

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Taste receptors are G protein-coupled receptors (GPCRs) mainly expressed in the tongue that have a role in taste recognition and taste signal transduction. The taste signaling pathway is mediated by several signaling molecules, including PLCβ2 and TRPM5 [1]. Bitter taste recognition mediated by bitter taste receptors helps the host defense against harmful compounds like toxins. There are 25 bitter taste receptor isoforms, and each receptor recognizes a different range of bitter agonists [2]. Beyond their taste-related roles, diverse functions of bitter taste receptors and downstream signals have been recently reported. Bitter taste receptors are expressed in human airways and cancer cells, which suggests they may be a therapeutic target of upper respiratory tract infection and cancer [34]. Bitter taste receptors are also located in human skin [5], yet, biological roles of these receptors are not evident.
Skin provides the first-line defense against external threats, and skin aging can deteriorate the quality of life [6]. Various factors can cause skin aging. One is diet, as high carbohydrate consumption accelerates skin aging [7]. Delayed wound healing is a major feature of skin aging. Wound healing has several steps, including hemostasis, inflammation, proliferation, and remodeling [8]. Epithelial mesenchymal transition (EMT), and re-epithelialization by migrating keratinocytes are key for wound healing. Moreover, matrix metalloproteinases (MMPs) and tissue inhibitor of metalloproteinases (TIMPs) play major roles in wound healing [910].
D-galactose (D-gal), a reducing sugar that reacts with free amines and generates advanced glycation end products (AGEs), is associated with aging [11]. AGEs can induce cellular senescence in human skin cells by targeting skin proteins, such as collagen, leading to a loss of flexibility [12]. The p53-p21 molecular pathway is involved in cell cycle modulation and is closely related to the D-gal-induced aging process. p21 is the downstream target of p53. Activated p21 induces cell cycle arrest, leading to cellular senescence. Oxidative stress occurs as AGEs generated by D-gal treatment bind with the receptor for AGEs. Antioxidant enzymes are also downregulated by D-gal treatment [1314]. The SIRT1/mechanistic target of rapamycin (mTOR) signaling pathway is reportedly related to D-gal-induced aging. Previous studies have demonstrated that D-gal administration affected the SIRT1/mTOR pathway [1516].
It has been reported that polymorphisms of the TAS2R16 and TAS2R38 genes are associated with human longevity [1718], suggesting a potential role for the bitter taste receptors encoded by these genes in the aging process. In addition, several agonists that activate bitter taste receptors have been shown to possess therapeutic properties against aging. For example, 2 TAS2R50 agonist, amarogentin exerted anti-aging and neuroprotective properties in yeast and PC12 cells, respectively [1920]. Moreover, the bitter agonist aloin alleviated oxidative stress and cognitive damage in D-gal-induced aging in mice [2122]. However, there is lack of evidence supporting the aging-related function of bitter taste receptors.
A previous study reported that TAS2R10 and TAS2R16 are found in human skin tissues [5]. In addition, certain agonists that activated these bitter taste receptors were revealed to possess anti-aging properties. The TAS2R16 agonist salicin inhibited senescence-associated β-galactosidase (SA-β-gal) activity and cell cycle arrest in senescent human umbilical vein endothelial cells [23]. Salicin also suppressed oxidative damage in a collagen-induced arthritis rat model [24]. Moreover, TAS2R10 agonist, camphor ameliorated the SA-β-gal activity in human primary dermal fibroblasts and suppressed ultraviolet (UV)-induced wrinkle in mouse skin [25]. Based on these previous studies, we hypothesized that the bitter taste receptors expressed in human keratinocytes exert anti-aging effects in D-gal-induced aging of HaCaT cells by inhibiting cellular senescence and enhancing wound healing. Biomarkers related to cell cycle arrest, antioxidant enzymes, the SIRT1/mTOR pathway, and the EMT process were examined to demonstrate the effects of bitter taste receptors in D-gal-induced aged HaCaT cells.
Go to : 
MATERIALS AND METHODS
Cell culture and transfection
HaCaT cells were purchased from the Cell Lines Service (Eppelheim, Germany) and cultured in Dulbecco's Modified Eagle's Medium (Welgene, Gyeongsan, Korea) with a 10% fetal bovine serum (Gibco, Gaithersburg, MD, USA) and 1% penicillin-streptomycin (100 U/mL and 100 µg/mL) (Invitrogen, Carlsbad, CA, USA). HaCaT cells were maintained in a 37°C, 5% CO2, humid environment. Mycoplasma infection was tested by using an e-Myco™ VALiD Mycoplasma polymerase chain reaction (PCR) detection kit (iNtRON Biotechnology, Burlington, MA, USA), and no infection was detected in the cells used. D-gal (Sigma-Aldrich, St. Louis, MO, USA) was used to induce skin aging, and stocks were prepared under dim light.
