

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Skeletal muscles have key roles in supporting carbohydrate metabolism and energy expenditure as they contain numerous mitochondria [123]. Mitochondria are entirely responsible for increasing endurance-exercise tolerance of skeletal muscles to fatigue and improving energy metabolism, resulting in enhanced exercise performance [4]. This process generates cellular energy in the form of ATP production, intracellular calcium homeostasis, amino acid metabolism, oxidant–antioxidant balance, and apoptosis [5]. Skeletal muscle consists of 4 major myosin heavy chain (MyHC) fibers depending on the expression of slow-twitch with Myh7 (type I), fast-twitch with Myh2 (type IIa), Myh1 (type IIx), or Myh4 (type IIb) [6]. Myh7 fibers (slow-twitch, oxidative) are rich in mitochondria, resistant to fatigue, and sustain long-term energy demands. In addition, they facilitate fatty acid oxidation and more efficient ATP production. In contrast, Myh4 fibers (fast-twitch) contain glycogen and glucose as energy sources. These mitochondria-poor fibers primarily rely on rapid glycolysis for ATP production; thus, they are more sensitive to fatigue, which is attributed to sudden activity in a short duration [7]. Other fiber types such as Myh2 (fast-twitch, oxidative) and Myh1 (fast-twitch, oxido-glycolytic) show a combination of characteristics intermediate between those of Myh7 and Myh4 fibers [8910].
Mitochondria undergo continuous biogenesis, generally via expression of Ppargc1a, the gene that encodes peroxisome proliferator-activated receptor-gamma coactivator 1α (PGC-1α) [11]. Ppargc1a overexpression in mice has been reported to enhance mitochondrial concentrations, exercise capacity, oxidative capacity, and fatigue resistance [12]. PGC-1α is a co-activator of a broad range of nuclear transcription factors, including nuclear respiratory factor (NRF) 1 and 2, estrogen-related receptor α (ERRα), as well as myocyte enhancer factor-2 (MEF-2). PGC-1α drives mitochondrial biogenesis via the expression of the gene that encodes mitochondrial transcription factor A (Tfam), which directly stimulates mitochondrial DNA replication and transcription [131415]. Key steps in several exercise-induced pathways of the skeletal muscle involve the activation of AMP-activated protein kinase (AMPK), which has been shown to regulate the phosphorylation and activation of PGC-1α [16].
Ergogenic aids, such as vitamins, proteins, polyphenols, amino acids, creatine monohydrates, and herbs, help improve physical performance, delay fatigue, enhance energy production, regulate oxidative stress, and accelerate metabolic activity [171819]. Gynostemma pentaphyllum (Thumb.) Makino has been widely used as a traditional tea or herb medicine in Asia, including China, South Korea, and Japan. G. pentaphyllum is used to treat various diseases, including hepatitis, diabetes, and cardiovascular diseases [3]. Additionally, it reportedly elicits various effects and possesses numerous pharmacological properties, such as anti-inflammatory, lipid metabolism-regulatory, anti-diabetic, neuroprotective, and anti-obesity properties [2021222324252627]. Dammarane gypenosides are considered the major bioactive constituents in G. pentaphyllum and possess hepatoprotective, antioxidant, anti-obesity, anti-inflammatory, hypolipemic, hypoglycemic, and anticancer properties [2128293031]. Moreover, polysaccharides or gypenosides from G. pentaphyllum have been investigated for their anti-fatigue and anti-oxidative activities [3233]. Previously, we demonstrated the anti-obesity effect of G. pentaphyllum extract (GPE) and gypenoside L (GL), a saponin isolated from G. pentaphyllum. Both GPE and GL boosted glucose uptake and GLUT4 expression via activation of the AMPK-acetyl-CoA carboxylase (ACC) signaling pathway and promoted mRNA expression of Ppargc1a in skeletal muscle cells [34]. However, whether GPE and GL regulate the mitochondrial biogenic pathway via PGC-1α expression in C2C12 myotubes remains unclear.
Thus, we hypothesized that GPE and its active compound GL activate the PGC-1α pathway and regulate various genes associated with exercise performance, such as those governing muscle differentiation, muscle fiber switching, lactate metabolism, and energy metabolism. To this end, we investigated the effect of GPE and GL on muscle differentiation and mitochondrial metabolism in murine C2C12 myotubes.
MATERIALS AND METHODS
Chemicals and reagents
Creatine monohydrate was purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), penicillin/streptomycin solution, horse serum (HS), and phosphate-buffered saline (PBS) were obtained from GIBCO BRL Life Technologies (Grand Island, NY, USA). Primary anti-AMPKα (total or phosphor-form), anti-SIRT1 (total or phosphor-form), and anti-p38 (total or phosphor-form) antibodies, as well as secondary antibodies against anti-rabbit and anti-mouse immunoglobulin G (IgG) horseradish peroxidase (HRP)-linked antibodies, were obtained from Cell Signaling Technology (Beverly, MD, USA). Anti-Myh1 and anti-NRF2 (total or phosphor-form) antibodies were obtained from Abcam (Abcam Inc., Cambridge, UK). PGC-1α and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). MitoTracker® Red CMXRos and ProLong™ Gold Antifade Mountant with 4′,6-diamidino-2-phenylindole (DAPI) were purchased from Invitrogen (Carlsbad, CA, USA).
Preparation of GPE
G. pentaphyllum was purchased from Hunan Tea Group Co., Ltd (Changsha, Hunan, China), and a voucher specimen (20210301-1) was identified by Professor Zhou Ribao, College of Medicine, Hunan University of Traditional Chinese Medicine. The GPE and GL were prepared at BTC Corporation, according to a previously described method [34]. In brief, 1 kg of dried G. pentaphyllum leaves were extracted with a hot water (20 L) and 50% EtOH aqueous solution (15 L). After each extraction, the extracts were combined, concentrated, and filtered. Quantification of GL in GPE was performed using high-performance liquid chromatography (HPLC) with a diode-array detector. GPE contains active components such as GL (1.8%, w/w), gypenoside LI (GLI) (1.4%, w/w), and ginsenoside Rg3 (0.15%, w/w). A representative chromatogram of the standard references and the corresponding peaks in the GPE are presented in Supplementary Figs. 1 and 2.
