

 PDF
PDF Citation
Citation Print
Print


Introduction
Cholangiocarcinoma is a type of cancer originating from biliary epithelial cells and is classified as intrahepatic cholangiocarcinoma (ICC), extrahepatic cholangiocarcinoma, and hilar cholangiocarcinoma according to its anatomical position [1,2]. Although cholangiocarcinoma is rare cancer, its morbidity and mortality have increased rapidly worldwide in recent decades. ICC accounts for approximately 10% of all cholangiocarcinomas and its malignancy degree is second only to hepatocellular carcinoma in primary liver cancer. Currently, the only choice for ICC treatment is surgery. However, only 15% of patients with ICC undergo surgery. Additionally, the overall survival time for patients with ICC after diagnosis is only 6–24 months, and the morbidity and mortality of ICC are nearly the same, after surgery, the median survival time of patients with ICC ranges from 27 to 36 months [2–4]. Few studies have evaluated the mechanisms of the poor prognosis of ICC or described reliable biomarkers for ICC. Thus, further investigations are urgently needed.
Many studies have shown that most (70%) of the genome is transcribed into long non-coding RNA (lncRNA) [5,6]. LncRNA is a long RNA (> 200 nucleotides) lacking a functional open reading frame and does not encode proteins. LncRNAs are classified into five categories based on their locations in the transcript: sense, antisense, bidirectional, intronic, and intergenic. Many lncRNAs have been studied because of their impact on diseases, particularly cancer. For example, lncRNA-HOX transcript antisense RNA promotes prostate cancer [7], lncRNA-retinoic acid receptor-related orphan receptor (ROR) predicts poor prognosis in pancreatic cancer [8]. Although lncRNA plays an important role in gene regulation, studies of lncRNA are limited. Cancer-associated susceptibility 15 gene (CASC15) is a newly discovered lncRNA which located at 6p22.3. Our previous study reported that CASC15 plays an important role as an oncogene in hepatocellular carcinoma [9]. Recent studies have reported that CASC15 may have an influence on ICC by promoting an inflammatory microenvironment [10,11]. However, the other mechanism of CASC15 in ICC remains unclear.
Peroxiredoxin 2 (PRDX2) is a member of the peroxiredoxin family, which is a large family of antioxidant enzymes that act as antioxidants in cells [12]. PRDX2 has been reported to function in cancer cells by regulating H2O2-dependent signaling [13] and reactive oxygen species-related signaling pathways [14]. Recent studies showed that PRDX2 is highly expressed in many cancers and plays a protective role in lung cancer [15], breast cancer [16], and esophagus cancer [17]. Thus, PRDX2 is a key protein in cancer cell proliferation, invasion, and apoptosis. A previous study showed that PRDX2 might function in cancer by inducing phosphoinositide 3-kinase (PI3K)/AKT pathway [18]. However, the function and functional mechanism of PRDX2 in ICC remains unclear. Thus, we focused on the relationship between lncRNA-CASC15 and PRDX2 to explore the mechanism of these two important molecules in ICC.
Materials and Methods
1. Tissues and cell lines
ICC tissue and para-carcinoma tissue were provided by the Key Laboratory of Precision Diagnosis and Treatment for Hepatobiliary and Pancreatic Tumor of Zhejiang Province. These tissues were collected from patients with hepatectomy in 2013–2016 at Medicine School of Zhejiang University, and pathological examination confirmed the diagnosis of ICC. Clinical data were all collected for clinicopathologic and survival analysis. Tissues were stored at −80°C. ICC cell lines (CCLP, RBE, 9810, and HuCCT1) were provided by the Key Laboratory of Precision Diagnosis and Treatment for Hepatobiliary and Pancreatic Tumor of Zhejiang Province. Cells were grown in Roswell Park Memorial Institute 1640 (9810, RBE, and CCLP) and Dulbecco’s modified Eagle’s medium (HuCCT1, Biological Industries, Cromwell, CT) containing 10% fetal bovine serum (FBS), purchased from Thermo Fisher Scientific (Waltham, MA). Cells were cultured at 37°C in a 5% CO2 incubator.
2. RNA extraction and quantitative polymerase chain reaction
We used TRIzol reagent (Thermo Fisher Scientific) for the extraction of total RNA from the tissues and cells. NanoDrop ND-1000 UV/visible photometer (Thermo Fisher Scientific) was used to assess RNA purity. We used a real-time PCR instrument (Bio-Rad, Hercules, CA) to evaluate the relative expression of CASC15. Primers for glyceraldehyde-3-phosphate dehydrogenase (GAPDH; F: 5′-CGGAGTCAACGGA-TTTGGTCGTAT-3′, R: 5′-AGCCTTCTCCATGGTGGTGAAGAC-3′) and CASC15 (F: 5′-CACACGCATGGAAAACCC-AG-3′, R: 5′-GAGGACCTGAGCTGTAAGCC-3′) were designed by and purchased from Shanghai Biological Engineering Technology Co. (Shanghai, China). Primers for PRDX2 (H-prdx2-S: 5′-TAATGATTTGCCTGTGGGACG-3′; H-prdx2-A: 5′-CGTTGGGCTTAATCGTGTCA-3′) were designed by Wuhan Servicebio Co. (Wuhan, China).
3. Western blotting
Proteins were extracted by radioimmunoprecipitation assay (Biyuntian Biotechnology Co., Shanghai, China) with 1% phenylmethylsulfonyl fluoride and 1% phosphate inhibitor. Proteins were fractionated in 3-morpholinopropane-1-sulfonic acid/sodium dodecyl sulfate running buffer (Thermo Fisher Scientific) with a protein electrophoresis meter (Bio-Rad) and transferred to poly(vinylidene) fluoride membranes (Bio-Rad) with a Trans-Blot (Bio-Rad). The antibodies used were GAPDH (ab9485, Abcam, Cambridge, UK), caspase-8 (ab32397, Abcam), poly(adenosine diphosphate-ribose) polymerase (PARP) (ab32064, Abcam), BAX (ab32503, Abcam), cyclin E1 (ab33911, Abcam), cyclin D1 (ab134175, Abcam), cyclin B1 (ab32053, Abcam), CDK4 (ab108357, Abcam), CDK6 (ab124821, Abcam), p21 (ab109520, Abcam), N-cadherin (ab18203, Abcam), E-cadherin (ab15148, Abcam), PI3 kinase p110α (4255S, Cell Signaling Technology, Danvers, MA), AKT (9272S, CST), phospho-AKT (Ser473) (4060S, Cell Signaling Technology), c-Myc (9402S, Cell Signaling Technology), phospho-c-Myc (Ser62) (13748S, Cell Signaling Technology), and PRDX2 (10545-2-AP, Proteintech, Rocky Hill, NJ). We used GAPDH as an internal reference.
4. Cell interference, transfection, and infection
Cells were cultured in 6-well plates with 5 μL siRNA and 5 μL Lipofectamine 2000 (Thermo Fisher Scientific) for 6 hours. The short interfering RNA (siRNA) named as RiboTM h-CASC15 Smart Silencer was provided by Guangzhou Ruibo Biological Co. (Guangzhou, China) and had the following sequences: 5′-CCCTCAGGTGACTACAGAT-3′, 5′-GCTCAA-CCACATCTAATTT-3′, 5′-GCAACATGCTTCACTGTCT-3′, 5′-GATCGCTGGGAATTCTCCAC-3′, 5′-TCAGAGCTGGCTG-CCTGACA-3′, 5′-GCCAAGAAGAGTATGCAGAG-3′. SiRNAs named as PRDX2(h)-si-1,2,3 were provided by Guannan Biological Co. (Hangzhou, China; PRDX2(h)-si-1: AGA-UCAUCGCGUUCAGCAA tt, UUGCUGAACGCGAUGA-UCU tt; PRDX2(h)-si-2: ACCCUCUGGACUUCACUUU tt, AAAGUGAAGUCCAGAGGGU tt; PRDX2(h)-si-3: GACAGCAAGGAAUAUUUCU tt, AGAAAUAUUCCUUGCUG-UC tt).