Recombinant pcDNA3 plasmids carrying human bitter taste receptor (TAS2R16, TAS2R10) genes were generously gifted by Dr. Tai Hyun Park (Seoul National University, Korea). Plasmid transfection was performed using Lipofectamine™ 3000 Transfection Reagent (Invitrogen) according to the manufacturer's instruction. pcDNA3 (empty vector) was transfected into the control (Ctrl) group.
Animals and experimental design
Male C57BL/6N mice (11-weeks-old) were purchased from Central Lab, Animal Inc (Seoul, Korea). The animals were maintained under a 12-h light and 12-h dark cycle with 22 ± 2°C temperature and 50 ± 5% humidity conditions and fed a Modified American Institute of Nutrition-93G diet (Unifaith, Inc, Seoul, Korea). After a 1-week adaptation period, animals were randomly divided into 2 groups: a Ctrl group of mice subcutaneously (s.c.) injected with a 0.9% saline solution (Ctrl, n = 6) and a D-gal group of mice s.c. injected with D-gal (500 mg/kg/day) (D-gal, n = 6). The D-gal injection was carried out every day for eight weeks. Animals were anesthetized with isoflurane (Piramal Critical Care, Bethlehem, PA, USA). After sacrifice, mouse skin tissues were collected. This animal experiment complied with and followed the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978) and performed under the approved protocol by Institutional Animal Care and Use Committee (IACUC) of Ewha (IACUC approval No. 17-019).
Western blot
We performed a Western blot as previously described [26]. We used a RIPA lysis buffer to lyse HaCaT cells and separated protein samples in sodium dodecyl sulfate polyacrylamide gels. Separated proteins were transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). The primary antibodies p21, p53, MMP-2, MMP-9, TIMP-1, TIMP-2, catalase, superoxide dismutase 1 (SOD1), SIRT1, PLCβ2 (Santa Cruz Biotechnology, Santa Cruz, Dallas, TX, USA), E-Cadherin, Vimentin, mTOR, Phospho-mTOR (Cell Signaling Technology, Danvers, MA, USA), N-Cadherin (BD Biosciences, San Jose, CA, USA), Glutathione Peroxidase I (AbFrontier, Seoul, Korea), TRPM5, β-actin (Abcam, Cambridge, MA, USA), TAS2R10, and TAS2R16 (LifeSpan BioSciences, Seattle, WA, USA) were used for detection. LumiFlash™ Ultima Chemiluminescent Substrate (Visual Protein, Taipei, Taiwan) was used to visualize the protein bands and ImageJ software (NIH, Bethesda, MD, USA) was used for quantification.
Reverse transcription PCR analysis
We used TRIzol reagent (Invitrogen) to isolate total RNA from either cells or mouse skin tissues, and generated complementary DNA (cDNA) by reverse transcription using RevertAid Reverse Transcriptase (Thermo Fisher Scientific, Waltham, MA, USA). Real-time PCR experiments were performed using a Rotor-Gene SYBR Green PCR Kit (QIAGEN, Vedbaek, Denmark). PCR analysis was done on the reverse-transcribed cDNA samples from in vivo skin tissue-isolated RNA and the cDNA samples were amplified by thermal cycling steps. The primers used were: human TAS2R10, 5′-ACG TGT AGT GGA AGG CAT CT-3′ (forward) and 5′-TGA AAT AGC TAA GCC GGT GAG-3′(reverse); human TAS2R16, 5′- GGC ATC TCT CGC TTC TGT CT-3′ (forward) and 5′-CCC AGG TGA TTG TTA AGT TGC A-3′ (reverse); human GAPDH, 5′-AGA AGG CTG GGG CTC ATT TG-3′ (forward) and 5′-AGG GGC CAT CCA CAG TCT TC-3′ (reverse); mouse Tas2r114, 5′-GCA GGC AGA TGG AAT CAA AT-3′ and 5′-ACT GCT TCA GCC GAC TGT TT-3′; mouse Tas2r118, 5′-GCC TGT CCT TTT ATT TTG GAA G-3′ and 5′-GGA AGA TGG GGT GAG TGA AG-3′; mouse Gapdh, 5′-AAC TTT GGC ATT GTG GAA GG-3′ and 5′-TGT GAG GGA GAT GCT CAG TG-3′.
Wound healing assay
HaCaT cells seeded in 6-well plates were transfected with the recombinant plasmids. When cells reached up to ≤ 90% confluence, the cell monolayer was scratched with 200 µL sterile pipette tips into 5 lines and treated with 200 mM D-gal. We pictured and measured the wound widths of each group at 0 h, 24 h, and 48 h with ImageJ software (NIH).
Senescence-associated β-galactosidase assay
A senescence β-Galactosidase Staining Kit (Cell Signaling Technology) was used to identify senescent cells. After transfection of recombinant plasmids, cells were treated with 200 mM D-gal. After PBS wash, cells were fixed and a pH-adjusted staining solution was added to each well. The plate was sealed with parafilm, covered to avoid lights, and incubated overnight at 37°C without CO2. We photographed the 5 fields of each group and quantified the stained areas of cells by ImageJ software (NIH).
Statistical analysis
Data were expressed as mean ± SE of the mean from more than 3 independent replicates. Statistical analysis was performed by 1-way analysis of variance with Newman-Keuls comparison (post hoc) or unpaired 2-tailed Student's t-test using GraphPad PRISM (GraphPad Software, SanDiego, CA, USA). A P-value < 0.05 was considered statistically significant.
Go to :
RESULTS
Expression of bitter taste receptors in HaCaT cells and mouse skin tissues
We performed PCR assays to examine the expression of bitter taste receptors in D-gal-induced aged human keratinocytes, HaCaT cells and mouse skin tissues. As shown in Fig. 1A, human bitter taste receptors, TAS2R16 and TAS2R10 were upregulated by D-gal treatment compared to the Ctrl group in HaCaT cells (P < 0.05 for both). Mouse Tas2r118 is the ortholog of human TAS2R16, and mouse Tas2r114 was identified as the ortholog of human TAS2R10 in Mouse Genome Informatics [27282930]. Subcutaneous injection of D-gal is a well-established and effective method for inducing skin-aging in animal models. Aging-related features were observed in the skin tissues of C57BL/6 mice injected with D-gal, as declined in dermal thickness and type I collagen fiber content [31]. In addition, subcutaneous injection of D-gal reduced skin elastin and moisture contents [32], damaged skin integrity and hair follicles [33], and reduced skin elasticity [34]. Consistent with HaCaT cells, the mouse bitter taste receptors, Tas2r118 (P < 0.01) and Tas2r114 (P < 0.05) were upregulated in the D-gal group in mouse skin tissues (Fig. 1B). The levels of downstream taste signaling proteins, TRPM5 and PLCβ2 were upregulated by D-gal treatment compared to the Ctrl group (P < 0.05 for both) (Fig. 1C).
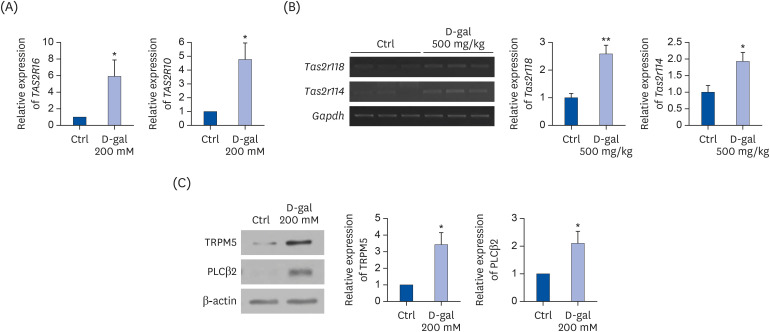 | Fig. 1Expression of bitter taste receptors in HaCaT cells and mouse skin tissues. (A) The mRNA levels of human bitter taste receptors TAS2R16 and TAS2R10 in D-gal-induced aged HaCaT cells. GAPDH was used as a loading Ctrl. (B) The mRNA levels of mouse bitter taste receptors Tas2r118 and Tas2r114 in D-gal-induced aged mouse model. Gapdh was used as a loading Ctrl. (C) Protein expression of TRPM5 and PLCβ2 in D-gal-induced aged HaCaT cells. Quantified results of TRPM5 and PLCβ2 are shown. β-actin was used as a loading Ctrl. A P-value lower than 0.05 was considered statistically significant.Ctrl, control; D-gal, D-galactose; mRNA, messenger RNA.
*P < 0.05, **P < 0.01
|
Effects of human bitter taste receptor TAS2R16 on skin aging in D-gal-induced aged HaCaT cells
TAS2R16's transfection efficiency in HaCaT cells was confirmed by Western blot assay (Fig. 2A). