

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Alzheimer's disease (AD) is one of the most representative disease of various chronic degenerative diseases with clinical symptoms such as memory loss, speech impairment, and cognitive dysfunction [1]. According to the World Health Organization, more than 30 million people have AD worldwide, and its prevalence is expected to reach over 106 million by 2050 [2]. Although the cause of AD is multifactorial, evidence suggests comprehensive risk factors for AD, including age, heredity, and sex [3]. In addition, environmental factors such as air pollution, noise, and lack of greenness have emerged as reasonable risk factors for cognitive decline related to AD [4]. The pathogenesis of AD has been reported to be complex, but the most widely known cause is excessive production of amyloid beta (Aβ) [5]. Aβ is produced through the amyloidogenic pathway, which is a mechanism by which amyloid precursor proteins are degraded by enzymes such as β-secretase and γ-secretase [6]. Once Aβ accumulates in the brain, it causes oxidative stress, which leads to lipid peroxidation in neuronal cell membranes, leading to the generation of free radicals and reactive oxygen species (ROS) such as hydroxyl radicals, superoxide, and hydrogen peroxide [7]. The intracellular production of free radicals and ROS mediates apoptotic signaling pathways that lead to spontaneous cell death [89]. As a result, neuronal damage and death induced by oxidative stress can cause cognitive impairment in AD [10]. Thus, inhibition of oxidative stress is considered an important mechanism for the prevention and treatment of AD. Currently, the AD medications used in the clinic are aimed at delaying the progression of AD or alleviating symptoms rather than fundamentally treating AD. In addition, it is necessary to develop a material derived from natural products with few side effects because problems such as resistance and toxicity have been reported when AD medication is administered long term [11].
Populus tomentiglandulosa (PT), which is widely distributed in Korea, is a plant belonging to the Salicaceae family; PT is artificially developed through the crossbreeding of P. davidiana and P. alba [12]. Phenolic compounds and flavonoids are the main components of Populus extracts and are involved in various physiological activities [13]. Furthermore, many studies have reported that Populus species, including P. nigra, P. alba, and P. davidiana, showed several pharmacological activities such as antioxidant, anti-inflammatory, and protective effects against aluminum-induced hepatic toxicity [141516]. To date, most studies on PT were mainly related to its use as timber or an ornamental plant, and several studies on physiological activity have reported antioxidant activity and protective effects against cerebral ischemia-induced neuronal damage. The PT extract showed 1,1-diphenyl-2-picrylhydrazyl (DPPH) and azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) radical scavenging activities in vitro and significantly increased antioxidant enzyme levels, such as those of superoxide dismutase 1, superoxide dismutase 2, catalase, and glutathione peroxidase, in rats [1317]. In addition, PT extract has been reported to inhibit neuronal loss and alleviate gliosis in the gerbil hippocampal CA1 area induced by transient global cerebral ischemia [18]. Therefore, PT is expected to have a neuroprotective effect on the brain through the inhibition of oxidative stress. However, the effect of PT on cognitive dysfunction in AD has not yet been studied.
Based on this background, we hypothesized that PT has a protective effect on AD through the inhibition of oxidative stress. In the present study, we investigated the protective effect of the ethyl acetate (EtOAc) fraction of PT against AD using an Aβ25-35-induced mouse model. Space perceptive ability, object cognitive ability, and long-term memory were evaluated through behavioral tests to assess the cognitive ability. Moreover, the protective mechanisms of PT against AD were investigated.
Go to : 
MATERIALS AND METHODS
Reagents
Aβ25-35, donepezil (DO), Griess reagent, malondialdehyde (MDA), and 2’,7’-dichloro-fluorescein diacetate (DCF-DA) were obtained from Sigma-Aldrich Co. (St. Louis, MO, USA). Sodium chloride (NaCl) was purchased from LPS Solution Co. (Daejeon, Korea). Trichloroacetic acid (TCA) and thiobarbituric acid (TBA) were obtained from Biosesang Inc. (Seongnam, Korea) and Acros Organics Inc. (New Jersey, USA), respectively. n-Butanol and acetic acid were acquired from Duksan (Ansan, Korea). Pyridine was purchased from Wako Pure Chemical Co. (Osaka, Japan). Dimethyl sulfoxide (DMSO) was purchased from Biopure Reagent Co. (Daejeon, Korea). Phosphoric acid was acquired from Samchun Pure Chemical Co., Ltd. (Seoul, Korea).
Sample preparation and HPLC
The dried leaves of PT were extracted with ethanol under reflux three times (86°C, 3 h), and the extracts were combined and evaporated to yield a brown residue. They were then suspended in distilled water and partitioned in turn using organic solvents. HPLC analysis was performed in an isocratic elution HPLC system (Waters 2489, Waters 1525 Binary HPLC, MA 01757, USA) using a reverse-phase YMC-Pack Pro C18 column (4.6 × 150 mm, 5 μm) with UV detection at 267 nm.
Animals and experimental protocols
Five-week-old male ICR mice weighing 25–27 g were purchased from Orient Inc. (Seongnam, Korea). Experimental mice were housed in plastic cages and with a controlled temperature (20 ± 2°C) and humidity (50 ± 10%) under a 12 h light/dark cycle; they were provided with normal pellet diets and water. The animal protocol used in this study was approved by the Ethics Committee (No. PNU-2020-2536) of the Pusan National University Institutional Animal Care and Use Committee (PNU-IACUC). After a week of adaptation, the mice were divided into five groups (n = 7 in each group), and there was no significant difference in body weight between the groups.
Aβ25-35 was dissolved at a concentration of 25 nM/5 μL in 0.9% NaCl and incubated at 37°C for three days. The mice were then injected with an anesthetic mixture of zoletil and rumpun through the abdominal cavity, followed by intracerebroventricular (i.c.v.) injection [19]. The anesthetized mice were placed on a stereotaxic apparatus, and Aβ25-35 or 0.9% NaCl was injected a rate of 1 μL/min using a 25 μL glass syringe after drilling the brain bone using a manual hand drill at an anterior/posterior distance of −0.8 mm, medial/lateral of 1.5 mm, and dorsal/ventral of 2.2 mm from the bregma. After four days, oral administration of PT was conducted for 14 days using zonde gavage. The experimental schedule is illustrated in Fig. 1, and i.c.v. injection and oral administration for each group were as follows: normal group = i.c.v. injection of 0.9% NaCl + oral administration of drinking water; Control group = i.c.v. injection of Aβ25-35 (25 nM/5 μL) + oral administration of drinking water; PT50 group = i.c.v. injection of Aβ25-35 (25 nM/5 μL) and oral administration of the EtOAc fraction of PT (50 mg/kg/day); PT100 group = i.c.v. injection of Aβ25-35 (25 nM/5 μL) + oral administration of EtOAc fraction of PT (100 mg/kg/day); DO group = i.c.v. injection of Aβ25-35 (25 nM/5 μL) + oral administration of DO (5 mg/kg/day). The day after completion of the behavioral tests, the mice were anesthetized with zoletil and rompun mixture intraperitoneally, and then the abdomen was opened to collect organs such as the liver and kidney. The whole brain was used for the experiments.
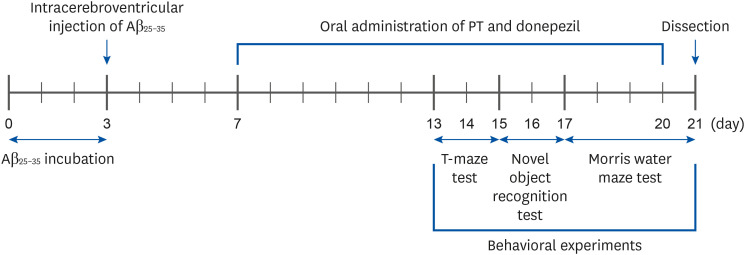
T-maze test
The T-maze test was carried out in accordance with the method described by Kim et al. [20]. The structure of the apparatus was T-shaped, which consisted of the right and left arms with removable doors and stems. The wall and bottom of the apparatus were made of black plastic board (start and target stem, 50 cm; width, 13 cm; height, 20 cm). On the training day, after removing only the door of the right arm, the mice were placed at the start point of the apparatus. The mice freely navigated between the start point and the right arm for 10 min, and the number of times the mice entered the right arm was recorded. Twenty-four hours later, on the test day, the door of the left arm was also removed, and mice were placed at the start point of the apparatus. The mice were allowed to explore the inside of the apparatus and enter both arms, and the number of times the mice entered the right arm (old arm) and left arm (new arm) was recorded. Using these results, the space perceptive ability was calculated as follows: the space perceptive ability (%) = (the number of times of entering the right arm/the number of times of entering the right arm and left arm) × 100.
Novel object recognition test
The novel object recognition test was performed following the steps of Bevins and Besheer [21]. On the day of training, the mice were placed at the center of a 40 × 40 × 40-cm square black plastic box with two identical objects fixed on the floor. For 10 min, the mice roamed freely inside the box, exploring the two objects; the number of times the mice touched both objects was recorded. Twenty-four hours later, on the test day, one of the two identical objects was replaced with another of a similar size. The mice were placed in the center of the box for 10 min, and the number of times the mice touched each object was counted. Using these results, the object cognitive ability was calculated as follows: The object cognitive ability (%) = (the number of times of touching the novel object/ the number of times of touching the novel object and familiar object) × 100.
Morris water-maze test
The Morris water-maze test was performed in accordance with the method of He et al. [22]. The apparatus used in the maze test was a round-shaped water tank of stainless steel with a diameter of 95 cm and height of 45 cm that was evenly divided into quadrants at the center point of the circle. An 8 cm-diameter platform was placed in the center of one quadrant, and the surface of the water remained 1 cm higher than the platform. The platform position remained unchanged during the experiment. The black poster paint was dissolved in water so that the platform was not visible, and the water temperature was maintained at 22 ± 2°C. Each visual cue was attached to the wall of the tank corresponding to each quadrant, and the mouse was able to find the platform by looking at these four cues. The tracking path was recorded with the camera using the SMART video tracing software (version 3.0; Panlab, Spain) to analyze the movement of the mice. The duration of the experiment was four days in total, and the first three days were training sessions, and the last one day was a test session. During the 3-day training period, training was conducted three times at 4-h intervals each day, and mice were placed randomly in the center of the remaining three quadrants, excluding the one on which the platform was located. The mice were allowed to swim freely in the tank for 60 s, and the tracking pathway and latency time to reach the hidden platform were recorded. When the mice reached the platform within 60 s, the mice stayed there for 10 s, and if the mice did not reach the platform within 60 s, they were guided to reach the platform and then stayed there for 10 s to remember the platform's location. On day 4 of the experiment, on the test day, three different tests were conducted at 4-h intervals. The first test was conducted in the same way as the training session, but when the mice reached the platform or after 60 s, the mice were immediately removed without training. In the second test, after the platform was removed, the time the mice stayed in the target quadrant was recorded for 60 s. In the final test, the water in the tank was transparent, and the platform was repositioned in the tank and exposed above the water surface. The latency time to reach the exposed platform was recorded for 60 s.
Measurement of hepatic toxicity
To evaluate hepatic toxicity, the serum levels of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were analyzed using glutamic oxaloacetic transaminase (GOT) and glutamic pyruvate transaminase (GPT) assay kits (AM103-K and AM102; Asan Pharm., Seoul, Korea) according to the method of Jang and Choung [23].