Cells were cultured in 6-well plates with 1 μg plasmid PRDX2 and 3 μL Polyjet reagent (SignaGen, Shandong, China) for 24 hours. The plasmid PRDX2 was provided by Vigen Biosciences (Shandong, China).
The cells were cultured in 6-well plates with lentivirus (multiplicity of infection=10:1) (Jikai Gene, Shanghai, China) for 24 hours. The sequence of lentivirus-CASC15 was 5′-GAGCAGATAGCTGAAGAGAGA-3′.
5. Flow cytometry
Digital BD LSR II flow cytometry (BD Biosciences, Franklin Lakes, NJ) was used to evaluate apoptosis and cell cycle progression. Collected cells were mixed with binding buffer and 5 μL fluorescein isothiocyanate-labeled annexin V and 5 μL propidium iodide (BD Biosciences) to examine apoptosis. To evaluate cell cycle progression, we preserved the cells in 75% alcohol for 24 hours and mixed with cell cycle liquid (BD Biosciences).
6. Transwell migration and invasion assays
A Transwell assay (Millipore, Billerica, MA) was performed to evaluate migration and invasion. The upper chamber was filled with 4×104 cells in 200 μL FBS-free medium, and the lower chamber was filled with 700 μL 10% FBS medium. In the invasion assay, 20 μL (1:8, Matrigel:medium) was added to the upper chamber for 30 minutes at 37°C before the experiment. Wright-Giemsa dye (Jiancheng Technology Co., Nanjing, China) was used to stain the cells.
7. 5-Ethynyl-2′-deoxyuridine staining
Cells were cultured in a confocal culture dish for 48 hours, and a 5-ethynyl-2′-deoxyuridine (EdU) DNA Proliferation In Vitro Detection kit (C10310, Ruibo Biological Technology Co.) was used to test cell proliferation. Fluorescence microscopy (Olympus, Tokyo, Japan) was used to observe fluorescence.
8. Cell Counting Kit-8 assay and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
A Cell Counting Kit-8 (CCK-8) Kit (Dojindo, Kumamoto, Japan) was used to detect cell viability. Cells were cultured in 96-well plates (3,000 cells per well). After 0, 24, 48, 72, and 96 hours, we added 100 μL CCK-8 solution and incubated the cells for 3 hours at 37°C. A DYNEX MUM microplate reader (DYNEX Technologies, Chantilly, VA) was used to detect the absorbance at 450 nm. An MTT kit (C11019-2, Ruibo Biological Technology Co.) was used to measure active cells. After 48 hours of incubation of 0, 2, 3, 5, 10, 15, 25, 40, 60, and 100 ng/mL 5-fluorouracil (5-FU), 100 μL MTT solution was incubated with the cells for 4 hours at 37°C. Next, we added 100 μL formazan solving liquid; after 4 hours incubation at 37°C, we detected the absorbance at 570 nm.
9. Fluorescence in situ hybridization and immunofluorescence
Fluorescence in situ hybridization (FISH) and immunofluorescence experiments were carried out to examine the intracellular localization of CASC15 and PRDX2. ICC cells were grown on cover glass and then fixed with 4% paraformaldehyde (with DEPC). Next, probe for CASC15 and antibody for PRDX2 were prepared, with CASC15 marked as red and PRDX2 marked as green. CASC15 probe is 5′-CY3-GCGTGCTTCGGGCAGGGCTACA-3′ and PRDX2 antibody used was the same as western blotting. After hybridization, photos were acquired under a fluorescence microscope.
10. RNA pulldown and RNA immunoprecipitation
RNA pulldown was used to identify the proteins that bind to RNA. We designed RNA probes CASC15-sense-F: TAA-TACGACTCACTATAGGGCAAAAGGGGGAACTCTGA-TG; CASC15-sense-R: CTGTCATCTCTCCCCCACAT; CAS-C15-antisense-F: CAAAAGGGGGAACTCTGATG; and CAS-C15-antisense-R: TAATACGACTCACTATAGGGCTGTCAT-CTCTCCCCCACAT) as complementary strands of sense and antisense CASC15 which were used to mark CASC15. After incubation in cell lysis buffer, RNA-protein complexes were obtained. The complex was separated and resolved with sodium dodecyl sulfate polyacrylamide gel electrophoresis and visualized by silver staining, showing bands at 25–170 kDa. silver staining was performed for detecting whether there were differentially expressed proteins between sense-CASC15 and antisense-CASC15 group. Mass spectrometry was performed to analyze the binding proteins.
RNA immunoprecipitation (RIP) assay was performed to detect the sequence of RNA in the RNA-protein complex. The RIP kit was supplied by Merck (Billerica, MA). We used an antibody for PRDX2 and IgG to detect the specific protein-RNA complex. After splitting the protein, quantitative polymerase chain reaction was conducted to evaluate the RNA sequence.
11. Tumor formation and immunohistochemistry
All studies including animal experiments were approved by the Zhejiang Medical Experimental Animal Care Commission. Twenty male nude mice were provided by the Key Laboratory of Precision Diagnosis and Treatment for Hepatobiliary and Pancreatic Tumor of Zhejiang Province. All nude mice received humane care and our experiments complied with the institution’s guidelines. Transfected cells were collected for subcutaneous injection. We injected 5 million cells into each mouse subcutaneously with 100 μL phosphate-buffered saline into the axillary region. After 1 month, tumor formation was observed. An immunohistochemistry (IHC) assay was performed to stain the monoclonal antibodies for PRDX2, PI3K, AKT, PAKT, and Ki67, the antibodies used were the same as western blotting.
12. Bioinformatic analysis and Statistical analysis
Gene expression Profiling Interactive Analysis (GEPIA) was used for bioinformatic analysis of PRDX2 in ICC, the data source is The Cancer Genome Atlas database (TCGA).
SPSS Statistics ver. 17 software (SPSS Inc., Chicago, IL) was used to perform statistical analysis. Data are presented as the mean±standard deviation. The t test was used to analyze significant differences, and p < 0.05 was considered to indicate a significant difference. The experiments were repeated three times, and error bars represent standard deviation.
Results
1. LncRNA-CASC15 highly expressed in ICC tissues and decreased the overall survival rate
ICC patients from 2013 to 2016 were selected for our study. Ninety-five patients underwent surgery. We collected their tumor tissue and adjacent tissue specimens, extracted the RNA, and detected the relative expression of CASC15. CASC15 was highly expressed in ICC tissues (Fig. 1A). We followed up 81 patients (14 patients were lost to follow-up); 50 patient tumor tissues highly expressed CASC15, while the other 31 patients showed low expression of CASC15. High expression of CASC15 indicated a poor prognosis, and the results were significant (Fig. 1B).
