

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The increasing prevalence of obesity has made it a serious public health problem worldwide [1]. An adipose tissue dysfunction is the primary defect in obesity and may link obesity to other diseases [1]. Previous studies have demonstrated the molecular mechanisms of obesity in adipose tissue [2]. Excessive adipose tissue stimulates the release of adipokines, including chemokines, cytokines, and hormones [34]. Increased cytokines secreted from adipose tissues activate the immune system and affect insulin signaling, which causes insulin resistance in the body [5]. These factors contribute to the association between obesity and other diseases, including type 2 diabetes, hypertension, and cardiovascular diseases, which are the most prevalent diseases in modern society. People who suffer from the prevalence of diseases caused by obesity have increased morbidity and mortality. Therefore, obesity and its comorbidities could destroy the quality of personal life and burden the social economy [678].
Aster yomena (Kitam.) Honda (AY) is a traditional herb for treating cough and asthma. AY has several therapeutic properties, including antioxidant, anti-inflammation, and anti-cancer [91011]. Han et al. [12] reported that AY inhibited adipogenesis in 3T3-L1 adipocytes by decreasing adipogenic transcription factors via the AMP-activated protein kinase (AMPK) pathway. On the other hand, the effects of ethyl acetate fraction from AY (EFAY) against obesity-related metabolic dysfunctions in vivo have not been reported. This study examined whether AY improves obesity and insulin resistance under a high-fat diet (HFD)-induced obesity mice model. Among the extract and fractions from AY, the ethyl acetate fraction from AY (EFAY) was chosen as an active fraction in the present study. EFAY exhibited the strongest activity in eliminating oxidative radicals among other extracts and fractions, such as ethanol, n-butanol, dichloromethane, and n-hexane [13]. Moreover, EFAY inhibited adipocyte differentiation in 3T3-L1 preadipocytes by regulating adipogenesis and lipolysis [14]. Based on these reports, this study investigated the effects of EFAY against insulin resistance in a HFD-induced obese mice model.
Go to : 
MATERIALS AND METHODS
Reagents and instruments
Thiobarbituric acid (TBA) was obtained from Acros Organics (Geel, Belgium). n-Butanol (n-BuOH) was supplied by Yakuri Pure Chemical Co. (Kyoto, Japan). Malondialdehyde (MDA), sulfanilamide, and trichloroacetic acid were purchased from Sigma Chemical Co. (Saint Louis, USA). Sodium chloride (NaCl) was obtained from the LPS solution (Seoul, Korea). Pyridine was acquired from Wako Pure Chemical Industries., Ltd. (Osaka, Japan). Radioimmunoprecipitation assay (RIPA) buffer was procured from Elpis Biotech. (Daejeon, Korea). Polyvinylidene fluoride (PVDF) membrane was obtained from Millipore Co. (Billerica, USA). The primary and secondary antibodies were manufactured by Cell Signaling Tech. (Beverly, USA), Abcam Co. (Cambridge, UK), Calbiochem Co. (San Diego, USA), and Bioss Inc. (Beijing, China).
Animals and diets
Male C57BL/6J mice aged 5 weeks were purchased from Jung-Ang Lab Animal Inc. (Seoul, Korea). The mice were housed in plastic cages and maintained under controlled conditions at 20 ± 2°C and 50 ± 10% relative humidity with a 12 h light-dark cycle. The Pusan National University Institutional Animal Care and Use Committee approved the procedures in this experiment (PNU-2017-1758). After a week of adaptation, the mice were divided into 4 groups defined as normal, control, EFAY100, and EFAY200 groups (6 mice per group). The normal group was fed a normal diet (ND) containing 10% fat (% of energy consumption as fat, D12450B), and the other 3 groups were fed a HFD containing 60% fat (% of energy consumption as fat, D12492) (Table 1) [1516]. The diets were manufactured by Research Diets (New Brunswick, NJ). All mice could access the diet and water freely. From the end of the 12th week of the experimental period, the mice were orally administered for 4 weeks as in the following: normal and control groups were given the vehicle (water); the EFAY100 group was given EFAY at a dose of 100 mg/kg body weight; the EFAY200 group was given EFAY in a dose of 200 mg/kg body weight. The vehicle of EFAY is water and the administration was conducted once daily. After 16 weeks of dietary supplementation, the mice were fasted for 16 h and sacrificed with CO2 gas under anaesthesia. Blood samples were collected via the heart and centrifuged to separate serum. The obtained serum and tissues were kept at −80°C for further experiments.
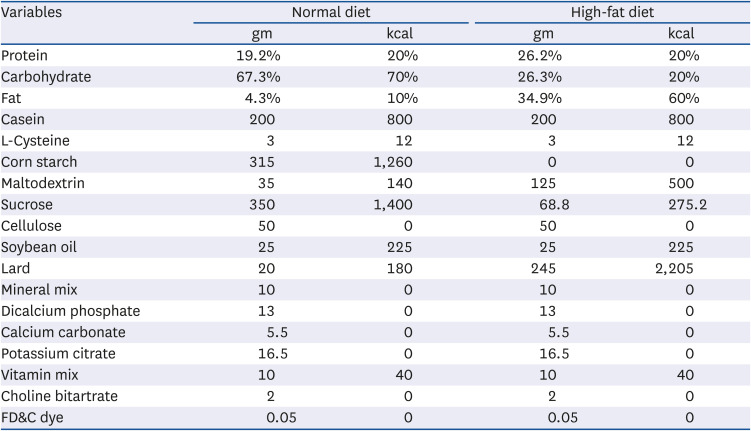
Table 1
Compositions of diets
![]()
Measurement of body weight, organ weight, and FER
The body weights of the mice were measured every week, and the food intake was measured every other day. Tissues including the brain, liver, kidney, and epididymal fat were weighed after being cleansed with 0.9% NaCl. The food efficacy ratio (FER) was calculated using the following equation [17]. FER (%) = [body weight gain (g)/ food intake (g)] × 100.
Measurement of serum lipid profiles
The serum levels of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were determined using serum glutamic oxaloacetic transaminase and serum glutamic pyruvic transaminase assay kits (AM103-K and AM102; Asan Pharm., Seoul, Korea). The triglyceride (TG), total cholesterol (TC), and high-density lipoprotein (HDL) cholesterol levels in the serum were measured by enzymatic assay kits (AM157S, AM-202, and AM-203; Asan Pharm., Seoul, Korea). The serum low-density lipoprotein (LDL) cholesterol levels were calculated using the following equation [18]. LDL cholesterol (mg/dL) = [(TC − HDL cholesterol) − (TG/5)].
Measurement of blood glucose, insulin, and homeostasis model assessment of insulin resistance (HOMA-IR) levels
The fasting blood glucose was monitored every week. Blood samples were collected from the tail vein of mice after 16 h fasting. The blood glucose level was measured using a blood glucose meter (AGM-3000; Allmedicus Co., Ltd., Gyeonggi-do, Korea). The serum insulin level was measured using an ELISA assay kit (EZRMI-13K; Millipore, Schwalbach, Germany). The index of HOMA-IR was calculated using the following equation [1920]. HOMA-IR = [Fasting blood glucose (mmol/L) × fasting insulin (μU/mL)]/22.5.
Measurement of lipid peroxidation in liver
The lipid peroxidation levels in tissues were analyzed by measuring the MDA level using the procedure described by Ohkawa et al. [21]. Briefly, approximately 0.1 g of the liver from each mouse and 0.9% NaCl solution were mixed (1:9 v/v) and homogenized. The mixed reagent containing 0.67% TBA and 15% TCA was added to the homogenate and boiled for 20 min. The mixtures were immediately transferred to ice for cooling, n-BuOH:pyridine mixture (15:1 v/v) was added. After centrifugation at 3,000 rpm for 10 min, the absorbance of the n-BuOH-pyridine layer was measured at 540 nm. The lipid peroxidation level was calculated using a standard curve derived from MDA and expressed as MDA equivalents.
Western blot analysis
For protein extraction, the tissues were homogenized with RIPA buffer supplemented with a protease inhibitor cocktail. The homogenates were separated by centrifugation, and the supernatants were used in the subsequent experiment. Equal amounts of protein from the supernatants were separated on 8–13% sodium dodecyl sulfate-polyacrylamide gels and then transferred to PVDF membranes. The membranes were blocked with 5% skim milk in phosphate-buffered saline containing tween 20 for 50 min at room temperature and incubated overnight at 4°C with the primary antibodies. The antibodies used in this study were as follows. The primary antibodies used were β-actin (1:1000; Cell Signaling Tech.; Beverly, USA), fatty acid synthase (FAS, 1:500; Cell Signaling Tech.), acetyl-CoA carboxylase (ACC, 1:500; Cell Signaling Tech.), phospho-ACC (p-ACC, 1:500; Cell Signaling Tech.), AMPK (1:500; Cell Signaling Tech.), phospho-AMPK (p-AMPK, 1:500; Cell Signaling Tech.). The membranes were rinsed and incubated with the appropriate secondary antibodies (1:500; Cell Signaling Tech.) for 1 h at room temperature. The immunoreactive proteins were visualized using a chemiluminescent imaging system (CoreBio, Seoul, Korea).
Statistical analysis
The data in the present study are expressed as mean ± SD. Statistical differences between the groups were analyzed using a one-way analysis of variance (ANOVA) and a Duncan’s post hoc test. P values < 0.05 were considered significant (n = 6).
Go to :
RESULTS
Effects of EFAY on the body weight gain, food intake, and organ weights
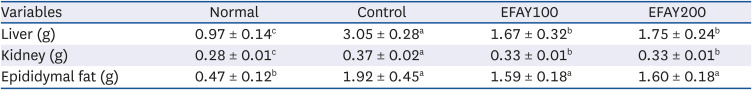
The initial body weight among the 4 groups showed no significant difference (Table 2 and Fig. 1). As expected, HFD-fed groups gained significantly higher body weight than the normal group at week 16. On the other hand, EFAY administration for 4 weeks suppressed body weight compared to the HFD-fed control mice at week 16. There was no significant difference in food intake among the groups, whereas the FER of mice fed a HFD was increased significantly compared to mice fed with ND. In contrast, EFAY administration decreased the FER significantly compared to HFD-fed control mice. The EFAY100 group showed a significantly lower body weight gain and FER than the control group. Hence, it was assumed that EFAY might affect body weight gain by decreasing FER. In the organ weights, the HFD-fed control group increased the weights of liver, kidney, and epididymal fat significantly compared to the ND-fed normal group (Table 3). On the other hand, the oral administration of EFAY to HFD-fed mice decreased the liver and kidney weights significantly. Although there was no significant difference, the epididymal fat weight showed decreased patterns in the groups with EFAY administration. EFAY administration regulated the weight gains of the body and organ weights in HFD-fed mice.
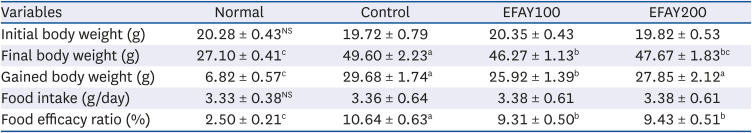
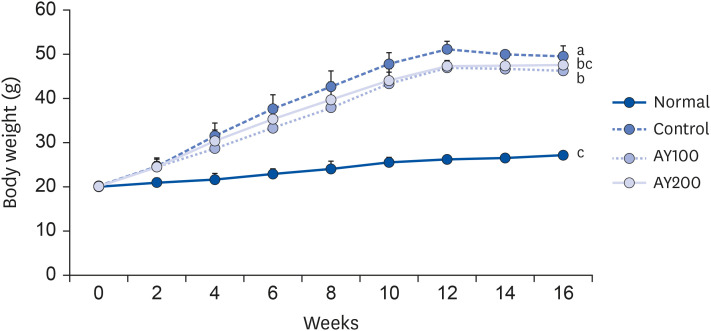 | Fig. 1Effects of the ethyl acetate fraction from Aster yomena (Kitam.) Honda on the body weight change in high fat-fed C57BL/6J mice for 16 weeks. The values are reported as the mean ± SD (n = 6).Normal, 10% fat diet + oral administration of drinking water; Control, 60% fat diet + oral administration of drinking water; EFAY100, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (100 mg/kg); EFAY200, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (200 mg/kg).
a–cMeans with the different letters are significantly different (P < 0.05) by a Duncan’s multiple range test.
|
Table 2
Effects of ethyl acetate fraction from Aster yomena (Kitam.) Honda on body weight gain and food intake in high fat-fed C57BL/6J mice for 16 weeks
The values are reported as the mean ± SD (n = 6).
Normal, 10% fat diet + oral administration of drinking water; Control, 60% fat diet + oral administration of drinking water; EFAY100, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (100 mg/kg); EFAY200, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (200 mg/kg); NS, not significant.
a–cMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple range test.
![]()
Table 3
Effects of ethyl acetate fraction from Aster yomena (Kitam.) Honda on brain, liver, kidney, and epididymal fat weights in high fat-fed C57BL/6J mice for 16 weeks
The values are reported as the mean ± SD (n = 6).
Normal, 10% fat diet + oral administration of drinking water; Control, 60% fat diet + oral administration of drinking water; EFAY100, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (100 mg/kg); EFAY200, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (200 mg/kg).
a–cMeans with the different letters are significantly different (P < 0.05) by Duncan's multiple range test.
![]()
Effect of EFAY on the serum lipid profile
The concentrations of TG, TC, and HDL cholesterol were measured to explore the effect of EFAY on lipid profile in serum. The LDL cholesterol level was calculated using other lipid levels. The HFD-based groups showed a significant increase in the serum concentrations of all kinds of lipids compared to the ND-based group (Fig. 2). In contrast, the serum concentrations TG, TC, and LDL cholesterol in the EFAY-administered groups were decreased significantly. In particular, LDL cholesterol level was reduced with 47.26% inhibition in the EFAY100 group compared to the control group of 100%. EFAY administration, particularly 200 mg/kg, elevated the serum HDL cholesterol concentration compared to the HFD control group. These results suggested that EFAY might improve the lipid profile, which was disrupted by consuming HFD in mice.
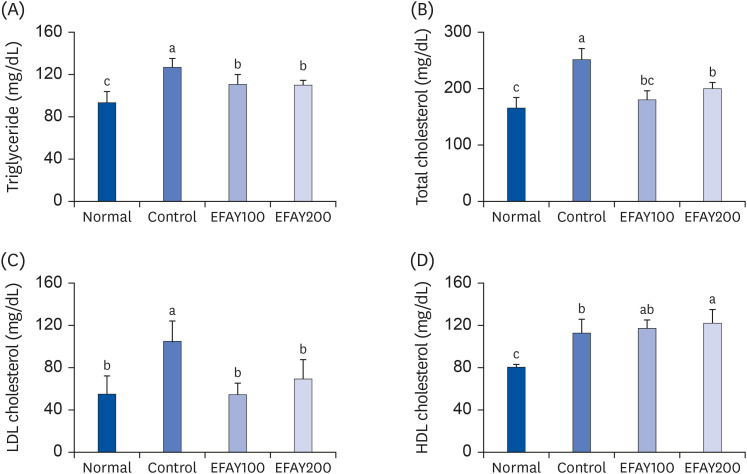 | Fig. 2Effects of ethyl acetate fraction from Aster yomena (Kitam.) Honda on the lipid profile in high fat-fed C57BL/6J mice for 16 weeks. (A) Triglyceride; (B) Total cholesterol; (C) LDL cholesterol; (D) HDL cholesterol. The values are reported as the mean ± SD (n = 6).Normal, 10% fat diet + oral administration of drinking water; Control, 60% fat diet + oral administration of drinking water; EFAY100, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (100 mg/kg); EFAY200, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (200 mg/kg).