Measurement of lipid peroxidation
MDA levels in tissues were determined according to the procedure of Kim et al. [24]. The brains, livers, and kidneys of the mice were homogenized with 0.9% NaCl. The homogenized solution was centrifuged at 3,000 rpm for 10 min at 4°C, and the supernatant was collected. The supernatant was added to a solution of thiobarbituric acid reactive substances mixed with 20% acetic acid, TBA, and 15% TCA, and the mixture was boiled for 20 min. Immediately after boiling, the mixed solution was cooled on ice, added to a solution of n-butanol and pyridine in a 15:1 ratio, and centrifuged at 3,000 rpm for 20 min at 4°C. The absorbance value of the supernatant was measured at 540 nm using a microplate reader (Thermo Fisher Scientific, Vantaa, Finland), and lipid peroxidation levels were presented as MDA equivalents through the calculation of the standard curve.
Measurement of nitric oxide (NO) generation
NO generation in tissues was evaluated using the method described by Lee et al. [25]. The homogenized tissue supernatant was diluted with distilled water, and the Griess reagent was added in the same amount as the diluent. The mixed solution was incubated at room temperature for 30 min, and the absorbance was measured at 540 nm using a microplate reader. The level of NO generation was presented as NaNO2 equivalents by calculating the standard curve.
Measurement of ROS production
ROS production was measured using the DCF-DA assay [26]. The homogenized tissue supernatant was diluted with 50 mM phosphate buffered saline (pH 7.4) and added to 12.5 mM DCF-DA solution dissolved in DMSO. The fluorescence value was read at an excitation wavelength of 480 nm and an emission wavelength of 535 nm for 60 min using a fluorescence spectrophotometer (FLUOstar OPTIMA, BMG Labtech, Ortenberg, Germany).
Statistical analysis
The data are expressed as mean ± SD. The statistical significance of the values was analyzed using the IBM SPSS version 23 program (IBM Corporation, NY, USA) by one-way analysis of variance, followed by Duncan's multiple range test and Student's t-test. Statistical significance was set at P-values less than 0.05.
Go to :
RESULTS
Extraction yield and HPLC analysis
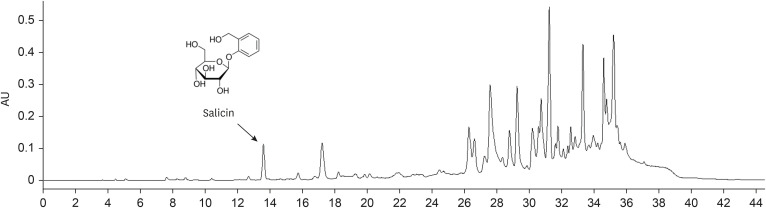
The dried leaves of PT (1,442.6 g) were extracted with ethanol under reflux three times (86°C, 3 h), and the extracts were combined and evaporated to afford a brown residue (278.3 g). Then, it was suspended in distilled water and partitioned in turn using n-hexane (45.7 g), chloroform (35.5 g), EtOAc (36.0 g), and n-butanol (59.9 g). Salicin was isolated from the EtOAc fraction of PT (data not shown) and was confirmed by HPLC analysis (Fig. 2).
Effect of PT on the space perceptive ability in the T-maze test
To test the effect of PT on the space perceptive ability in Aβ25-35-induced mice, a T-maze test was conducted. In the normal group, the percentages of entry into old and new routes were 38.02% and 61.98%, respectively (Fig. 3). In contrast, the control group had 51.08% and 48.82%, respectively, with no significant difference in the percentage of old and new routes. This result showed that the spatial perceptive ability of mice was impaired by Aβ25-35 injection. However, in the PT50 and PT100 groups, the percentage of entry into the new route was 62.23% and 66.18%, respectively, and it increased significantly compared with that of the control group. In addition, the percentage of entries into new routes in the DO group was 60.54%. These results indicated that PT improved the spatial perceptive ability to a level similar to that of DO in the Aβ25-35-induced AD mouse model.
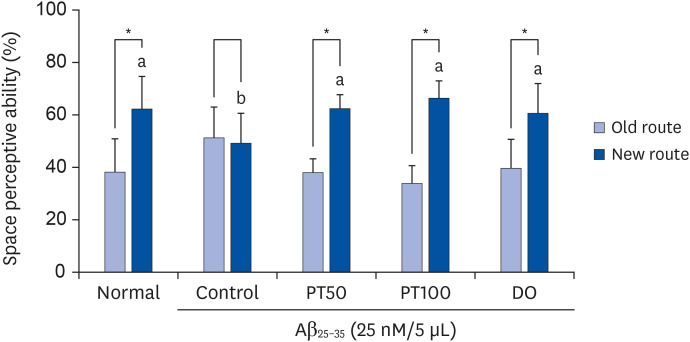 | Fig. 3Effect of EtOAc fraction from PT on space perceptive ability in the T-maze test.The results are expressed as the mean ± SD. The asterisk (*) indicates that the space perceptive abilities for old and new routes were significantly different, as determined by Student's t-test (P < 0.05). Different letters (a, b) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test (n = 7). Normal = 0.9% NaCl i.c.v. injection + drinking water; Control = Aβ25-35 i.c.v. injection (25 nM/5 μL) + drinking water; PT50 = Aβ25-35 i.c.v. injection (25 nM/5 μL) + oral administration of the EtOAc fraction from PT (50 mg/kg/day); PT100 = Aβ25-35 i.c.v. injection (25 nM/5 μL) + oral administration of the EtOAc fraction from PT (100 mg/kg/day); DO = Aβ25-35 i.c.v. injection (25 nM/5 μL) + oral administration of donepezil (5 mg/kg/day).
PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta; EtOAc, ethyl acetate.
|
Effect of PT on the object cognitive ability in the novel object recognition test
A novel object recognition test was conducted to evaluate the effect of PT on cognitive ability. As shown in Fig. 4, the percentage of mice in the normal group touching the novel object was 63.09%, which was significantly higher than the touching percentage of familiar objects (36.91%). In contrast, the control group had values of 48.41% and 51.59%, respectively, and there was no significant difference between the percentage of familiar and novel objects, indicating that the object cognitive ability of mice was impaired in the control group. In contrast, the PT50 and PT100 groups were 55.17% and 59.41%, respectively, indicating that PT significantly increased the recognition ability toward novel objects. These results showed that PT had a protective effect on the cognitive ability of Aβ25-35-induced mice.
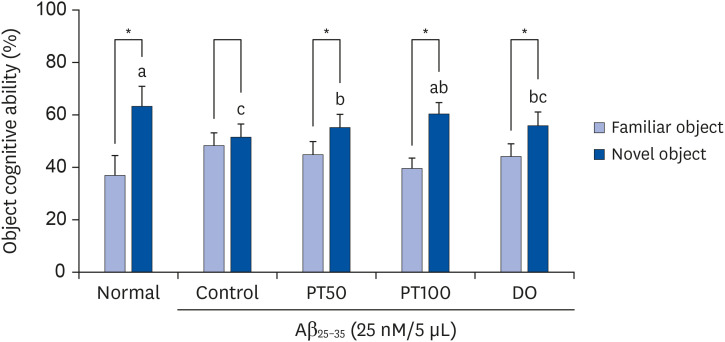 | Fig. 4Effect of ethyl acetate fraction from PT on objective cognitive ability in novel object recognition test.The results are expressed as mean ± SD. The asterisk (*) indicates that the object cognitive abilities for familiar and novel routes are significantly different, as determined by Student's t-test (P < 0.05). Different letters (a–c) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test (n = 7). The mice were grouped and treated as described in Fig. 3.
PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta.
|
Effect of PT on Morris water-maze test
The Morris water-maze test was conducted to investigate spatial learning and long-term memory in an Aβ25-35-induced mouse model. Fig. 5 shows the tracking pathway and the time required to reach the platform. During the experimental period, the control group showed a long and complicated pathway and arrived at the platform later than the normal group, suggesting that spatial and long-term memory was impaired by Aβ25-35. However, the PT50 and PT100 groups showed increasingly shorter and simpler pathways as the training progressed. In addition, the PT50 and PT100 groups showed reduced time to reach the platform compared to the control group. In particular, on the last day of the experiment, the PT50 and PT100 groups showed very short pathways and significantly decreased the time to reach the platform compared with the control group, which was similar to the normal and DO groups. In the second test, after the platform was removed, the percentage of time that the mice stayed in the target quadrant was evaluated. As shown in Fig. 6, the control group spent significantly less time in the target quadrant than the normal group. However, the PT50 and PT100 groups stayed longer in the target quadrant than the control group, and these results were not significantly different from those of the normal and DO groups. In the final test, the water in the tank was replaced to a transparent one, the platform was exposed, and the latency time to reach the platform was measured. As shown in Fig. 7, there was no significant difference in the time to reach the exposed platform among all groups, suggesting that exercise and visual abilities did not affect the experimental results.
 | Fig. 5Effect of ethyl acetate fraction from PT on representative tracking pathway (A) and escape latency (B) in the Morris water-maze test.The results are expressed as mean ± SD. Different letters (a, b) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test (n = 7). The mice were grouped and treated as described in Fig. 3.
PT, Populus tomentiglandulosa; DO, donepezil.
|
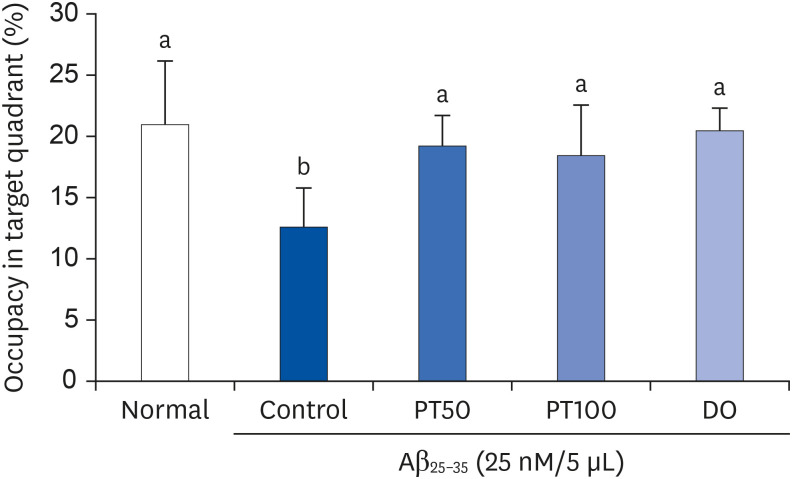 | Fig. 6Effect of ethyl acetate fraction from PT on the percent of time stayed in the target quadrant in Morris water-maze test.The results are expressed as mean ± SD. Different letters (a, b) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test (n = 7). The mice were grouped and treated as described in Fig. 3.
PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta.
|
 | Fig. 7Effect of ethyl acetate fraction from PT on escape latency to hidden platform (A) and the exposed platform (B) in the Morris water-maze test.The results are expressed as mean ± SD. Different letters (a, b) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test. NS indicates no significant differences among groups (n = 7). The mice were grouped and treated as described in Fig. 3.
PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta.
|
Effects of PT on hepatic toxicity
To evaluate hepatic toxicity, serum levels of AST and ALT were measured. The serum ALT and AST levels in all experimental groups were within the normal range, and there were no significant differences among the groups (Fig. 8). These results indicate that PT did not induce liver toxicity.
