
BC supplementation suppressed tumor volume and delayed tumor formation in vivo
We confirmed the anti-CSC effect of BC in vivo using the mouse xenograft model. In the BC 6 and BC 12 groups, we found that 6 out of 12 tumors had developed, while 9 out of 12 tumors had formed in the TC group at the time of sacrifice (Table 1). BC supplementation tended to inhibit tumorigenesis by 33% compared to the TC group, but the difference of in tumor incidence was not statistically significant.

Table 1
BC suppresses tumor incidence and final tumor volume1)
| Treatment group | Tumor incidence | Final tumor volume (mm3) |
|---|---|---|
| TC | 9/12 | 348.5 ± 91.3a |
| BC 6 | 6/12 | 145.0 ± 43.8b |
| BC 12 | 6/12 | 117.8 ± 31.2b |
TC, tumor control; BC, β-carotene; BC 6, tumor injection + BC 6 mg/kg b.w.; BC 12, tumor injection + BC 12 mg/kg b.w.
1)Number of mice with tumor incidence and final volume of mice injected with CD133+CD44+ HCT116 cells. Comparison among groups were conducted by χ2 (P < 0.05). The letter labels on the bar graph indicate the values that significantly differed from each other (P < 0.05) according to one-way analysis of variance for multiple comparisons.
![]()
The tumor volume was significantly reduced by BC supplementation at 6 and 12 mg/kg BW. The mean tumor volume was 348.5 ± 91.3 mm3 in group TC, 145.0 ± 43.8 mm3 in group BC 6, and 117.8 ± 31.2 mm3 in group BC 12 (Table 1), corresponding to tumor volume reductions of 58.4% (P < 0.05) and 66.2% (P < 0.05), respectively compared to the TC group. In addition, BC supplementation delayed tumor formation. Both BC-supplemented groups formed tumors more slowly than the TC group (P < 0.05; Fig. 4A).
 | Fig. 4BC supplementation suppressed tumor formation and expression of CSC markers in vivo. Balb/c nu/nu mice were pretreated with BC for 3 weeks and CD133+CD44+ HCT116 cells were injected subcutaneously for 7 weeks with BC feeding. (A) Tumor latency was measured. (B) mRNA expressions of CSC markers, CD44, CD133, ALDH1A1, and NOTCH1, were detected using real-time polymerase chain reaction. (C) Protein expressions of Notch1, SOX2, and β-catenin in tumors were detected by western blot. β-actin was used as a loading control.TC, mice were fed control diet (tumor control); BC, β-carotene; BC 6, tumor injection + BC at 6 mg/kg BW; BC 12, tumor injection + BC at 12 mg/kg BW.
*,†The labels on the bar graph indicate the values that significantly differed from each other (P < 0.05) by one-way analysis of variance for multiple comparisons.
|
BC supplementation suppressed expression of CSC markers and Wnt/β-catenin signaling in vivo
Furthermore, the mRNA levels of CSC markers were analyzed from isolated tumors (Fig. 4B). The mRNA expression level of CSC markers, including CD44, CD133, ALDH1A1, and NOTCH1, was suppressed by BC supplementation at both concentrations. In particular, BC supplementation at 12 mg/kg BW down-regulated these genes by 64.9% (P < 0.01), 58.3% (P < 0.05), 94.3% (P < 0.01), and 67.8% (P < 0.01), respectively, compared to the TC group. BC supplementation also suppressed the protein expressions of CSC markers, including Notch1 and Sox2, and β-catenin in the CSC signaling pathway, compared to the TC group (Fig. 4C).
Go to :
DISCUSSION
In the present study, BC suppressed self-renewal capacity by decreasing colony formation and sphere formation. In addition, BC down-regulated the mRNA expressions of CSC markers, including CD44, CD133, ALDH1A1, and NOTCH1 in CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cells. CSC markers and the protein expressions of Notch1, Sox2, and β-catenin in Wnt/β-catenin CSC signaling pathway were suppressed in BC-treated human primary cells isolated from tumors of CRC patients. Similar trends were seen in mice. Moreover, the number and size of tumors were decreased, and the tumor formation was delayed by BC supplementation in the xenograft mice model. This study is the first to identify the effects of BC on CSCs in CD133+CD44+ colon CSC cells and the CRC xenograft mouse model.
CD133+CD44+ cells were isolated from HCT116 and HT-29 CRC cell lines and used in the present study. CD133 is a type 1 transmembrane glycoprotein and was first shown to be a marker of colon CSC in immunodeficient mouse models [21]. CD133+ cells showed increased and more aggressive tumorigenicity with fewer cells CD133− cells [22]. CD133+ tumors were resistant to chemotherapy in CRC and were associated with a poor prognosis due to metastasis [2324]. As a transmembrane glycoprotein receptor, CD44 plays a critical role in cancer progression, especially in cell attachment, invasion, and migration [25]. Like CD133 cells, CD44+ cells also exhibited increased colony-forming ability and tumorigenicity in immunodeficient mouse models compared to CD44− cells [26]. The detection of CSCs with 2 CSCs markers increased tumor forming ability and was more reliable than the single marker in detecting human CRCs. For example, CD133+CD44+ populations could form tumors, but not CD133−CD44−, CD133+CD44−, and CD133−CD44+ populations [27]. Previously, we reported that only 10,000 CD133+CD44+ HT-29 colon cells could form tumors in the xenograft mouse model [15], and 1 × 105 CD133+CD44+ HCT116 cells could form tumors within little more than 2 weeks in the present study.