a–cMeans with the different letters are significantly different (P < 0.05) by a Duncan’s multiple range test.
|
Effects of EFAY on fasting blood glucose, serum insulin, and HOMA-IR
The fasting blood glucose and serum insulin levels were investigated to evaluate the insulin activity in mice. There was no significance in the initial fasting blood glucose among all groups (Fig. 3A). The HFD increased the fasting blood glucose significantly compared to ND in mice at the end of the experiment. On the other hand, when EFAY was administered orally for 4 weeks, there were significant declines in the fasting blood glucose level at week 16. The serum insulin level in mice was also increased by ingesting HFD, whereas the EFAY treatment produced a significant remarkable decrease in insulin level compared to the HFD control group (Fig. 3B). Next, HOMA-IR was calculated to evaluate insulin resistance in mice. As a result, the HFD mice showed a higher HOMA-IR than the ND mice, which was derived from the increase in fasting blood glucose and serum insulin level (Fig. 3C). The EFAY-administered mice exhibited significantly lower HOMA-IR because of the reduced fasting blood glucose and serum insulin levels compared to HFD control mice. These results suggest that HFD consumption could accompany insulin resistance and obesity, and EFAY supplementation might attenuate insulin resistance by decreasing the fasting blood glucose and serum insulin level.
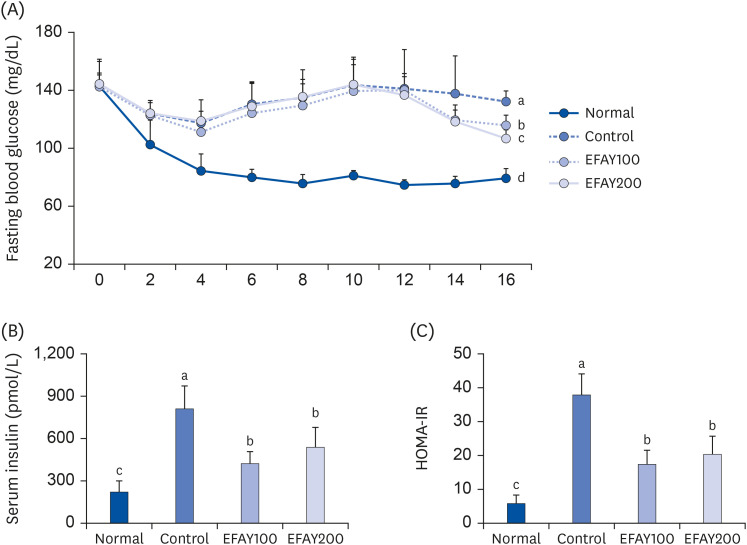 | Fig. 3Effect of ethyl acetate fraction from Aster yomena (Kitam.) Honda on fasting blood glucose (A) serum insulin (B), and HOMA-IR (C) in high fat-fed C57BL/6J mice for 16 weeks. The values are reported as the mean ± SD (n = 6).Normal, 10% fat diet + oral administration of drinking water; Control, 60% fat diet + oral administration of drinking water; EFAY100, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (100 mg/kg); EFAY200, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (200 mg/kg).
a–dMeans with the different letters are significantly different (P < 0.05) by a Duncan’s multiple range test.
|
Effects of EFAY on FAS, p-AMPK/AMPK, and p-ACC/ACC protein expressions in the liver
FAS, p-AMPK/AMPK, and p-ACC/ACC protein expression were measured using Western blot analysis to understand the protective mechanisms of EFAY against lipogenesis and insulin resistance. As a result, the HFD enhanced the protein expressions of FAS compared to ND in the liver of mice (Fig. 4). In response to the administration of EFAY in HFD fed groups, lower FAS protein expression levels were observed. The levels of p-AMPK/AMPK and p-ACC/ACC protein were increased in the HFD-fed mice treated with EFAY the HFD control mice in the liver. These results suggest that EFAY might suppress lipogenesis and insulin resistance by regulating the FAS and AMPK pathway in obesity-induced mice.
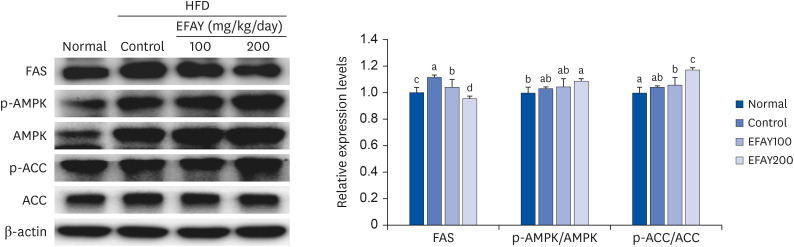 | Fig. 4Effects of the ethyl acetate fraction from Aster yomena (Kitam.) Honda on FAS, p-AMPK/AMPK, and p-ACC/ACC protein expressions in the liver of high fat-fed C57BL/6J mice for 16 weeks. The values are reported as the mean ± SD (n = 6).HFD, high-fat diet; EFAY, ethyl acetate fraction from AY; Normal, 10% fat diet + oral administration of drinking water; Control, 60% fat diet + oral administration of drinking water; EFAY100, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (100 mg/kg); EFAY200, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (200 mg/kg); FAS, fatty acid synthase; p-AMPK, phospho-AMPK; AMPK, AMP-activated protein kinase; p-ACC, phospho-ACC; ACC, acetyl-CoA carboxylase.
a–dMeans with the different letters are significantly different (P < 0.05) by a Duncan’s multiple range test.
|
Effects of EFAY on serum AST, ALT, and hepatic lipid peroxidation
The levels of AST and ALT in serum and lipid peroxidation in the liver were investigated to evaluate HFD-induced hepatic damage in mice (Fig. 5). The consumption of HFD increased the serum AST and ALT levels in mice. On the other hand, treatment of HFD-fed mice with EFAY reduced the serum AST and ALT levels significantly. The effects of EFAY against lipid peroxidation were also determined by measuring the MDA concentration in the liver. Consistent with the serum AST and ALT levels, HFD increased hepatic MDA generation compared to ND, whereas it was decreased by EFAY administration. These results suggest that EFAY may protect the liver from HFD-induced hepatic injury by inhibiting lipid peroxidation and consequently blocking the leakage of AST and ALT into the serum.
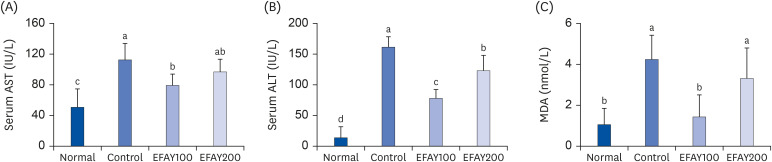 | Fig. 5Effects of the ethyl acetate fraction from Aster yomena (Kitam.) Honda on AST (A), ALT (B), and MDA (C) levels in high fat-fed C57BL/6J mice for 16 weeks. The values are reported as the mean ± SD (n = 6).AST, aspartate transaminase; Normal, 10% fat diet + oral administration of drinking water; Control, 60% fat diet + oral administration of drinking water; EFAY100, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (100 mg/kg); EFAY200, 60% fat diet + oral administration of ethyl acetate fraction from Aster yomena (Kitam.) Honda (200 mg/kg); ALT, alanine aminotransferase; MDA, malondialdehyde.
a–dMeans with the different letters are significantly different (P < 0.05) by a Duncan's multiple range test.
|
Go to :
DISCUSSION
The excessive consumption of high calories and low energy expenditure are major causative factors for obesity and its comorbidities [22]. Obesity is characterized as an excessive accumulation of adipose tissue in the body [23]. When adipocyte becomes overly accumulated, it may result in adipocyte endocrine dysfunction, promoting dyslipidemia [24]. Adipose tissue, as an immune organ, produces a variety of pro-inflammatory molecules, such as adipokines (e.g., leptin and adiponectin) and cytokines (e.g., tumor necrosis factor-α and interleukin) [25]. These pro-inflammatory factors have been implicated as active participants in the development of metabolic disorders, such as insulin resistance [26]. Insulin resistance is an impaired sensitivity to insulin of its target organs, such as adipose tissue, muscle, and liver [262728]. Pro-inflammatory stimuli released from adipose tissues lead to chronic low-grade inflammation in the body and the development of systemic insulin resistance [29]. Previous studies have reported that AY inhibited adipogenesis and inflammation in adipocytes and macrophages by reducing pro-inflammatory stimuli, including nuclear factor kappa-light-chain-enhancer of activated B cells and mitogen-activated protein kinases [1230]. In addition, obesity-induced by a HFD leads to an inflammatory response in adipose tissue and impairs hepatic lipid metabolism. The anti-adipocyte differentiation activity of EFAY in 3T3-L1 cells was previously investigated [14]. On the other hand, the effects of EFAY on hepatic lipid metabolism and insulin resistance in obese mice have not been investigated. Therefore, this study determined the effects of EFAY on the hepatic lipid metabolism and insulin resistance in HFD-fed obese mice.
In the body weight, there was no significance between EFAY100 and 200. According to Lee et al. (2017), the administration of AY at a dose of 250 and 500 mg/kg/day to high-fat diet-induced obese mice decreased the body weight significantly compared to the mice administered AY at 100 mg/kg/day [31]. Therefore, a higher administered concentration of EFAY is needed.
The consumption of HFD can trigger the standard features of obesity and insulin resistance in rodents [32333435]. As C57BL/6J mice have susceptibility to HFD-induced obesity and insulin resistance, feeding HFD to C57BL/6J mice is a suitable model for identifying the mechanisms underlying obesity and insulin resistance [363738]. Previous studies showed that C57BL/6J mice showed more significant weight gain when the mice were fed 60% HFD for 16 weeks [3940]. These results showed increased body and organ weight gain by consuming a HFD in mice. On the other hand, the EFAY treatment of HFD mice decreased the body and organ weights compared to the HFD control mice.
Several studies reported that prolonged HFD led to obesity and insulin resistance [35414243]. Therefore, the effects of EFAY on HFD-fed obesity mice were subsequently investigated by measuring the lipid profile in the serum. The HFD-based control mice degenerated dyslipidemia that was associated with higher TG, TC, and LDL cholesterol concentrations. In contrast, the administration of EFAY to HFD mice ameliorated dyslipidemia by reducing the levels of TG, TC, and LDL cholesterol. Regarding LDL-cholesterol, the EFAY200 group had higher levels than the EFAY100 group. On the other hand, other lipid levels, such as TG and HDL–cholesterol, had a concentration-dependent effect. According to Lee et al. [31], there was no significant difference in the lipid profiles between the 100 and 250 mg/kg groups. In addition, caffeic acid, one of the major active compounds in EFAY, lowered the serum TG and TC in HFD-fed mice [44]. The observations of the fasting blood glucose and serum insulin levels confirmed that HFD caused dietary-induced insulin resistance in mice. On the other hand, EFAY administration with feeding HFD improved the fasting blood glucose and serum insulin levels. In the blood glucose levels, EFAY200 decreased blood glucose (106.5 3 mg/dL) significantly compared to EFAY100 (115.3 mg/dL) at week 16. This leads to a decline of HOMA-IR, which was used as an index for assessing insulin resistance [45]. The HOMA-IR and serum insulin levels in the EFAY200 group (20.46 and 540.2 pmol/L, respectively) were higher than EFAY100 (17.6 and 425.3 pmol/L, respectively), but the differences were not significant. These results suggest that EFAY might help improve insulin resistance.
Excessive lipid accumulation results from an imbalance between synthesis oxidation of lipids [45]. The liver is a central organ where lipid metabolism is modulated and crucial in lipid synthesis [46]. FAS, a hepatic lipogenic enzyme, is involved in fatty acid synthesis [47]. HFD-fed mice showed higher FAS protein expression than ND-fed mice in the liver, whereas protein expressions were down regulated by the EFAY treatments. In the cellular system, the ethanol extract of AY inhibited FAS mRNA expression in 3T3-L1 adipocytes [12]. These results suggested that EFAY might reduce the production of lipids in the liver and decrease lipid accumulation. Next, AMPK pathway-related protein expression was measured to understand how EFAY regulates insulin resistance. AMPK modulates the glucose and lipid metabolisms in the liver. Activated AMPK inhibits hepatic glucose production, lipogenesis, and fatty acid oxidation [48]. Therefore, AMPK has attracted considerable attention as a therapeutic target for treating metabolic abnormalities, including obesity and type 2 diabetes [49]. ACC is a downstream target of AMPK and increases phosphorylation when AMPK is activated [50]. The oral administration of EFAY to HFD-fed mice increased the levels of AMPK and ACC phosphorylation, suggesting that EFAY up-regulates the AMPK pathway. These are consistent with the observation that EFAY mediates the activation of AMPK and ACC in 3T3-L1 adipocytes by elevating AMPK and ACC phosphorylation [14]. Previous studies reported that plant extracts like EFAY also activated AMPK and its downstream substrates, which helped regulate the glucose metabolism and protect hyperglycemia in obese mice [5152]. In the ACC expression, EFAY200 did not show significance compared to EFAY100. On the other hand, EFAY200 significantly activated p-ACC, which led to an increase in p-ACC/ACC. Han et al. [12] also reported an AY treatment at concentrations of 50–200 μg/mL in differentiated 3T3-L1 cells significantly enhanced p-ACC expression [12]. These results show that EFAY might ameliorate the glucose metabolism in HFD-fed mice by inhibiting the AMPK pathway in the liver.
Several markers can measure hepatic injuries, such as AST, ALT, and MDA. AST and ALT reflect the damage to hepatocytes, which could be caused by hyperlipidemia [535455]. MDA is generated when hepatic lipids are oxidated by hepatic injury, which could be triggered by obesity [5657]. These results show that HFD elevated the AST, ALT, and MDA levels, indicating that HFD caused hepatic injury. EFAY reversed those results in HFD-fed mice, suggesting that EFAY might protect the liver from hepatic injury induced by feeding mice a HFD.
This study examined the biological activity of AY at concentrations of 100–500 mg/kg in mice [29]. Furthermore, there was no toxicity until the concentration was 1 g/kg in mice [58]. Although there was no toxicity at the 200 mg/kg in other experiments, the EFAY200 group showed higher AST and ALT levels than the EFAY100 group. The higher AST and ALT of EFAY200 may be related to higher MDA levels; hence, an additional toxicity test is needed.
Natural agents contain active substances, which show various bioactivities in the body. AY contains several active substances, such as esculetin, caffeic acid, and apigenin [9]. Esculetin decreased adipogenesis by regulating the AMPK pathway in 3T3-L1 adipocytes [59]. Caffeic acid prevented hyperlipidemia and obesity in C57BL/6 mice by modulation of hepatic lipogenesis gene expression [60]. Apigenin also alleviated dyslipidemia, hepatic steatosis, and insulin resistance in the HFD-induced obese mice [61]. Therefore, the anti-obesity effect might come from the active substances from AY.
In conclusion, these results suggest that EFAY decreased bodyweight, which is linked with improved lipid profile and insulin resistance in the HFD-fed mice. These effects might be derived from the deactivation of FAS and activation of the AMPK pathway in the liver. Therefore, AY could be a natural agent for preventing obesity and its comorbidities, such as insulin resistance.
Go to :
 XML Download
XML Download