Cell culture and treatment procedures
Murine C2C12 myoblast cells were purchased from American Type Culture Collection (Manassas, VA, USA). The C2C12 myoblasts were cultured and maintained in growth medium consisting of DMEM supplemented with 10% (v/v) FBS and 1% (v/v) penicillin/streptomycin solution under a humidified atmosphere of 95% air and 5% CO2 at 37°C. For differentiation induction, C2C12 myoblasts at 85–95% confluence were incubated in differentiation medium with DMEM containing 2% (v/v) HS every 2 days. The obtained C2C12 myotubes were treated with/without GPE (20 μg/mL) or GL (0.36 μg/mL). Creatine monohydrate (CrM, 5 μg/mL) was used as a positive control. All controls were incubated with equal volumes of the vehicles used for the GPE, GL, and CrM treatments.
Quantitative reverse transcription polymerase chain reaction assay (qRT-PCR)
After 4 days of cell differentiation, total RNA was isolated by using an RNeasy® Plus Mini kit (Qiagen, Valencia, CA, USA) according to the manufacturer's instructions. First-strand reverse transcription was performed with 2 μg total RNA using a Hyperscript™ RT master mix kit (GeneAll Biotechnology, Seoul, Korea). The levels of transcripts were determined by performing qRT-PCR on a Rotor-Gene 3000 instrument (Corbett Research, Mortlake, Australia) and using a Rotor-Gene™ SYBR Green Kit (Qiagen) as a double-stranded DNA-specific dye, according to the manufacturer's instructions. The primers used in the present study were ordered from and synthesized by Bioneer Co. (Daejeon, Korea); their sequences are listed in Table 1. The qRT-PCR consisted of 41 cycles, comprising 1 cycle of 3 min at 94°C and 40 cycles of 10 s at 95°C, 15 s at 60°C, and 20 s at 72°C. The amount of mRNA expression was normalized with the housekeeping gene Gapdh to determine the relative expression ratios for each mRNA gene expression relative to the control group. For statistical analysis purposes, all experiments included a minimum of three independent PCR reactions.
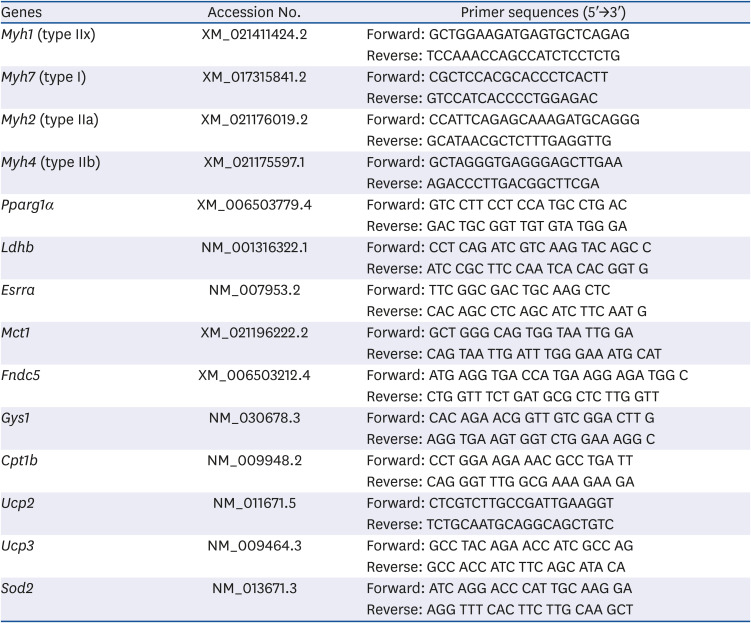
Table 1
Primer sequences used in qRT-PCR for the analysis of mRNA expression
![]()
Western blot analysis
After 7 days of differentiation, cells were washed, harvested, and lysed with an ice-cold lysis buffer (20 mM HEPES pH 7.5, 150 mM NaCl, 1 mM EGTA, 1 mM EDTA, 100 mM NaF, 10 mM sodium pyrophosphate, and 1% Triton X-100) containing 0.2 mM PMSF, 5 mM iodoacetic acid, and a phosphatase/protease inhibitor (iNtRON Biotechnology, Inc., Seongnam, Korea). The lysates were collected, and protein concentrations were quantified using a BCA protein assay kit (Thermo Scientific, Rockford, IL, USA). Equal amounts of protein (50 μg) were separated by 10% (v/v) Sodium dodecyl sulphate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride nitrocellulose membranes by electroblotting. The membrane blots were blocked overnight at 4°C in blocking buffer (5% skim milk in Tris-buffered saline with 0.1% Tween-20 [TBST]) and incubated with the following primary antibodies: Myh1, p-AMPK α (Thr172), AMPKα, p-p-38, p-38, p-SIRT1, SIRT1, PGC-1α, p-NRF2, NRF2, and GAPDH, (1:1,000, in 5% skim milk in TBST). The blots were washed thrice with TBST and incubated with a horseradish peroxidase-conjugated anti-rabbit IgG HRP-linked or anti-mouse IgG HRP-linked antibody (1:5,000, in 5% skim milk in TBST). Subsequently, the signal was visualized using an enhanced chemiluminescence technique with the Luminata™ Forte Western Chemiluminescent HRP Substrate (Millipore, Billerica, MA, USA), according to the manufacturer's protocol. Band intensity was quantified using an ImageQuant™ LAS 500 imaging system (GE Healthcare Bio-Sciences AB, Uppsala, Sweden).
Confocal microscopy and immunocytochemistry
C2C12 myoblasts (2 × 104 cells/mL) were seeded on an ibiTreat μ-Slide 8-well polymer coverslip (Ibidi GmbH, Martinsried, Germany) and differentiated using a common culture method. After differentiation with GPE, GL, or CrM, the coverslips were washed with 1× PBS and fixed with 4% formaldehyde/PBS for 15 min at room temperature, after which the myotubes were permeabilized with 0.5% Triton X-100 in PBS for 15 min. After washing thrice with 1× PBS, the myotubes were blocked with 3% bovine serum albumin in 1× PBS for 1 h at room temperature and immunostained with Myh1 primary antibody (1:3,000) overnight at 4°C. The coverslips were labeled with AlexaFlour® 488 goat anti-rabbit IgG secondary antibody (1:100) for 1 h at room temperature. MitoTracker® Red CMXRos and ProLong™ Gold Antifade Mountant with DAPI (Invitrogen) prepared in PBS were added at final concentrations of 50 nM and 1 µM, respectively. Myh1 expression was visualized by fluorescence microscopy (LSM 800; Carl Zeiss, Jena, Germany).
Measurement of mitochondrial reactive oxygen species (ROS) production
The intracellular production of ROS in C2C12 myotubes was assayed using the 2′7′-dichlorofluorescein diacetate (DCF-DA) assay, as previously described [35]. C2C12 myoblasts were cultured for 24 h in a 96-well plate and treated with or without GPE, GL, or CrM for 1 h. Myotubes were washed once with PBS and then incubated for 3 h with 50 µM tert-butyl hydrogen peroxide (tBHP). Cells were washed with PBS and stained with 20 mM DCF-DA for 45 min. The level of DCF fluorescence was detected by a fluorescence reader at excitation/emission wavelengths of 495 nm/529 nm. Data are presented as relative fluorescence units/well.
Statistical analysis
All data represent five replicate independent experiments. Statistical analysis of the collected data was performed using GraphPad Prism (GraphPad Prism Software Inc., La Jolla, CA, USA). All values are expressed as the mean
±
SE. Statistical analyses were performed using the Student's t-test or performing a one-way analysis of variance. P-value < 0.05 was considered indicative of a statistically significant difference.
RESULTS
Treatment with GPE and GL enhance myoblast differentiation
To investigate the role of GPE and GL in myoblast differentiation, we first examined their effects on cell viability during a 4-day differentiation period. The concentrations of GPE and GL used in the treatment manifested no cytotoxic effects in C2C12 myotubes (Fig. 1). This was further confirmed by using qRT-PCR to assess the mRNA expression of specific markers (Myh1, Myh7, Myh2, and Myh4). There was a noticeable increase in the Myh1 mRNA expression level in cells treated with GPE and GL compared to that of control cells (Fig. 2A). The mRNA expression level of Myh1 following treatment with GPE and GL was also higher than that observed following treatment with CrM. Similarly, the Myh7 expression level was significantly elevated in cells treated with GPE and CrM (Fig. 2B); however, the expression level of the CrM group was higher than that of the GPE group. In addition, GPE, GL, and CrM caused an elevation in the mRNA levels of Myh2 in C2C12 myotubes (Fig. 2C), but there was no significant difference among the groups. By contrast, the Myh4 mRNA expression level showed a significant reduction following GPE treatment (Fig. 2D). We next performed western blot analysis to examine the effects of GPE and GL on differentiation in C2C12 cells. As presented in Fig. 3A, myoblast cells, following treatment with GPE and GL, exhibited a gradual morphological change into an elongated shape and formed mature multi-nucleated myotubes. The GPE-treated cells showed increased Myh1 protein expression levels (Fig. 3B). Immunocytochemistry confirmed that treatment with GPE and GL augmented Myh1 expression, with a resultant elongated and widened cylindrical cell shape (Fig. 3C). The fluorescence signals from MitoTracker and Myh1 were quantified using Image J software. The fluorescence intensity of MitoTracker showed values similar to those of the treatment group cells. The fluorescence signal intensity increased with the stage of progression of muscle differentiation. The signals emitted from GPE-treated cells were significantly stronger than those emitted from GL- and CrM-treated cells (Fig. 4A and B). Taken together, these results demonstrated that GPE and GL augment the expression of differentiation markers in C2C12 cells.