Clonogenic assays and sphere formation assays were used to identify the effect of BC on the self-renewal capacity of colon CSCs in vitro. The ability to form colonies and spheres from single cells is positively related to the number of self-renewing cells present [2]. BC treatment inhibited colony and sphere formations in both CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cells, consistent with that seen in undifferentiated neuroblastoma stem-like cells [14]. Furthermore, BC treatment down-regulated the mRNA expression of CD44, CD133, ALDH1A1, and NOTCH1 and protein expression of Notch1 and Sox2 in the present study. ALDH1A1 is an aldehyde hydrogenase with a role in cellular detoxification and supports tumor growth of CSCs. When ALDH1A1 is expressed in CRC tissues, cancers become malignant and show poor prognosis [28]. Notch plays an important role in the cell-fate determination of stem cells. Notch activity was 10–30 fold higher in cancer-initiating cells than in normal colon cancer cell lines. Notch prevented CSC apoptosis by repressing a transcriptional factor, ATOH1 and cell cycle kinase inhibitor, p27 [57]. Moreover, the Wnt/β-catenin signaling pathway regulates the growth and maintenance of clonospheres and the functional integrity of CSCs [729]. Numerous studies have demonstrated the association between the inhibition of the Wnt/β-catenin signaling pathways and the anti-CSC effect of natural compound. For example, sulforaphane and curcumin have been shown to regulate Wnt/β-catenin signaling and modulate self-renewal capacity in colon and breast CSCs [3031].
Although cancer cell lines are widely used for tumor biology research, they present some limitations. For instance, they may not accurately recapitulate the characteristics of original tumor cells because while primary cancers grow in 3 dimensions and under partially hypoxic conditions, cancer cell lines grow in a controlled experimental environment, which may lead to genetic, phenotypic, and metabolic differences between cancer cell lines and real tumors [32]. In the present study, primary cells isolated from human CRC were used to confirm the anti-CSC effect of BC shown in the isolated cancer cell lines. BC treatment suppressed the mRNA expressions of CSC markers, including CD44, CD133, and ALDH1A1 in primary cells from human CRC, consistent with the results from both CD133+CD44+ HCT116 and CD133+CD44+ HT-29 cell lines. These results indicate that CD133+CD44+ cells from those cell lines can represent colon CSCs and were appropriate models to investigate the anti-CSC effect of BC. In a recent study, BC was shown to exert anti-CSC potential by regulating the expression of histone acetylation-related miRNAs, H3 and H4 acetylation, and global DNA methylation in colon CSCs [20].
The anti-colon cancer effects of BC have been demonstrated to various models, not only in the xenograft model of CSCs but also in the chemical-induced model. In the present study, BC supplementation (6 and 12 mg/kg BW) suppressed the growth of colon tumors and expression of CSC markers in the tumors of the xenograft mice model. Previously, oral supplementation of 6 mg/kg BW of BC inhibited tumor formation by 60% in subcutaneously injected neuroblastoma xenograft model [13]. Moreover, BC supplementation (2 and 22 mg/kg BW) reduced tumor incidence by 50% in the DMH-induced colon cancer mice model [11]. BC supplementation of 6 and 12 mg/kg BW twice a week (1.7 and 3.4 mg/kg BW/day) is equivalent to about 6 and 12 mg/day for a 70-kg person, respectively [33]. Considering that 7–17 mg of BC exists in 100 g of orange-colored carrot (Daucus carota L.) [34], the dosages of BC used in the present study were in the physiological ranges.
Although BC has shown potential anticancer effects in several cancers, including lung cancer [35], prostate cancer [36], and neuroblastoma [13], there is a lack of clinical evidence of the anti-colon cancer effect of BC. BC intakes in most previous clinical studies were 1 to 6 mg/day [37]. Dose differences and absorption discrepancies between humans and mice can lead to discrepant results. It is also important to consider that the anticancer mechanisms of BC in the chemical-induced or xenograft model may differ from those in naturally occurring tumors in clinical studies. Therefore, further studies with various study designs, doses, and induction methods of tumors need to be examined. Notably, the β-Carotene and Retinol Efficacy (CARET) Trial [38] and the α-Tocopherol, β-Carotene Cancer Prevention (ATBC) Study [39] revealed that high-dose BC supplementations increased the risk of lung cancer in smokers and asbestos workers. Thus, the use of BC supplementation for cancer prevention should consider the heterogeneity among the participants, including the oxidative status of the subjects.
Differentiation is a common characteristic of both normal stem cells and CSCs. Once CSCs are differentiated, they exhibited less malignant and differentiated normal stem cells constitute colon tissues [40]. Therefore, differentiation can be an excellent strategy for treating cancer. However, BC did not affect the colon CSCs differentiation markers, including cytokeratin 20 and CDX1 in the present study (data not shown).
The present study found that BC inhibited colon CSCs by suppressing the self-renewal capacity of CSCs, down-regulating the expression of CSCs markers, including CD44, CD133, ALDH1A1, NOTCH1, and Sox2, as well as the Wnt/β-catenin signaling pathway. Taken together, the results suggest that BC is a potential therapeutic agent that targets colon CSCs.
Go to :
 XML Download
XML Download