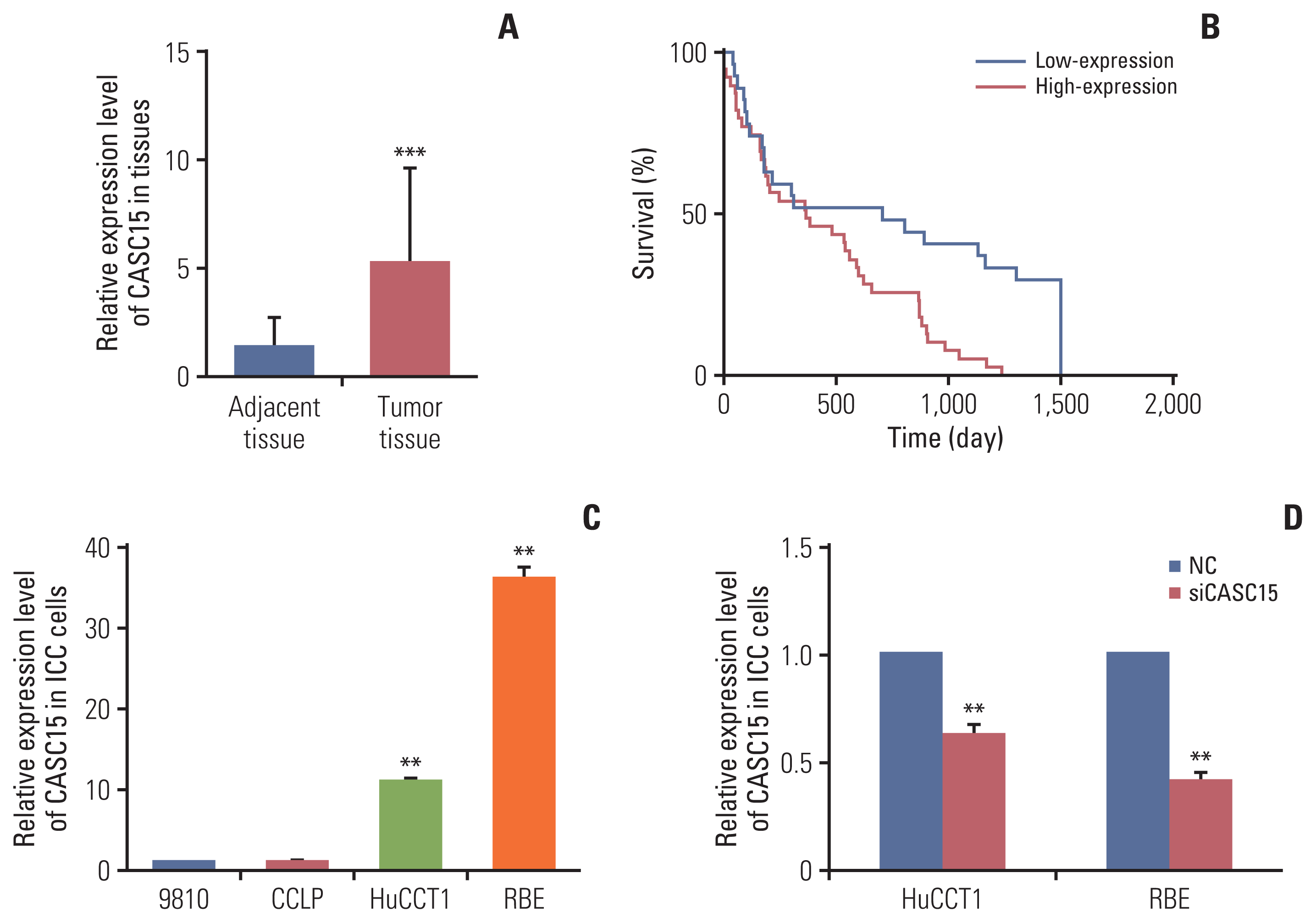
Fig. 1
Cancer-associated susceptibility 15 (CASC15) affected the prognosis of patients with intrahepatic cholangiocarcinoma (ICC). (A) Relative expression of long non-coding RNA (lncRNA)-CASC15 in tissues of 95 patients. (B) Survival curve analysis of the relationship between lncRNA-CANSC15 expression differences and survival time of patients. (C) Relative expression of ICC cell lines (CCLP, 9810, HuCCT1, and RBE). (D) The efficiency of knockdown of CASC15 in HuCCT1 and RBE cells. **p<0.01, ***p<0.001.
![]()
We analyzed the relationship between clinical data and relative CASC15 expression in the 81 patients. We found that CASC15 expression was related to the size of the tumor, Child-Pugh grade, and TNM stage. TNM staging is based on the American Joint Committee on Cancer (AJCC) cancer staging manual (8th edition). However, the difference in the expression of lncRNA-CASC15 was not significantly related to sex, age, α-fetoprotein, carbohydrate antigen 19-9, alanine aminotransferase, hepatitis B surface antigen, liver cirrhosis, vascular invasion, lymphatic metastasis, or distant metastasis (Table 1).
Table 1
Correlation between expression of CASC15 gene and clinical data of ICC
![]()
LncRNA-CASC15 was highly expressed in ICC tissues, indicating a larger tumor size, higher risk of surgery, higher tumor malignant degree, and poorer prognosis.
2. Decreased expression of lncRNA-CASC15 influences the function of ICC cells
We detected the relative expression of CASC15 in four types of ICC cells (CCLP, 9810, HuCCT1, and RBE). Next, we selected HuCCT1 and RBE for further experiments (Fig. 1C). We used siRNA-CASC15 to knock down the expression of CASC15 in HuCCT1 and RBE. The knockdown efficiency is good (Fig. 1D).
In the Transwell assay, the migration and invasion of HuCCT1 and RBE cells considerably decreased when CASC15 was knocked down (Fig. 2A and B, S1 Fig.). The EdU experiment showed that the cells were actively proliferating. In HuCCT1 and RBE cells, the EdU-stained cell proportion was significantly decreased in the knockdown group compared to in the control group (Fig. 2C–E). The CCK-8 test showed that cell viability was decreased in the knockdown group at 24, 48, 72, and 96 hours (Fig. 2F and G). Thus, CASC15 facilitated the migration, invasion, and proliferation of ICC cells.
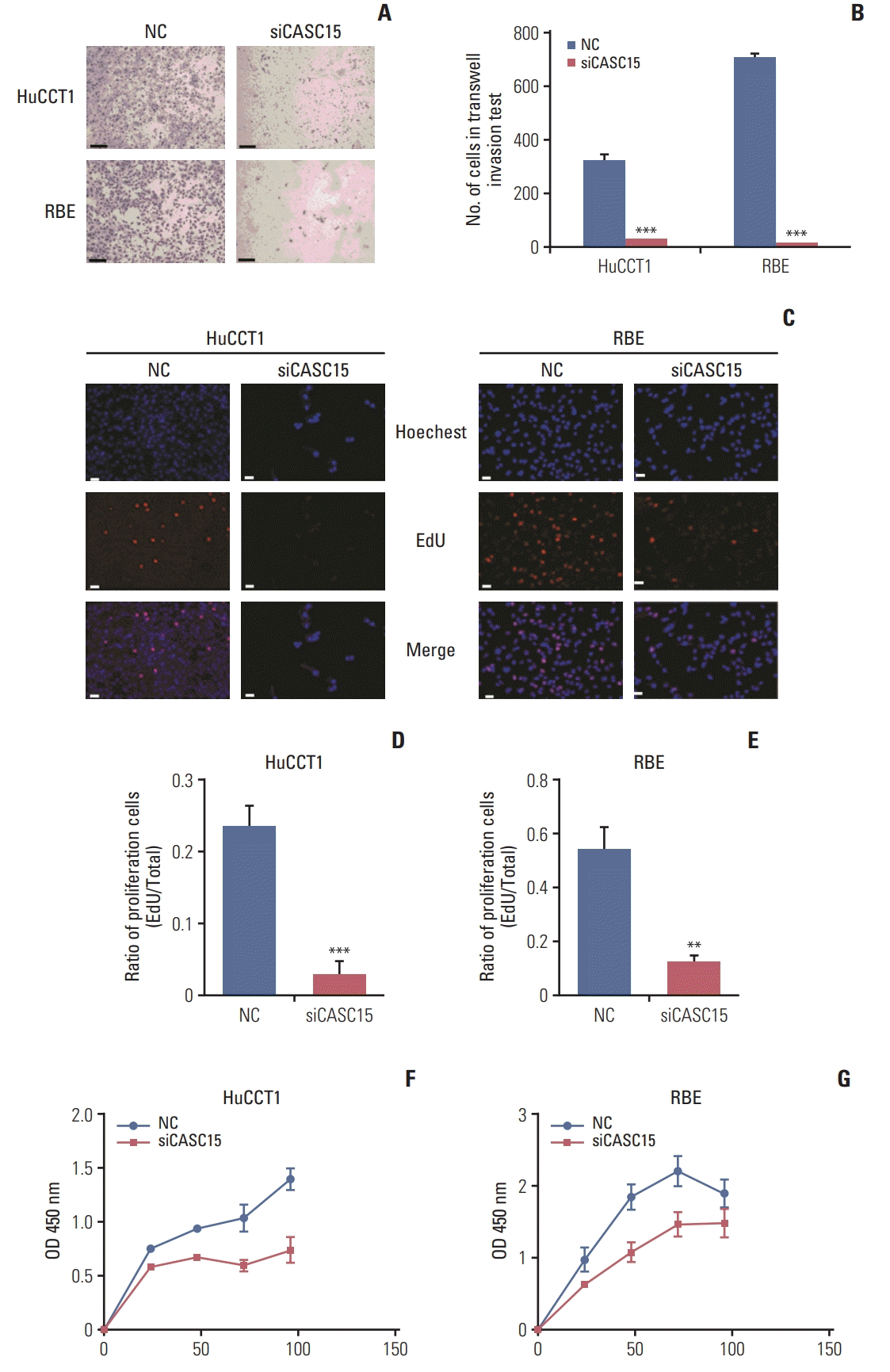
Fig. 2
The knockdown of cancer-associated susceptibility 15 (CASC15) expression inhibited cell invasion and proliferation. (A, B) Transwell invasion experiment and quantitative results. (C–E) EdU experiment showed that cell proliferation capacity was decreased in the siCASC15 group. Scale bars=100 μm. (F, G) Cell Counting Kit-8 (CCK-8) assay showed that the cell proliferation capacity decreased in the siCASC15 group. NC, negative control. **p<0.01, ***p<0.001.
![]()
We tested the apoptosis and cell cycle progression of HuCCT1 and RBE cells by flow cytometry. Apoptosis in the knockdown group was significantly increased when compared to the control group. (Fig. 3A and B). Western blotting showed that the expression of proteins relating to apoptosis such as BAX, cleaved PARP, and cleaved caspase-8 were increased in the knockdown group (Fig. 3E, left panel). Knocking down the expression of CASC15 promoted the apoptosis of ICC cells. In the knockdown group, the proportion of cells in G1 phase increased, whereas the proportion of cells in S phase decreased. After knocking down the expression of CASC15, G1/S transition was blocked (Fig. 3C and D). Western blotting showed that the expression of cyclin E1 and cyclin D1, and CDK4, and CDK6 was reduced in the knockdown group (Fig. 3E, right panel). These proteins affect the G1/S transition.
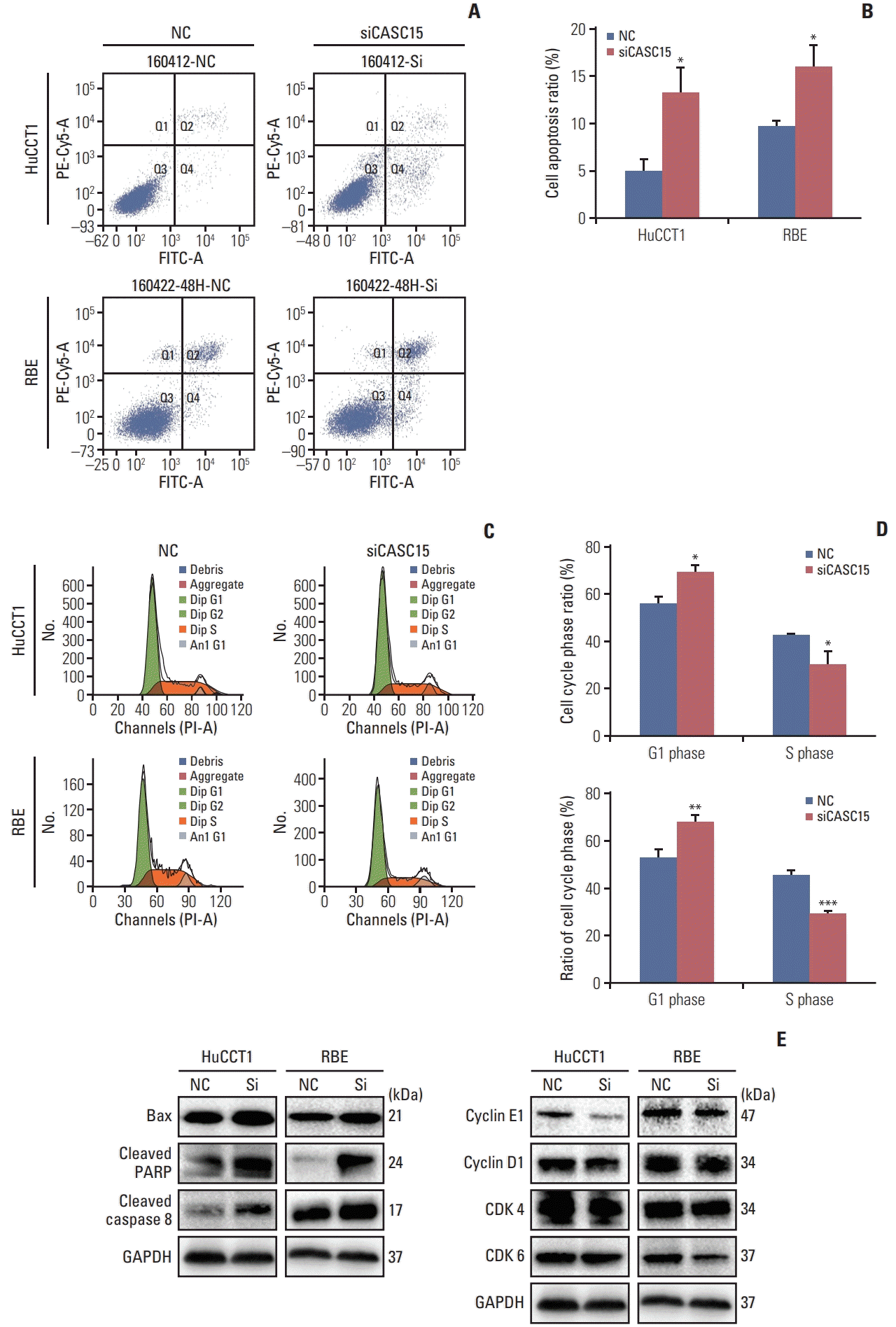
Fig. 3
The knockdown of cancer-associated susceptibility 15 (CASC15) expression promoted cell apoptosis and inhibited G1/S phase transformation. (A, B) Flow cytometry diagram of cell apoptosis and quantitative results. (C, D) Flow cytometry diagram of cell cycle and quantitative results. (E) Western blotting revealed that the expression of apoptosis and cell cycle-related proteins were changed in the siCASC15 group. GAPDH, glyceraldehyde 3-phosphate dehydrogenase; NC, negative control; PARP, poly(ADP-ribose) polymerase. *p<0.05, **p<0.01, ***p<0.001.
![]()
3. LncRNA-CASC15 is bound to protein PRDX2
We performed FISH to detect the location of CASC15 expression in ICC cells. CASC15 was expressed in both the cytoplasm and nucleus (Fig. 4A). RNA pulldown assay and mass spectrometry analysis showed that several proteins bind to CASC15, such as PRDX2, SFN, and ANXA2 (Fig. 4B). The RIP assay further verified that PRDX2 and SFN bound to CASC15 (Fig. 4C).
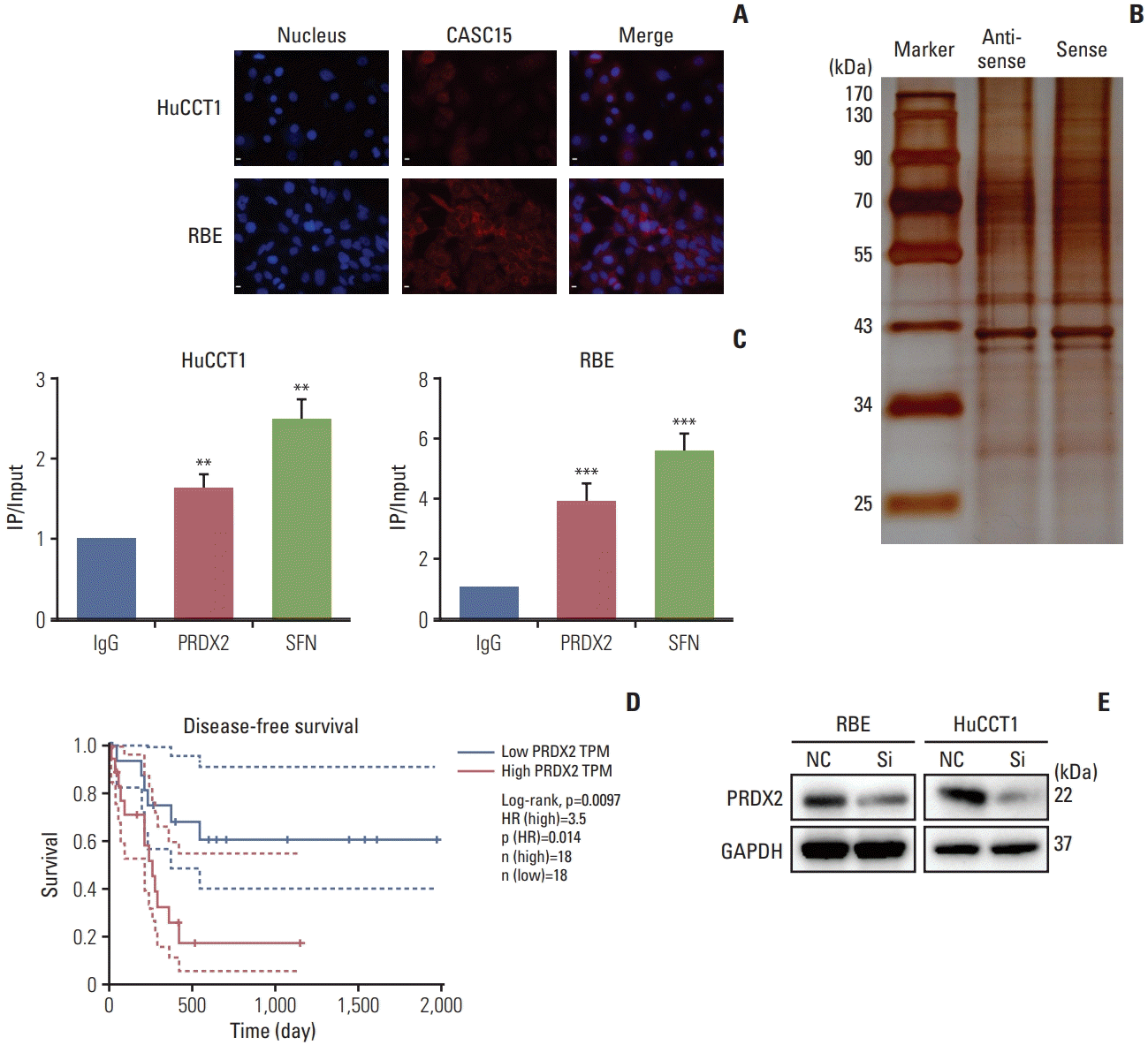
Fig. 4
Cancer-associated susceptibility 15 (CASC15) was associated with peroxiredoxin 2 (PRDX2) and regulated its expression. (A) Location of CASC15 in intrahepatic cholangiocarcinoma (ICC) cells. (B) RNA-protein complexes were resolved with sodium dodecyl sulfate polyacrylamide gel electrophoresis and visualized by silver staining, showing bands at 25–170 kDa. (C) RNA immunoprecipitation (IP)–quantitative polymerase chain reaction showed PRDX2 bound to CASC15. (D) Relationship of PRDX2 and disease-free survival of patients with ICC in The Cancer Genome Atlas database database. (E) Western blotting revealed that PRDX2 expression was decreased in the siCASC15 group. GAPDH, glyceraldehyde 3-phosphate dehydrogenase; NC, negative control. Scale bars=200 μm. **p<0.01, ***p<0.001.
![]()
TCGA revealed that the expression of PRDX2 was associated with disease-free survival of ICC (Fig. 4D). Thus, we chose PRDX2 as the key target protein of CASC15 for further experiments. Western blot analysis showed that the expression of PRDX2 decreased when the expression of CASC15 was knocked down (Fig. 4E). Additionally, FISH showed that the expression locations of PRDX2 and CASC15 were nearly coincident (S2 Fig.). Thus, PRDX2 was related to CASC15 in ICC cells.
4. PRDX2 affects ICC through the PI3K/AKT/c-Myc pathway
We used siRNA-PRDX2-1,2,3 to knock down the expression of PRDX2 in RBE and HUCCT1 cells (Fig. 5A, S3A Fig.). We selected siRNA-1,3 for further experiments. After knocking down PRDX2 expression, the proportion of cells in G1 phase increased, but the proportion of cells in S phase decreased, and western blotting revealed that the expression of cyclin B1 and cyclin D1 was reduced in the knockdown group (Fig. 5B and D, S3B Fig.). The result agreed with the results for CASC15 in ICC cells. In the Transwell invasion experiment, knockdown of PRDX2 expression decreased the number of cells that passed through the Transwell (Fig. 5C, S4A Fig.). The western blot results showed that proteins in endothelial mesenchymal transition (EMT) pathway such as E-cadherin and N-cadherin were altered, as observed in the Transwell assay (Fig. 5D, S3B Fig.).
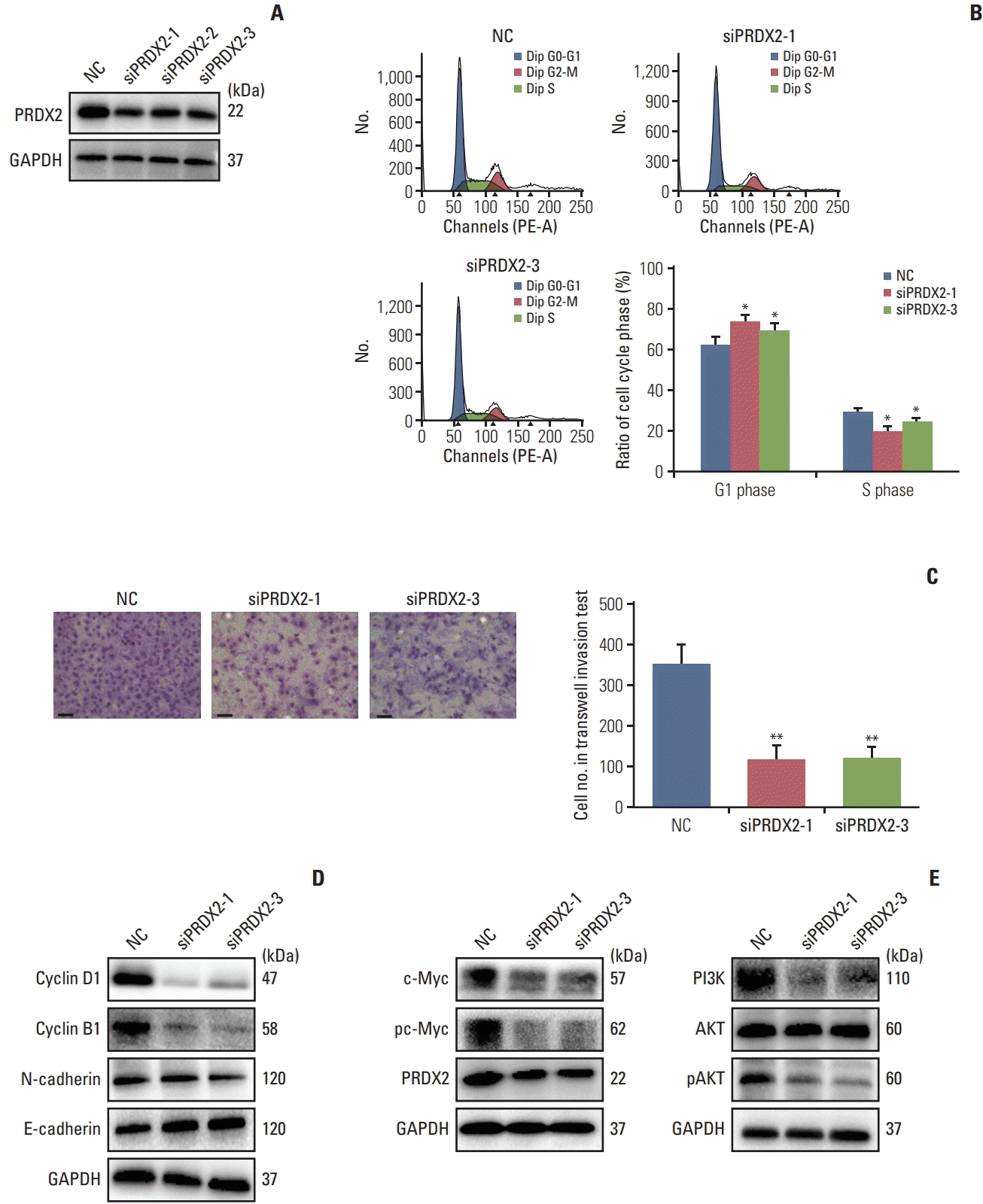
Fig. 5
The knockdown of peroxiredoxin 2 (PRDX2) expression inhibited the invasion and G1/S phase transformation of intrahepatic cholangiocarcinoma (ICC) cells. (A) Efficiency of PRDX2 knockdown in RBE cells. (B) Flow cytometry diagram of cell cycle and quantitative results. (C) Transwell invasion experiment and quantitative results of RBE cells. Scale bars=100 μm. (D) Western blotting showed that the expression levels of cell cycle and endothelial mesenchymal transition (EMT)–related proteins were altered in the siPRDX2 group of RBE cells. (E) Western blotting showed that the phosphoinositide 3-kinase (PI3K)/AKT/c-Myc pathway was inhibited in the siPRDX2 group of RBE cells. GAPDH, glyceraldehyde 3-phosphate dehydrogenase; NC, negative control. *p<0.05 and **p<0.01.
![]()
When PRDX2 expression was knocked down in ICC cells, the IC50 of 5-FU was greatly decreased (S4B Fig.). Compared to the control group, apoptosis in the knockdown group was significantly increased in the presence of 10 ng/mL 5-FU (Fig. 6E and F). Without 5-FU, apoptosis in the knockdown group changed very little (S4C Fig.).
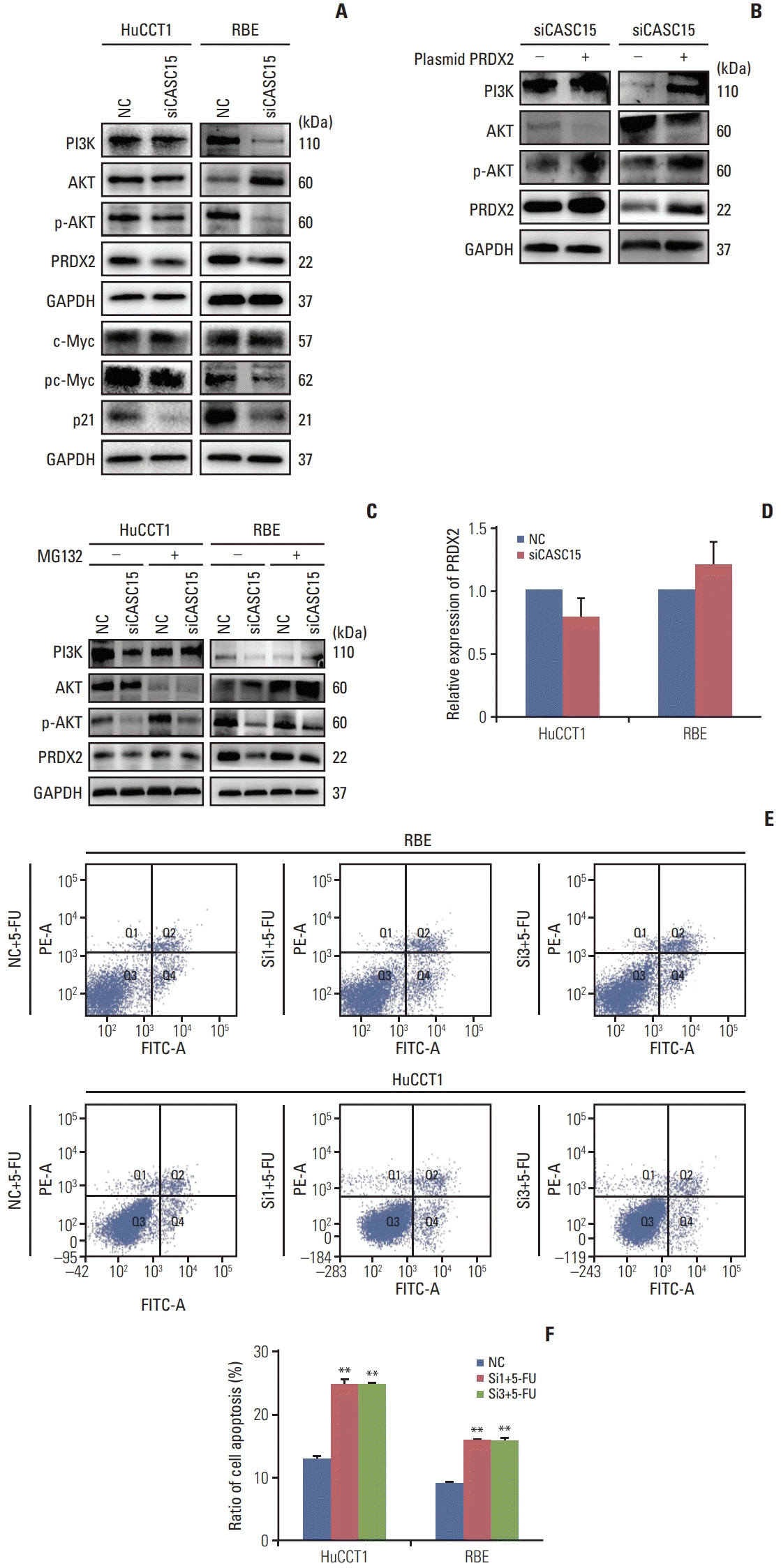
Fig. 6
Cancer-associated susceptibility 15 (CASC15) affected phosphoinositide 3-kinase (PI3K)/AKT/c-Myc pathway through peroxiredoxin 2 (PRDX2) and CASC15 regulated PRDX2 by inhibiting ubiquitination. (A) Western blotting showed that the PI3K/AKT/c-Myc pathway was inhibited in the siCASC15 group. (B) Western blotting showed that the PI3K/AKT/c-Myc pathway was enhanced when PRDX2 was overexpressed in the siCASC15 group. (C, D) Expression of PRDX2 was increased and the PI3K/AKT pathway was activated when proteasome formation was inhibited (left panel). Relative expression of PRDX2 changed little in the siCASC15 group (right panel). (E, F) Flow cytometry diagram of cell apoptosis and quantitative results. 5-FU, 5-fluorouracil; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; NC, negative control. **p<0.01.
![]()
After knocking down the expression of PRDX2 in RBE and HUCCT1 cells, western blot analysis showed that the PI3K/AKT/c-Myc pathway was blocked. Additionally, the protein p21, which regulates the cell cycle, and downstream proteins of the PI3K/AKT pathway were decreased in the knockdown group (Fig. 5E, S3C Fig.).
5. Overexpressed PRDX2 after knocking down the expression of CASC15 rescues the changes
Western blot analysis showed that PI3K/AKT/c-Myc pathway was blocked in the CASC15 knockdown group (Fig. 6A). Overexpressed PRDX2 after knocking down the expression of CASC15 rescued the suppression of the PI3K/AKT pathway (Fig. 6B). Additionally, overexpressed PRDX2 rescued the block of G1/S transition under knockdown of CASC15 (S5 Fig.). Thus, CASC15 functions in ICC possibly through PRDX2.
In the RNA pulldown assay and mass spectrometry analysis, we found that some ubiquitin-related proteins were bound to CASC15. Western blotting showed that the expression of PRDX2 was decreased in the CASC15 knockdown group (Fig. 4E). However, the relative expression of PRDX2 mRNA was only slightly changed (Fig. 6D). The decrease in PRDX2 and block of the PI3K/AKT pathway in the CASC15 knockdown group was rescued by adding MG132 (20 μg/mL for 2 hours) (Fig. 6C). Thus, CASC15 regulates the expression of PRDX2 by inhibiting proteasome formation.
6. Knockout of lncRNA-CASC15 expression suppresses ICC development in vivo
We selected HuCCT1 to carry out lentiviral transfection experiments. After 1 month of subcutaneous injection of cells after transfection, the tumor volume in the control group was 5.47±2.06 cm2, and the weight was 5.57±2.10 g. The tumor volume in the knockdown group was 0.37±0.51 cm2 (p < 0.001), and the weight was 0.38±0.51 g (p < 0.001). Knockdown of CASC15 decreased the growth of ICC in vivo (Fig. 7A–D).
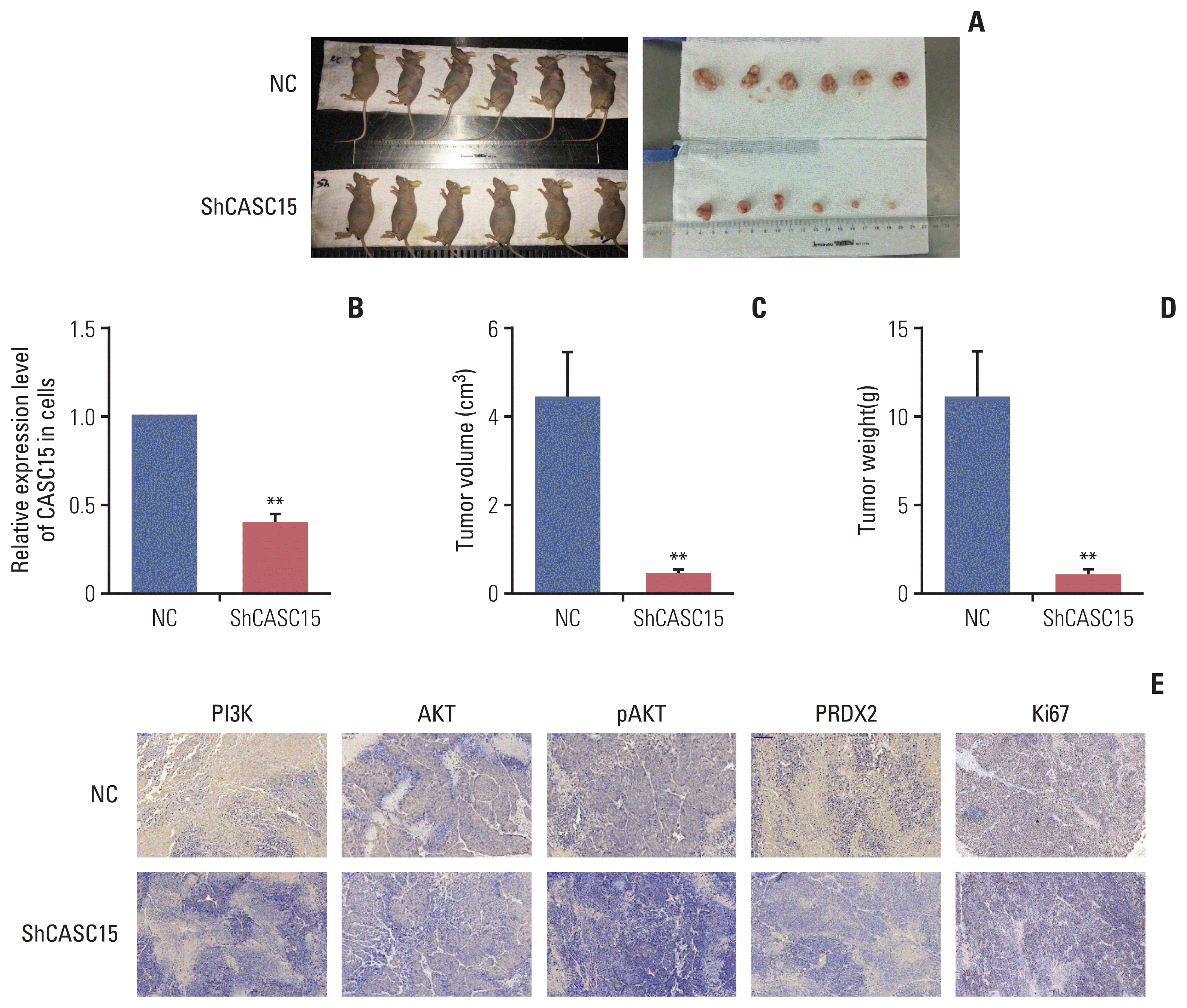
Fig. 7
The knockdown of cancer-associated susceptibility 15 (CASC15) in xenograft model inhibited the development of intrahepatic cholangiocarcinoma (ICC). (A–D) Knockdown of CASC15 in xenograft model decreased tumor size and weight. (E) Immunohistochemistry assay showed peroxiredoxin 2 (PRDX2), phosphoinositide 3-kinase (PI3K), AKT, pAKT, and Ki67 expression levels were decreased in the ShCASC15 group compared to in the control group. NC, negative control. Scale bar=100 μm. **p<0.01.
![]()
IHC showed that the expression of PRDX2 decreased in the knockdown group compared to in the control group, which matched the result in cell lines. IHC also showed that PI3K/AKT signaling pathway was inhibited in the knockdown group. CASC15 affected PRDX2 and PI3K/AKT signaling pathway in vivo as well (Fig. 7E).
Discussion
In our study, the expression of CASC15 in ICC tissues was higher than that in para-carcinoma tissues as observed for other tumor promoters, and CASC15 was highly expressed in ICC cell lines such as HuCCT1 and RBE cells. Previous studies showed that CASC15 promotes the progression and metastasis of cancers, indicating a poor prognosis. Based on our analysis of clinical data from patients with ICC, high expression of CASC15 indicated a poor prognosis, larger tumor size, and more advanced stage of TNM, suggesting that CASC15 facilitates the progression of ICC.
In cell functional experiments and animal experiments, we found that knockdown of CASC15 inhibited the proliferation, mitosis, tumor formation, migration, and invasion of ICC cells, while it increased the apoptosis of ICC cells. Knockdown of CASC15 decreased the expression of cyclin E1, cyclin D1, CDK4, and CDK6, inhibited the progression of cell cycle, the mitosis of ICC cells, and the proliferation of ICC cells. Many lncRNAs affect the cell cycle. For example, lncRNA-liver regeneration 1 was found to promote the expression of cyclin D1 by activating Wnt/β-catenin signaling [19]. Knockdown of CASC15 increased the expression of BAX, cleaved PARP, and cleaved caspase-8. Based on our results, lncRNA-CASC15 was involved not only in the death receptor-mediated pathway but also in the mitochondria-mediated pathway. EMT is the epithelial cell transition to mesenchymal cells or fibroblasts, giving cells migration and invasion abilities [20]. LncRNA-HIT [21] and lncRNA-ROR [8] are two lncRNAs involved in the EMT pathway. In the present study, knockdown of CASC15 decreased the expression of N-cadherin and migration and invasion of ICC cells. Thus, knockdown of CASC15 inhibited the migration and invasion of ICC cells.
We confirmed that protein-PRDX2 bound to CASC15 through RNA-pulldown and RIP experiments. Previous studies have shown that PRDX2 is associated with many cancers. Xu et al. [18] confirmed that PRDX2 affects the resistance to 5-FU in colon cancer. In our study, we showed that knockdown of PRDX2 inhibited the invasion of ICC cells, resulting in the block of G1/S transition and increasing the sensitivity to 5-FU. We further confirmed that the function of PRDX2 was linked to the PI3K/AKT/c-Myc signal pathway. A previous report showed that PRDX2 functions by activating PI3K/AKT signaling [18]. Our study clarified that the downstream of PI3K/AKT, c-Myc, was also activated by PRDX2. Furthermore, CASC15 is also an important activity factor of PI3K/AKT/c-Myc. Thus, CASC15 may regulate the function of ICC cells via PI3K/AKT/c-Myc. We showed that CASC15 activated PI3K/AKT through PRDX2 by rescue experiments, indicating that PRDX2 is an important downstream molecule. Some ubiquitination related proteins were found to bind CASC15 in RNA-pulldown experiments. CASC15 may regulate the expression of PRDX2 through the ubiquitination pathway. Although no studies have shown that CASC15 or PRDX2 is related to ubiquitylation, our study confirmed that CASC15 adjusted the expression of PRDX2 through ubiquitylation.
Our study showed that lncRNA-CASC15 was highly expressed in ICC and promoted the development of ICC. However, we did not perform overexpression experiments. We only considered CASC15 knockdown or knockout, and did not confirm that the function of CASC15 was enhanced by overexpression. Additionally, we only evaluated the basic mechanism of CASC15 and its relationship with PRDX2. We found that knockdown of CASC15 decreased the expression of PRDX2 and it was rescued by inhibiting proteasome formation, but we did not evaluate the mechanism or how it bound to the proteasome.
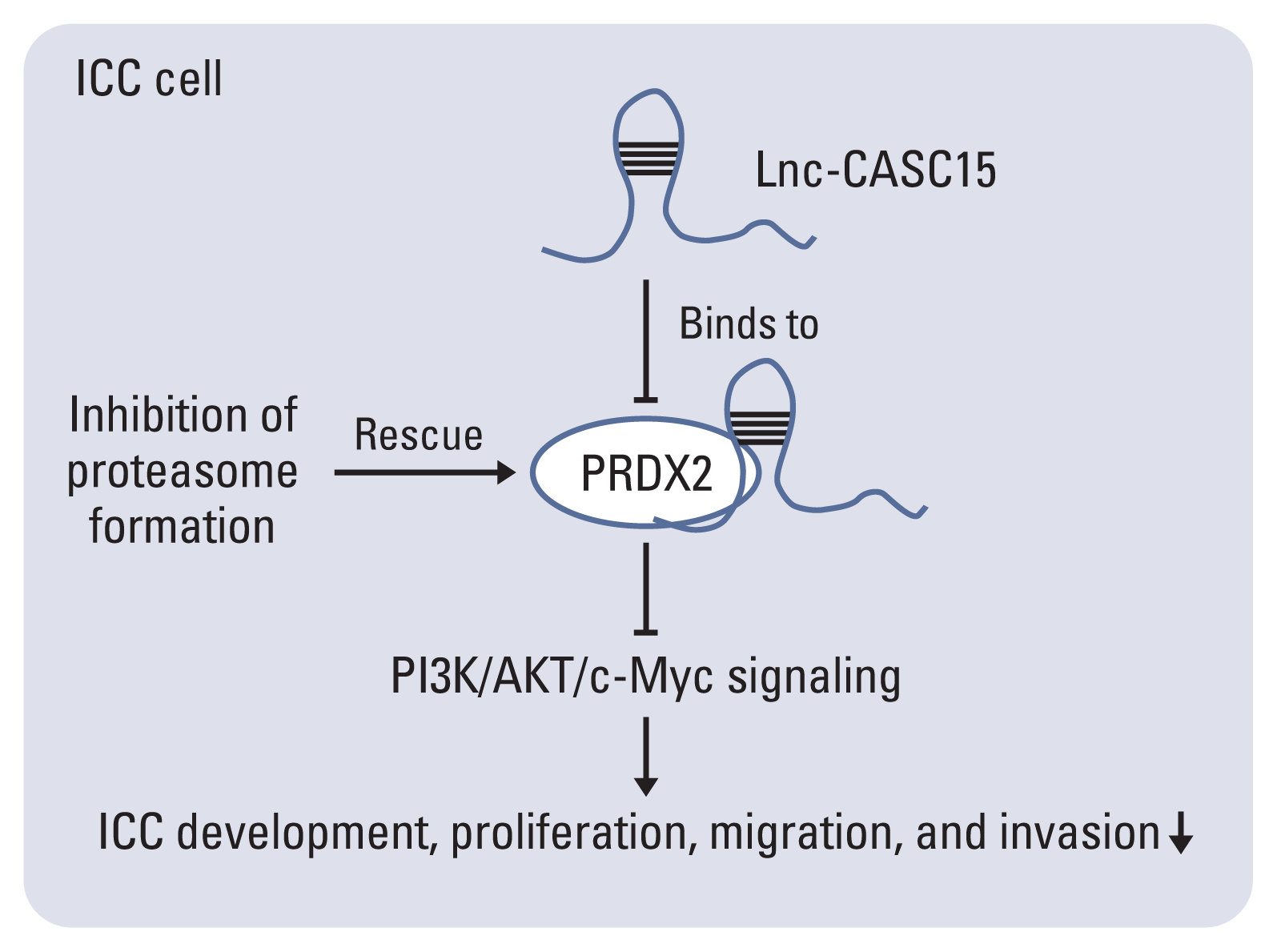
We thought that lncRNA-CASC15 is possibly an oncogenic lncRNA functioning in ICC (Fig. 8). CASC15 is likely to promote the migration, invasion, proliferation, and tumor formation of ICC cells and inhibits the apoptosis of ICC cells. High expression of CASC15 indicates a poor prognosis and high-grade malignancy. CASC15 activates the PI3K/AKT/c-Myc signal pathway possibly by binding to PRDX2 and inhibiting its degradation, resulting in ICC development. Given the role of CASC15 in ICC, lncRNA-CASC15 should be considered as a new promising therapeutic target in ICC therapy.
 XML Download
XML Download