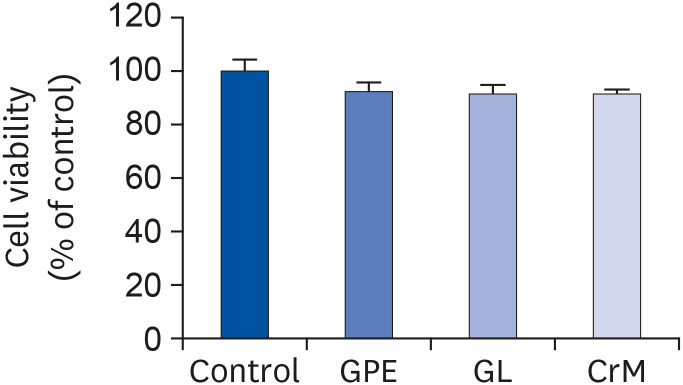
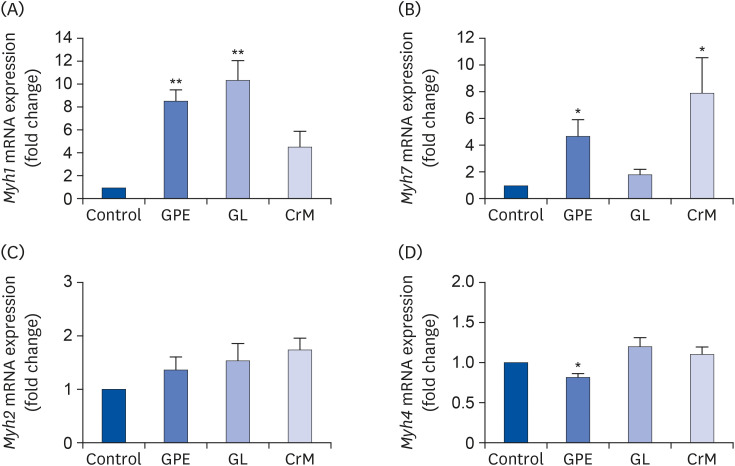
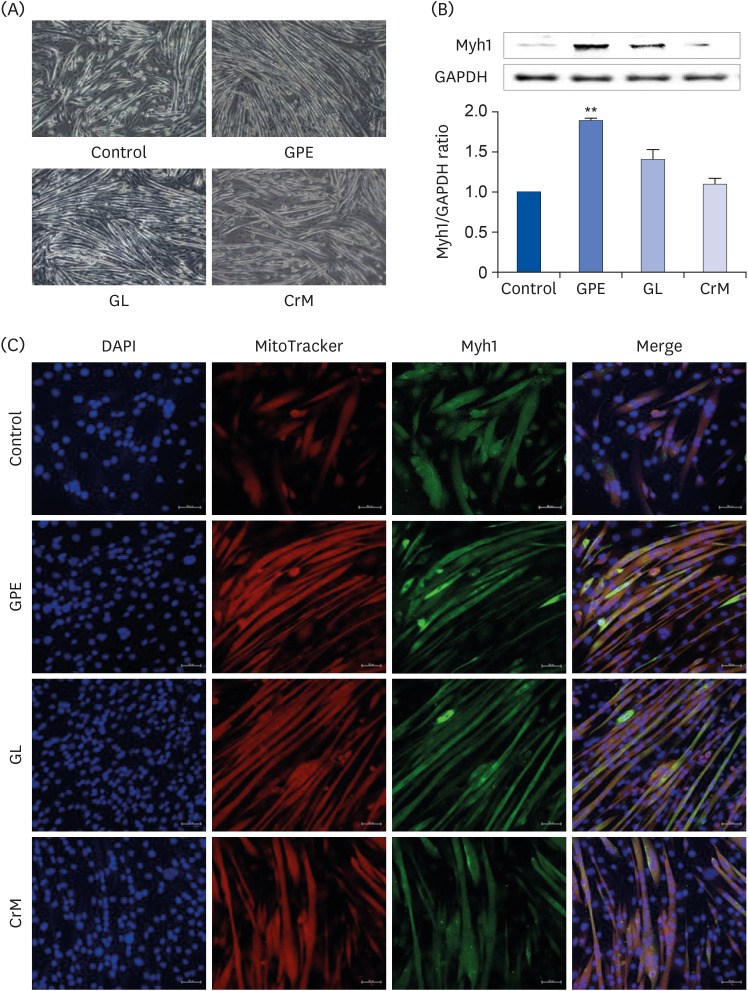
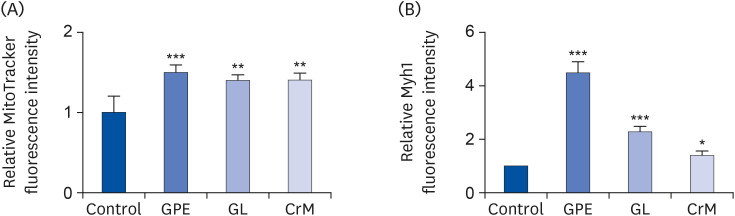
Fig. 1
Cell-toxicity analysis result. C2C12 cells were incubated in differentiation medium with GPE, GL, or CrM for 4 days. MTT assays showed no cytotoxic effects from the GPE, GL, and CrM treatments in C2C12 cells. All data represent five replicates (n = 5), and values are shown as means ± SE.
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM; creatine monohydrate.
![]()
Fig. 2
Effect of GPE and GL on the mRNA expression of MyHC genes in C2C12 myotubes. C2C12 cells were incubated in differentiation medium with GPE, GL, or CrM for 4 days. The mRNA expression levels of Myh1 (A), Myh7 (B), Myh2 (C), and Myh4 (D) were determined by qRT-PCR. All data represent five replicates (n = 5), and values are shown as means ± SE.
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM, creatine monohydrate.
*P < 0.05 and **P < 0.01 indicate a significant difference from the control group.
![]()
Fig. 3
Effect of GPE and GL on the differentiation of myoblasts into myotubes. (A) C2C12 cells were incubated in differentiation medium with GPE, GL, or CrM for 4 (immunofluorescence staining) and 7 (western blot) days. (B) Protein expression of Myh1 was assessed by western blotting analysis. GAPDH was used as a loading control. All data represent five replicates (n = 5), and values are shown as means ± SE. (C) Immunofluorescence staining of Myh1 in C2C12 myotubes. The cells were fixed and immunostained for Myh1 (green), MitoTracker (red), and DAPI (blue). Myotube formation was observed by fluorescence microscopy (original magnification, 200×) (scale bar = 50 μm).
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM, creatine monohydrate; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; DAPI, 4′,6-diamidino-2-phenylindole.
**P < 0.01 indicate a significant difference from the control group.
![]()
Fig. 4
Quantification of MitoTracker and Myh1 staining intensity in C2C12 cells. Differentiated C2C12 cells at day 4 were stained with MitoTracker (A) and Myh1 (B). Fluorescence intensity of immunofluorescence sections was quantified using Image J software. The results are expressed as means ± SEM (n = 5).
SEM, standard error of the mean; GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM, creatine monohydrate.
*P < 0.05, **P < 0.01, and ***P < 0.001 indicate a significant difference from the control group.
![]()
Effect of GPE and GL on lactate metabolism
To investigate the influence of GPE and GL on the molecular mechanism involved in lactate metabolism, we performed qRT-PCR. As shown in Fig. 5A, GPE and GL significantly augmented the mRNA expression of Ppargc1a by 8.18-fold and 11.75-fold, respectively, compared to the corresponding expression level in control myoblasts. C2C12 cells were treated with GPE and GL to examine the effect of GPE and GL on fatigue-related genes encoding lactate dehydrogenase (LDH)-B, ERRα, and monocarboxylate transporter (MCT) 1, which regulate lactate production and clearance in myotubes [36]. As shown in Fig. 5B-D, we determined that GPE and GL treatment markedly increased Esrra and Mct1 mRNA expression; however, it resulted in a non-significant increase in Ldhb mRNA expression. These results indicated that GPE regulates lactate metabolism by promoting Esrra and Mct1 expression.
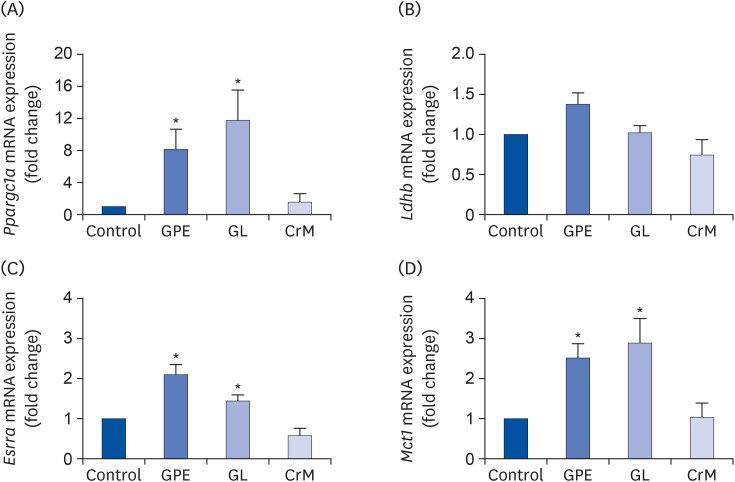
Fig. 5
Effects of GPE and GL on lactate metabolism in C2C12 myotubes. Differentiated C2C12 myotubes were treated with/without GPE, GL, or CrM. The mRNA expression levels of Ppargc1a (A), Ldhb (B), Esrra (C), and Mct1 (D) were evaluated by qRT-PCR. Target gene expression levels were normalized to the Gapdh expression level. Data are expressed as means ± SE (n = 5).
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM, creatine monohydrate; qRT-PCR, quantitative reverse transcription polymerase chain reaction.
*P < 0.05 indicate a significant difference from the control group.
![]()
Effects of GPE and GL on mitochondrial metabolism
We investigated whether GPE and GL regulate metabolic genes at the transcriptional level. As shown in Fig. 6, treatment of C2C12 myotubes with GPE and GL resulted in a significant elevation in the mRNA expression levels of fibronectin type III domain-containing protein 5 (Fndc5) (Fig. 6A), which is related to adipocyte browning, and glycogen synthase (Gys) (Fig. 6B). Furthermore, we observed that GPE and GL promote lipid metabolism by increasing the mRNA expression of carnitine palmitoyltransferase 1b (Cpt1b); however, only GPE produced a nearly 3-fold increase and a statistically significant difference compared to the corresponding parameters in untreated cells (Fig. 6C).
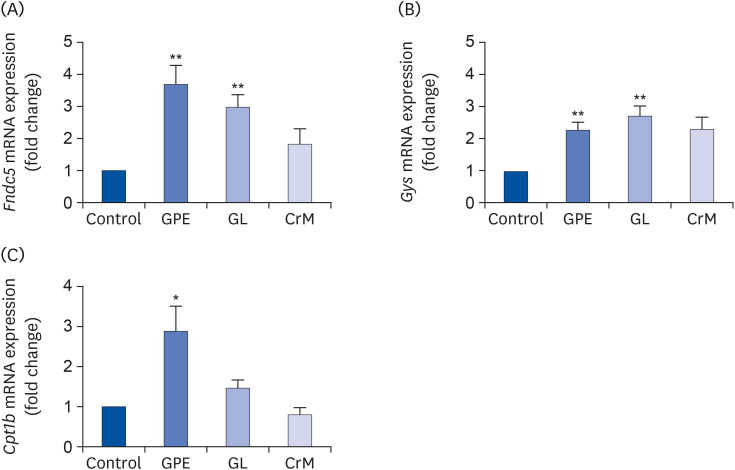
Fig. 6
Effects of GPE and GL on adipocyte browning, glycogen synthesis, and lipid metabolism in C2C12 myotubes. Differentiated C2C12 myotubes were treated with GPE, GL, or CrM for 4 days. The mRNA expression levels of genes related to adipocyte browning (Fndc5) (A), glycogen synthesis (Gys) (B), and lipid metabolism (Cpt1b) (C) were determined by qRT-PCR. Target gene expression levels were normalized to the Gapdh expression level. Each value represents the mean ± SE (n = 5).
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM, creatine monohydrate; Fndc5, fibronectin type III domain-containing protein 5 gene; Gys, glycogen synthase gene; qRT-PCR, quantitative reverse transcription polymerase chain reaction.
*P < 0.05 and **P < 0.01 indicate a significant difference from the control group.
![]()
Effect of GPE and GL on AMPK activation
To further elucidate the molecular mechanism(s) underlying the GPE- and GL-induced changes in muscle metabolism, we evaluated their effects on the AMPK pathway. As shown in Fig. 7A, treatment with GPE significantly increased the phosphorylation of the AMPK α-subunit at Thr172, which is within the active site of the AMPK-α subunit. In addition, GPE and GL activated p38 MAPK, a critical regulator of glucose uptake (Fig. 7B) [37]. Our previous results indicated that GPE and GL mediate glucose metabolism and metabolic functions via AMPK and p38 MAPK phosphorylation [38]. Activation of AMPK reportedly increases SIRT1 expression in skeletal muscles [39], while AMPK directly phosphorylates and activates PGC-1α by SIRT1, which deacetylates the lysine-site of PGC-1α and activates downstream genes related to energy expenditure [40]. Herein, we determined that treatment with GPE and GL in C2C12 myotubes resulted in increased protein levels of phosphorylated SIRT1 (Fig. 7C) and PGC-1α (Fig. 7D). Consequently, it was inferred that the effects of GPE and GL are mediated via the activation of AMPK/p38, SIRT1, and PGC-1α.
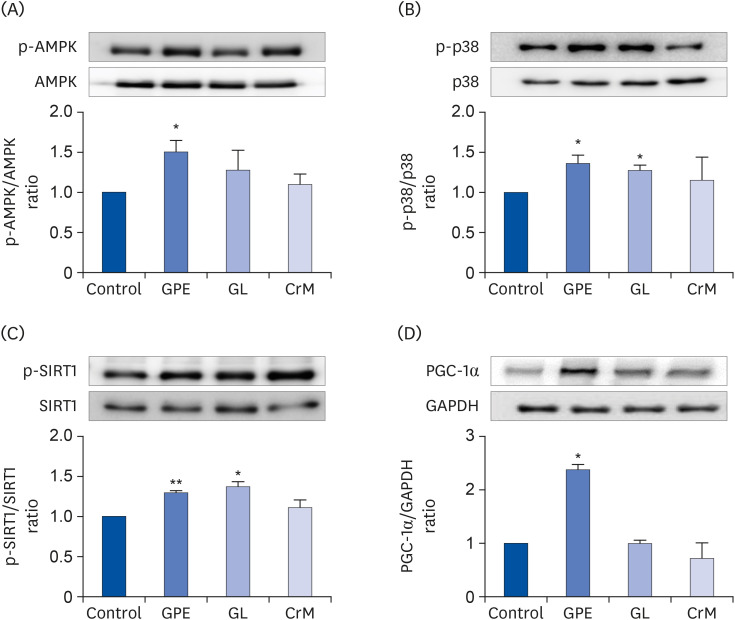
Fig. 7
Effect of GPE and GL on the AMPK/p38 MAPK/SIRT1/PGC-1α signaling pathway. C2C12 cells were cultured for 4 days in differentiation medium supplemented with/without GPE, GL, or CrM. The ratios of p-AMPK/AMPK (A), p-p38/p38 (B), p-SIRT1/SIRT1(C), and PGC-1α/GAPDH (D) were determined. Each bar represents the mean ± SE (n = 5) relative to the control group.
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM, creatine monohydrate; AMPK, AMP-activated protein kinase; PGC-1α, peroxisome proliferator-activated receptor-gamma co-activator 1α; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
*P < 0.05 and **P < 0.01 indicate a significant difference from the control group.
![]()
Effect of GPE and GL on oxidative stress-related gene expression
We next examined whether GPE and GL protect muscle cells from oxidative stress by activating related gene expression. In Fig. 8A and B, it is shown that treatment with GPE and GL significantly induced the mRNA expressions of the fatty acid metabolism/mitochondrial biogenesis markers, uncoupling protein (UCP) 2 (Ucp2) and 3 (Ucp3). The endogenous antioxidant response element Sod2 was also significantly upregulated, by approximately 1.6-fold following treatment of myotubes with GL (Fig. 8C). Given that NRF2 phosphorylation at Ser40, which is induced by several kinases, is a crucial process in its nuclear translocation [414243], we investigated the phosphorylation of NRF2 by performing western blot analysis. The results shown in Fig. 8D indicated that the ratio of NRF2 phosphorylation/total NRF2 was markedly increased following treatment with GPE and GL. The effects of GPE and GL on oxidative stress protective mechanisms are likely mediated by an intrinsic antioxidant capacity. Thus, we investigated whether GPE and GL indeed mediate oxidative stress in muscle cells by assessing ROS generation. Intracellular ROS was detected with DCF-DA, as shown in Fig. 8E. In C2C12 myotubes, tBHP treatment significantly increased intracellular ROS by 2.3-fold compared to the ROS level recorded in control cells. However, when the tBHP-treated C2C12 cells were stimulated with GPE and GL, the ROS levels were significantly reduced (Fig. 8E), indicating that GPE and GL treatments induce an intracellular anti-oxidative response.
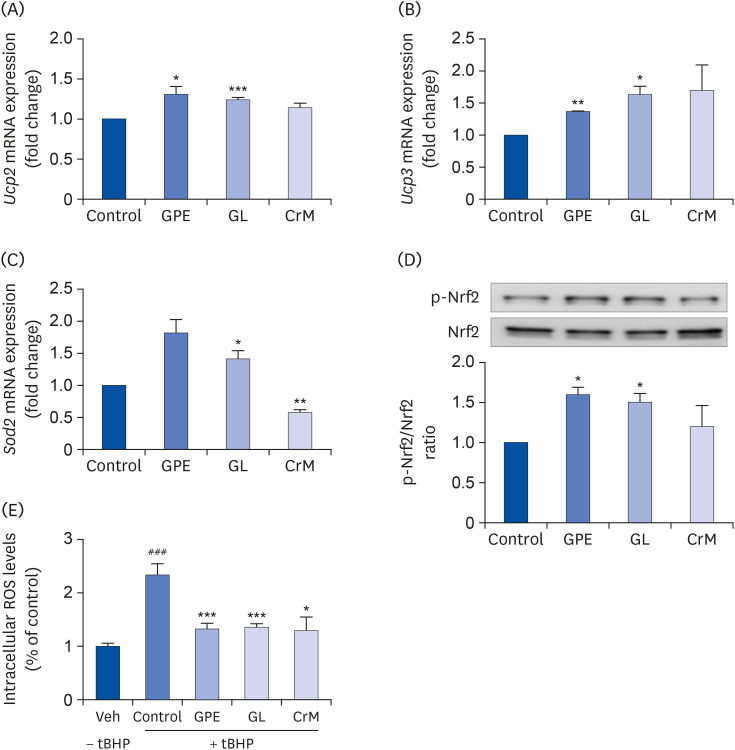
Fig. 8
Effect of GPE and GL on the relative mRNA expression level of oxidative stress-related genes in C2C12 myotubes. C2C12 cells were treated with GPE, GL, or CrM for 4 days. The mRNA levels of Ucp2 (A), Ucp3 (B), Sod2 (C), and the protein levels of phosphorylated NRF2 and NRF2 (D) were analyzed by qRT-PCR and western blot assay, respectively. Gapdh served as an internal control. The ratio of p-NRF2/NRF2 was determined. (E) Effects of GPE and GL on ROS levels in C2C12 myotubes. Cells were studied under normal conditions (Veh), treatment with tBHP for 3 h (Control), and combinations of tBHP with GPE, GL or CrM. ROS levels are presented as fold changes from control cells. All values are presented as the mean ± SE (n = 5).
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; CrM, creatine monohydrate; NRF, nuclear respiratory factor; qRT-PCR, quantitative reverse transcription polymerase chain reaction; ROS, reactive oxygen species; tBHP, tert-butyl hydrogen peroxide.
*P < 0.05, **P < 0.01, and ***P < 0.001 indicate a significant difference from control group. ###
P < 0.001 Indicate significant difference from tBHP alone.
![]()
DISCUSSION
G. pentaphyllum has been reported to exert anti-inflammatory, lipid metabolism-regulatory, anti-diabetic, neuroprotective, and anti-obesity effects [202122232425262734]. Gypenosides are a class of major bioactive compounds in GPE and have been evaluated for their beneficial cholesterol-lowering, antioxidant, hypoglycemic, and tumor-suppressive effects [44454647]. Gypenosides or polysaccharides from G. pentaphyllum help improve exercise tolerance, extend swimming time, prevent the rise in lactate levels, and mitigate oxidative stress during exhaustive exercise [3233]. However, the mechanisms of action of GPE and GL on muscle differentiation and mitochondrial metabolism in C2C12 myotubes remain inadequately described. The L6 and C2C12 myoblast cell lines (rat and mouse, respectively) are commonly used to evaluate myocellular metabolism and the regulatory processes involved in myogenesis. In a previous study, GPE enhanced glucose uptake, which was mediated by the AMPK and ACC signaling pathway, and also increased the gene expression of SIRT1 and PGC-1α in L6 cells [34]. In the present study, we evaluated the beneficial effects of GPE and GL, demonstrated by their association with increased myoblast differentiation, lactate metabolism, adipocyte browning, glycogen synthase, and anti-oxidative stress effects; these, in turn, are associated with the upregulation of genes and proteins related to the activated AMPK pathway in C2C12 skeletal muscle myotubes treated with the study compounds.
The compounds GL, GLI, and Rg3 are present at 1.8, 1.4, and 0.15 mg, respectively, per gram of GPE. Moreover, GL, GLI, and Rg3 are present in small quantities in raw G. pentaphyllum plants. GPE obtained by a newly patented extraction method contains large amounts of GL, GLI, and Rg3. In this study, we investigate the effects of both GPE and GL; regardless, it is also necessary to investigate the effects of GLI and Rg3 on mitochondrial metabolism. Interestingly, a study using a Caco-2 cell monolayer permeability model showed Caco-2 cell permeability of saponins from G. pentaphyllum, indicated the potential oral bioavailability of dammarane saponins from G. pentaphyllum [48]. The study indicated that GL exhibited a high permeability across Caco-2 monolayers, whereas GLI showed a 10-fold decrease in permeability, demonstrating the effects of the bioactive compound GL obtained from G. pentaphyllum. Therefore, our results suggest that GPE containing GL may have a potential for application in the production of nutraceuticals, functional foods, and food additives.
Skeletal muscle is the most plentiful tissue in the human body. It supports physiological conditions [49] and possesses a remarkable ability for regeneration following severe cell damage resulting from exercise, injury, or disease [50]. As skeletal muscle-derived stem cells, satellite cells are the basal source for myogenesis and undergo differentiation into myocytes to repair myofibers following activation by muscle injury. Satellite cells initiate myogenic differentiation and constitute their own satellite cell pool through self-renewal, which is necessary for continuous muscle regeneration [51]. In the present study, treatment of C2C12 myotubes with GPE and GL markedly promoted the differentiation and expression of Myh1 at both the mRNA and protein levels. Furthermore, the immunostaining results showed that GPE and GL treatments led to the fusion of myoblasts into multi-nucleated myotubes, as determined by changes in cell morphology and Myh1 expression. Treatment with CrM remarkably stimulated Myh7 mRNA expression levels, which were much higher than those of GPE-treated cells. However, GPE, GL, and CrM did not influence Myh2 mRNA expression in C2C12 myotubes, and GPE treatment caused a significant reduction in the Myh4 mRNA expression level. Zhang et al. [7] reported that PGC-1α transgenic mice and pigs manifested increased transcriptional induction of oxidative fiber markers, such as Myh7, Myh1, myoglobin, and TNNI1, with reduced expression levels of several glycolytic fiber genes (Myh2, Myh4, Casq1, and Tnni2). In the present study, GPE significantly augmented the Myh1 and Myh7 expression levels but not those of Myh2 and Myh4. These results showed that GPE treatment of C2C12 myotubes leads to the transformation of muscle fiber from a glycolytic type to an oxidative type. Therefore, an increased oxidative potential by treatment with GPE may help improve endurance performance and resistance to fatigue.
PGC-1α is a key transcriptional regulator of several genes involved in adjustment to exercise, such as those involved in lactate and fatty acid metabolism, antioxidant defense, muscle growth, and mitochondrial biogenesis [5525354]. In skeletal muscle, PGC-1α has also been shown to regulate muscle fiber switch, lipid oxidation, glucose uptake, and lactate catabolism. Lactate is considered an immediate energy source and an energy reserve contributor. PGC-1α controls lactate levels by promoting LDH-B conversion of lactate to pyruvate, and MCT1 expedites lactate uptake. The increase in Ldhb mRNA expression following GPE and GL treatment indicates the downregulation of lactate production when PGC-1α binds to an ERR-responsive element in the Ldhb promoter. Additionally, MCT1 expression is reportedly enhanced by PGC-1α. The activation of ERRα by PGC-1α promotes the transcription of MCT1, which directs lactate into muscle cells. In the current study, GPE and GL also upregulated Esrra and Mct1 mRNA expressions, implying that an increase in PGC-1α expression in the muscles induced by GPE and GL enhances lactate metabolism.
PGC-1α expression in skeletal muscle contributes to managing energy balance, which promotes energy expenditure. Recent studies demonstrated that PGC-1α expression in muscle induces an increase in the expression of fibronectin type III domain containing 5 (FNDC5) [15]. Irisin, one of the most important thermogenic adipomyokines secreted by FNDC5 cleavage, is involved in the browning of white fat cells and thermogenesis by activating UCP1. Irisin stimulates UCP1 expression and is positively regulated by PGC-1α [55]. Farahabadi et al. [56] reported that vitamin A increases FNDC5 gene expression via the extracellular signal-regulated kinases (ERK) 1/2-cAMP response element-binding protein (CREB)-PGC-1α-FNDC5 pathway. Chen et al. [57] showed that icariin induces irisin/FNDC5 expression in skeletal muscles via the AMPK pathway. Liu et al. [58] reported that gypenosides significantly reduce high-fat diet-induced obesity and enhance white adipose tissue browning signaling, including PGC-1α, PRDM16, and UCP1. Herein, we provided the first demonstration that GPE and GL elevate Fndc5 expression in C2C12 myotubes. After a period of exercise, AMPK is activated due to ATP consumption, which leads to acceleration in fatty acid β-oxidation, glucose uptake, and glucose phosphorylation, resulting in increased intracellular levels of glucose-6-phosphate (G6P) for ATP production [5960]. The activity of glycogen synthase (GYS), the main regulatory enzyme in glycogen synthesis, is mediated by the allosteric activator G6P. In general, AMPK negatively regulates glycogen synthesis via phosphorylation and inactivation of GYS at site 2 [59]. Thus, stimulating AMPK would lower muscle glycogen levels by inhibiting GYS enzyme activity. Recent research has shown that although 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) mediates GYS inactivation, elevated glucose transport promoted by the activation of AMPK leads to an accumulation of intracellular G6P levels. Consequently, AMPK activation enhances glycogen synthesis by activating GYS through glucose transport [6162]. We also observed that GPE and GL treatments increased the expression level of Gys mRNA in C2C12 myotubes. Furthermore, PGC-1α regulates lipid metabolism by stimulating the expression of Cpt1b target genes [63]. In the present study, we determined that GPE and GL may induce the upregulation of PGC-1α and help promote fatty acid metabolism.
PGC-1α is markedly activated by the immense energy demands of diverse physiologic and dietary conditions, and it interacts with several transcription factors. AMPK and SIRT1 signaling pathways influence PGC-1α activation directly through phosphorylation and deacetylation, respectively. In addition, p38 has an important role in activating the CREB/activating transcription factor 2 site on the PGC-1α gene promoter [3864]. PGC-1α induces the expression of Tfam and the mitochondrial regulator NRF2, which is an essential intracellular antioxidant response factor that activates mitochondrial genes in the nucleus, resulting in upregulation of mitochondrial biogenesis [65]. In addition, PGC-1α enhances mitochondrial uncoupling by inducing UCP2 and UCP3 in the inner mitochondrial membrane. UCP2 and UCP3 mediate the upregulation of energy spending, fatty acid metabolism, and thermogenic uncoupling. It has also been demonstrated that UCP2 and UCP3 contribute protective effects against mitochondrial oxidative stress damage by reducing the generation of ROS. Recent studies suggested that PGC-1α regulates the expression of endogenous antioxidant proteins. PGC-1α enhances the uncoupling capacity of the mitochondria and activates ROS-scavenging enzymes such as superoxide dismutase 2 (SOD2), catalase, and GPx1 [5366]. In the present study, we demonstrated that GPE and GL significantly increase the expression of UCP2, UCP3, SOD, and NRF2 nuclear translocation, which induces mitochondrial uncoupling and decreases ROS production. The induction of PGC-1α by GPE and GL signifies that PGC-1α activation is an instrumental factor in minimizing or preventing ROS production by upregulating the expression of antioxidant genes. Fig. 9 summarizes the proposed mechanisms through which GPE and GL improve muscle differentiation and mitochondria metabolism in C2C12 myotubes. Although GPE and GL mediate the activation of the PGC-1α signaling pathway, the effect of mitochondrial biogenesis in C2C12 myotubes remains unclear. Furthermore, whether GPE and GL could improve exercise performance through endurance training remains largely undescribed and will require further investigation.
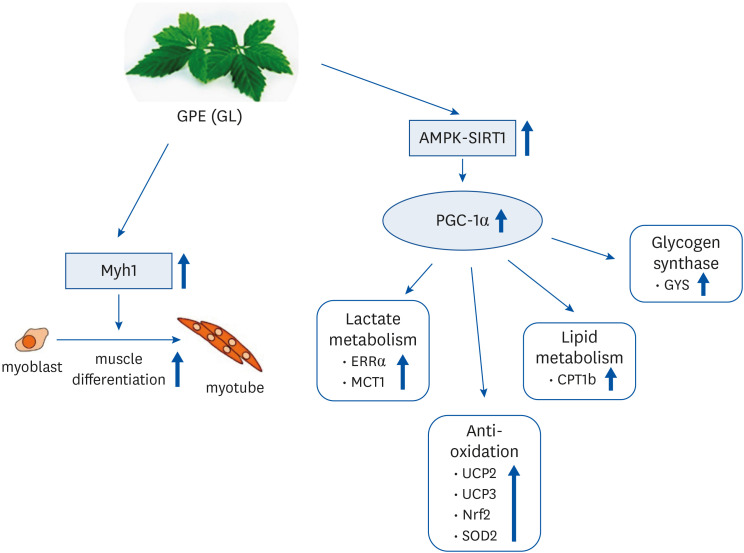
Fig. 9
Tentative mechanisms by which GPE and GL stimulate muscle differentiation and mitochondrial metabolism in C2C12 myotubes.
GPE, Gynostemma pentaphyllum extract; GL, gypenoside L; AMPK, AMP-activated protein kinase; PGC-1α, proliferator-activated receptor-gamma coactivator 1α; ERRα, estrogen-related receptor α; MCT1, monocarboxylate transporter 1; UCP, uncoupling protein; NRF, nuclear respiratory factor; SOD, superoxide dismutase; CPT, carnitine palmitoyl transferase; GYS, glycogen synthase.
![]()
The results of our investigation demonstrated that treatment with GPE and GL promoted the differentiation of myoblasts into myotubes and elevated Myh1 expression in C2C12 cells. In addition, GPE and GL stimulated lactate metabolism, upregulated PGC-1α expression by activating the AMPK/p38/SIRT1 signaling pathway, and enhanced the anti-oxidative stress effect by promoting the expression of associated genes in skeletal muscle cells. Thus, our findings underscore the potential use of GPE and GL as candidates in the further development of sports dietary supplements aimed at enhancing the health benefits of exercise. Further studies are warranted to elucidate the underlying molecular mechanism driving the effects of GPE and GL on exercise endurance capacity in animal models.
 XML Download
XML Download