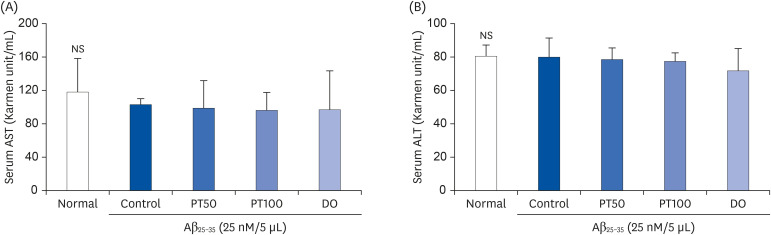 | Fig. 8Effects of ethyl acetate fraction from PT on AST (A) and ALT (B) in Aβ25-35-induced mice.The results are expressed as mean ± SD. NS indicates no significant differences among groups (P < 0.05) by Duncan's multiple range test (n = 7). The mice were grouped and treated as described in Fig. 3.
AST, aspartate aminotransferase; PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta; ALT, alanine aminotransferase.
|
Effects of PT on lipid peroxidation
As shown in Fig. 9, in the brain, the MDA level of the control group (136.53 nmol/mg protein) was significantly higher than that of the normal group (103.89 nmol/mg protein). However, the MDA levels in PT50 and PT100 groups were 119.14 nmol/mg protein and 111.37 nmol/mg protein, respectively, which were significantly decreased compared to those in the the control group. In particular, the PT100 group showed significantly higher MDA levels than the DO group (123.10 nmol/mg protein), which showed that PT administration reduced MDA levels in the brain in a dose-dependent manner. The MDA levels in the liver were significantly increased in the control group (50.02 nmol/mg protein) compared to that in the normal group (32.78 nmol/mg protein). On the other hand, in the PT50 and PT100 groups, the MDA levels in the liver were significantly reduced to 40.31 nmol/mg protein and 40.88 nmol/mg protein, respectively, compared to the levels in the control group, which showed similar results to the DO group (42.55 nmol/mg protein). Moreover, the control group (107.79 nmol/mg protein) showed significantly higher MDA levels in the kidney than the normal group (81.15 nmol/mg protein). In contrast, MDA levels in the PT50 and PT100 showed a significant decrease to 90.39 nmol/mg protein and 81.49 nmol/mg protein, comparable with the levels in the normal and DO groups (88.98 nmol/mg protein). Thus, our findings showed that PT decreased lipid peroxidation in an Aβ25-35-induced AD mouse model.
 | Fig. 9Effect of ethyl acetate fraction from PT on lipid peroxidation in brain (A), liver (B), and kidney (C) in Aβ25-35-induced mice.The data are expressed as mean ± SD. Different letters (a–d) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test (n = 7). The mice were grouped and treated as described in Fig. 3.
MDA, malondialdehyde; PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta.
|
Effects of PT on NO generation
As shown in Fig. 10, the NO concentrations in the brain, liver, and kidneys were measured in Aβ25-35-induced mice. The concentration of NO in the brain was 39.90 μmol/mg protein in the control group and 24.75 μmol/mg protein in the control group and normal group, respectively; thus, the control group showed significantly higher NO concentration than the normal group. However, the NO concentrations in the brain of PT50 and PT100 groups were 29.02 μmol/mg protein, 31.77 μmol/mg protein, similar to that of the normal group. The NO concentration in the brain of the DO group was 33.38 μmol/mg protein, and there was no significant difference in the control group. The NO concentration in the liver in the control group was 79.59 μmol/mg protein, significantly higher than that in the normal group (57.87 μmol/mg protein. However, in the PT50 and PT100 groups, the NO concentrations in the liver were 61.18 μmol/mg protein and 60.92 μmol/mg protein, respectively, and they were significantly decreased compared to those in the control group. In the kidney, like the brain and liver, the control group (90.99 μmol/mg protein) showed significantly higher NO concentration than that in the normal group (78.17 μmol/mg protein), suggesting that Aβ25-35 injection increases NO production in tissues. On the other hand, PT50 and PT100 groups had 62.24 μmol/mg protein and 72.48 μmol/mg protein, respectively, significantly lower than that seen in the control group. Furthermore, the administration of PT decreased the NO concentrations in the brain, liver, and kidney to the same level or higher compared to DO. These findings demonstrated that treatment with PT decreased NO generation in the brain, liver, and kidney of Aβ25-35-induced mice.
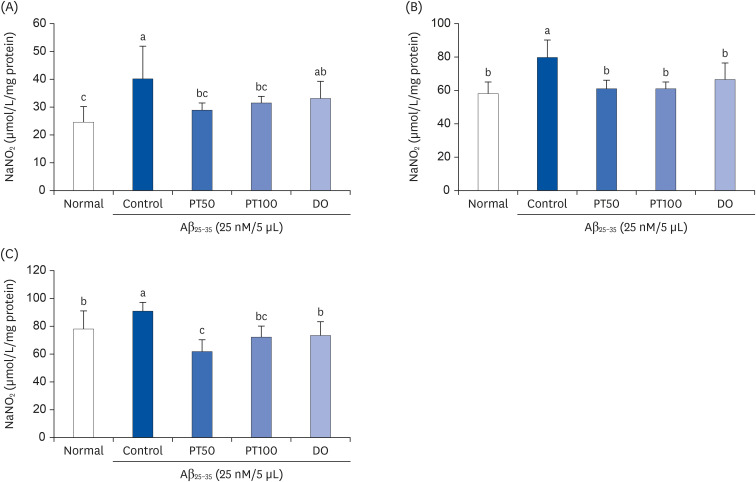 | Fig. 10Effect of ethyl acetate fraction from PT on NO generation in brain (A), liver (B), and kidney (C) in Aβ25-35-induced mice.The results are expressed as mean ± SD. Different letters (a–c) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test (n = 7). The mice were grouped and treated as described in Fig. 3.
PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta.
|
Effects of PT on ROS inhibition
ROS production in the brain, liver, and kidneys of Aβ25-35-induced mice was analyzed by fluorescence measurement. As shown in Fig. 11, the ROS production in the brain and liver was higher in the control group than that in the normal group. However, in the PT50 and PT100 groups, ROS production in the brain and liver was significantly decreased compared to that in the control group. In addition, in the brain, the PT50 group showed a significantly higher reduction in ROS production than the DO group. In the kidney, ROS production was significantly higher in the control group than in the normal group. However, the ROS production in the kidneys of the PT50 and PT100 groups decreased to the same extent as in the normal and DO groups. These data suggest that the administration of PT inhibited ROS production in Aβ25-35-induced mice.
 | Fig. 11Effect of ethyl acetate fraction from PT on reactive oxygen species production in the brain (A), liver (B), and kidney (C) of Aβ25-35-induced mice.The results are expressed as mean ± SD. Different letters (a–d) among groups indicate significant differences (P < 0.05) by Duncan's multiple range test (n = 7). The mice were grouped and treated as described in Fig. 3.
PT, Populus tomentiglandulosa; DO, donepezil; Aβ, amyloid beta.
|
Go to :
DISCUSSION
Overproduction of Aβ in the brain is known to induce ROS generation and oxidative stress, which are strongly associated with the pathogenesis of AD [2728]. Several mechanisms have been proposed to explain the relationship between Aβ and oxidative stress. It has been suggested that Aβ in the aggregated form may be inserted into the lipid membrane and directly generate ROS in mitochondria. Other mechanisms have proposed that Aβ may induce calcium influx, which breaks calcium balance or injures the mitochondrial respiratory chain, indirectly causing mitochondrial ROS production [29]. The ROS and free radicals produced through these mechanisms oxidize the cell components, including enzymes and organelles, eventually leading to neuronal damage and cognitive impairment [30]. In addition, Aβ consists of 39-43 amino acid residues, and its diverse fragments, such as Aβ1–28, Aβ25-35, and Aβ34–42, have biochemical and biophysical characteristics similar to those of the Aβ full-length peptide [3132]. Particularly, it has been demonstrated that the Aβ25–35 fragment is a functional area of Aβ that is responsible for its neurotoxicity characteristics [33]. Thus, Liu et al. [34] reported that i.c.v. injection of Aβ25–35 impaired cognitive ability associated with behavior such as spontaneous behavioral changes, passive avoidance, and water-maze learning ability in a mouse model [33]. They also suggested that injection of aggregated Aβ25–35 could cause more neuronal damage than the non-aggregated form. Therefore, based on these studies, we used aggregated Aβ25-35 to evaluate the AD mouse model in this study.
ICR (also known as Crl:CD1) mice, an albino strain, was named after the Institute of Cancer Research in the USA. The ICR mice were chosen to examine the improvement effects of PT under Aβ25-35-induced cognitive dysfunction not only in behavioral analyses but also in antioxidant activity analyses. The mice are one of the most frequently used outbred strains in research on toxicology, aging, and pharmacology because they have been described as docile, robust, and easy to handle [35]. Hsieh et al. [36] reported that ICR mice were as suitable as the most popular inbred strain, C57BL/6J, for evaluating behavior. Moreover, many studies have used ICR mice to explore the functional effects of natural materials under Aβ-induced cognitive dysfunction. Lee et al. [37] reported that black rice supplementation improves memory deficits induced by Aβ25-35 stimulation in ICR mice. Cho et al. [38] showed that Bidan kiwifruit has potential preventive efficacy in Aβ1-42-induced ICR mice.
Donepezil (DO) was used as the positive control. DO is a representative compound used for AD treatment in many countries because it is a powerful and specific inhibitor of acetylcholinesterase (AChE) [3940]. AChE acts as a rapid hydrolytic enzyme of the neurotransmitter acetylcholine (ACh) at cholinergic synapses, which can cause neuronal damage and cognitive impairment in AD. Therefore, AChE inhibition is closely related to symptomatic treatment of AD, and research on AChE inhibitors has been actively carried out [41]. In addition, previous studies on DO have indicated that: (1) DO provides protection against free radical-mediated neuronal degeneration in AD mouse brains [42]; (2) DO improves neuronal loss and cognitive dysfunction by suppressing the Aβ-induced inflammatory pathways in AD patients, a tauopathy mouse model, and LPS-treated animals [43]. Based on these studies, DO is used for AD through the inhibition of AChE and regulation of oxidative stress. PT also showed a protective effect against Aβ25-35-induced oxidative stress, which is related to the improvement of cognitive function.
We previously measured the antioxidant activity in vitro and its neuroprotective effect in a cellular system using ethanol extract and four fractions (n-butanol, EtOAc, chloroform, and n-hexane) of PT (data not shown). The results showed that the EtOAc fraction of PT had the strongest antioxidant and neuroprotective effects among the other extracts and fractions. Therefore, in reference to these studies and our results, we used the EtOAc fraction of PT to determine its protective effect on AD. The EtOH extract of PT contains catechin (9.1 ± 0.27 mg/g), caffeic acid, p-coumaric acid, chlorogenic acid, and gallic acid [44]. Many studies have demonstrated that catechin has antioxidative and neuroprotective effects [444546]. In the present study, salicin was isolated from the EtOAc fraction of PT (data not shown) and confirmed by HPLC analysis. We orally administered the EtOAc fraction of PT and then conducted behavioral tests such as the T-maze test, novel object recognition test, and Morris water-maze test to investigate the effect of PT on cognitive abilities. These three tests were used for assessment of the following: T-maze and novel object recognition to assess short-term cognitive ability, and Morris water-maze to assess long-term cognitive ability.
The T-maze test has been most widely used to evaluate spatial working memory ability, and it is useful to rodents during the current experience with the task [47]. Meanwhile, it has been reported that AD patients initially develop clinical symptoms, such as spatial memory loss, due to neuronal synaptic dysfunction [48]. Therefore, many studies have used the T-maze test to investigate the spatial memory loss in AD [4950]. In the T-maze test, mice are placed at the start point of the T-shaped apparatus and can choose between the right and left routes. According to spontaneous alternation, in the second trial, the mice tended to choose the new route more, reflecting former memory [51]. Lee et al. [52] demonstrated that the Aβ25-35-treated control group did not show a preference for new routes, suggesting that spatial cognitive impairment was caused by Aβ25–35 injection. Consistent with these previous reports, in our study, there was no significant difference in the percentage of old and new routes in Aβ25–35-induced mice. However, the administration of PT and DO significantly increased the percentage of treated mice choosing the new route compared to those in the control group. DO is a representative compound used for AD treatment in many countries and is widely used as a positive control in studies on AD treatment [3940]. Therefore, our findings indicate that PT has a protective effect on spatial cognitive impairment induced by Aβ25–35 at a level similar to that of DO.
The novel object recognition test is used to study the mechanisms or therapeutics of AD because it is a very useful method to assess the non-spatial learning ability of objects that depend on multiple brain regions in rodents [5354]. In the novel object recognition test, mice were allowed to explore two objects freely. Because mice instinctively prefer novelty, they spend most of their time exploring the novel object after recognizing a familiar objects [55]. According to Lu et al. [56], Aβ25–35-induced mice did not show significant differences in the percentage of exploring familiar and novel objects, indicating that the object cognitive ability was impaired by Aβ25–35. Based on the results in this study, the Aβ25–35-induced control group showed no significant difference between the ratio of exploring familiar and novel objects. On the other hand, PT treatment significantly increased the ratio of exploring novel objects, and these findings suggested that PT protected against Aβ25–35-induced cognitive impairment in mice.
The Morris water-maze test evaluates hippocampal-dependent learning ability, including spatial and long-term memory, in rodents that rely on visual cues to find escape platforms [5758]. Many studies have used the Morris water-maze test to investigate the learning and long-term memory abilities of AD mouse models [5960]. Kwon et al. [61] reported that Aβ25-35-injected mice did not show reduced time to reach the platform and also that they stayed significantly shorter in the target quadrant compared to normal mice. These results indicated that the spatial and long-term memory of mice was impaired by Aβ25-35. Similarly, our results showed that the Aβ25-35-induced control group reached the platform significantly late and stayed the shortest time in the target quadrant compared to the other groups. In contrast, administration of PT and DO significantly decreased the time to reach the platform and significantly increased the time spent in the target quadrant compared with the control group. Consequently, our data showed that PT improved spatial and long-term memory deterioration by Aβ25-35 injection to a degree similar to DO.
AST and ALT are enzymes that exist primarily in the liver; they also exist in red blood cells, muscle tissue, heart, pancreas, and kidney. However, if the body tissue or organs such as the liver or heart are damaged, AST and ALT are released into the bloodstream, which increases the level of enzymes in the blood. Therefore, the levels of AST and ALT in serum are directly associated with the extent of tissue injury, especially hepatic injury [62]. Based on the safety and effect of PT, we determined the doses of 50 and 100 mg/kg/day. In previous studies, doses of 100 and 200 mg/kg/day of PT were administered in a gerbil model for transient global cerebral ischemia [6364]. This demonstrated that hippocampal neurons were protected from cerebral ischemia after treatment with 200 mg/kg/day PT extract. There was no significant toxicity data at concentrations of PT treatment above 150 or 200 mg/kg/day. In order to determine doses that are not toxic and safe, we used lower doses (50 and 100 mg/kg/day) than those used in previous studies. In addition, we analyzed the serum levels of AST and ALT to evaluate hepatic toxicity. Our findings showed that ALT and AST levels in the serum of all experimental groups were within the normal range, and there were no significant differences among all groups. These results suggest that PT treatment does not cause hepatic toxicity. Regarding the other toxic potentials of PT, Lee et al. [65] reported that the ethanol extract of PT might not cause hepatotoxicity or nephrotoxicity. On the other hand, it showed hepatoprotective and nephroprotective effects. In addition, the ethanol extract from PT protected pyramidal neurons in the cornu ammonis 1 from transient global cerebral ischemia, indicating the neuroprotective effect of PT [6364].
Clinically, in the brains of AD patients, aberrant accumulation of Aβ plaques was primarily observed and almost consistently increased [66]. Histological analysis of the cresyl violet-stained brain indicated relevant cell death in the frontoparietal cortex and hippocampal sections of Aβ25-35-injected mice. Additionally, evaluation of the Congo red-stained brain in the same mouse model showed aggregation of Aβ peptides throughout the brain [67]. Although the present study did not include histological data from Aβ25-35-triggered mice, the regulatory effect of Aβ accumulation in the brain might be related to the histopathological changes in the brain of AD by PT. Further studies are required to verify the beneficial effects of PT against Aβ25-35-stimulated histological damage. Moreover, we previously analyzed Aβ accumulation in the brain (data not shown). PT treatment significantly reduced the expression of Aβ-production-related indicators.
As mentioned previously, Aβ plays a critical role in the pathogenesis of AD. Previous studies have shown that brain-derived Aβ circulates in peripheral tissues and organs [68]. Xiang et al. [69] reported that 125I-labeled Aβ40 was injected in the liver and kidney; consequently, brain-derived Aβ induces oxidative stress in Aβ-located organs such as the liver and kidney. Therefore, we analyzed the organs, including the brain, liver, and kidney, to study the effect of PT on AD.
Many studies have demonstrated that Aβ induces oxidative stress and causes lipid peroxidation in brain cell membranes [70]. Therefore, lipid peroxidation is a crucial mechanism for the neurodegeneration of AD patients, and there are reports that the brains of AD patients have higher levels of lipid peroxidation than that in age-matched controls [71]. MDA is widely used to evaluate the extent of lipid peroxidation because it is one of the by-products generated from the oxidation of unsaturated fatty acids [72]. Based on these results, previous studies have demonstrated that intravenous injection of Aβ25–35 significantly increased MDA levels in mouse tissue [73]. Moreover, in our study, the MDA levels in the brain, liver, and kidney showed that the Aβ25-35-induced control group was significantly higher than that in the normal group, indicating that Aβ25-35 caused oxidative stress and consequently led to lipid peroxidation in mouse tissue. Nevertheless, oral administration of PT significantly decreased the extent of lipid peroxidation compared to that in the control group, and these results were similar to those of DO. These results suggested that PT could effectively protect against oxidative stress and lipid peroxidation induced by Aβ25-35.
Aβ upregulates the expression of inducible nitric oxide synthase mRNA, which increases NO production [74]. NO, a type of reactive nitrogen species, has unstable free radical properties that cause oxidative stress [75]. Moreover, many studies have demonstrated that oxidative stress induced by NO is closely related to neuronal apoptosis pathways [7677]. Therefore, NO plays a critical role in the pathogenesis of AD because of its neurotoxicity [75]. Díaz et al. [78] revealed that Aβ25-35 treatment in neonatal rats significantly increased NO production in the hippocampus of the brain, and these mechanisms led to neurodegeneration and impaired spatial memory ability. Our results showed that the injection of Aβ25-35 significantly increased the NO concentration in the brain, liver, and kidney compared to that in normal mice. However, oral administration of PT showed the result of the suppressed NO production induced by Aβ25-35. To date, although there is no study about effect of PT on NO production in the AD mouse model, it was demonstrated that treatment with Populus species such as P. maximowiczii and P. davidiana inhibited NO production by more than 70% in LPS-induced RAW 264.7 murine macrophages [79]. The present study supports the hypothesis that PT inhibits Aβ25-35-mediated NO production and oxidative stress.
Aggregated Aβ is known to produce free radicals and ROS, which lead to the oxidation of DNA, proteins, and cell membranes [80]. These oxidative stresses consequently trigger neuronal cell death and play an important role in the development of AD [81]. According to Díaz et al. [82], the hippocampus of Aβ25–35-treated rats showed a significant increase in ROS production compared to that in normal rats. As shown in our data, in the brain, liver, and kidney, ROS production in Aβ25-35-induced mice was significantly elevated compared to that in normal mice, indicating that Aβ25-35 increased ROS production, which induced oxidative stress. However, PT treatment in mice significantly reduced ROS production. These results indicate that treatment with PT could protect against oxidative damage by inhibiting ROS production.
Oxidative stress is induced by an imbalance between antioxidants and oxidants. Several studies have reported that various antioxidant enzymes (glutathione peroxidase, superoxide dismutase, and catalase) are defense mechanisms that reduce and protect against oxidative stress [83]. Antioxidant enzymes break down and eliminate oxidants such as free radicals and reactive species. The activity of antioxidant enzymes has been reported to be decreased in patients with AD [84]. Therefore, reduced levels of lipid peroxidation, NO concentration, and ROS production by PT administration in the present study may be related to increased antioxidant enzymes. In addition, Lee et al. [63] demonstrated that antioxidant enzymes were significantly increased in rats treated with PT. Further studies on the roles of PT in antioxidative defense mechanisms in AD models, including antioxidant enzymes and balances between oxidative stress, are needed to clarify the effect of PT. Furthermore, neurotoxicity induced by Aβ accumulation in the brain induces oxidative stress and neuronal loss, thereby resulting in an inflammatory response [85]. Astrocytes and microglia are activated in the AD brain and release cytotoxic molecules, including pro-inflammatory cytokines (e.g., TNFα), upon exposure to Aβ. Consistently, the interaction between oxidative stress and inflammation provokes AD pathology. Our study group previously determined that PT-treated SH-SY5Y neuronal cells significantly downregulated the expression of inflammatory proteins iNOS and COX-2 in Aβ25–35 stimulation, which indicated that PT has a protective effect on Aβ25–35-mediated inflammation (data not shown). In the present study, we confirmed the antioxidant activity of PT in AD mice. Based on the interaction between oxidative stress and inflammation, PT may also exert anti-inflammatory activity. PT may also exert potential anti-inflammatory activity, although further studies are needed to understand the inflammation-related mechanisms of PT in Aβ25-35-injected mice.
In conclusion, the AD medications used in the clinic are aimed at delaying the progression of AD or alleviating symptoms rather than the fundamental treatment of AD. Moreover, it is necessary to develop a material derived from natural products with few side effects, because problems such as resistance and toxicity have been reported when AD medication is administered in the long term [11]. The present study demonstrated that administration of the EtOAc fraction of PT has a protective effect on short- and long-term memory and cognitive ability damaged by Aβ25-35 injection. Moreover, the EtOAc fraction of PT played an inhibitory role in Aβ25-35-induced oxidative stress by regulating lipid peroxidation, NO, and ROS. Therefore, the administration of PT is believed to protect against cognitive impairment by a mechanism that suppresses oxidative stress in an Aβ25-35-induced AD mouse model. Taken together, our study suggests that PT and active compounds from PT can be used as natural materials for AD improvement, thus suggesting the therapeutic potential of this agent for AD treatment, although further studies and clinical trials of this for AD are needed.
Go to :
 XML Download
XML Download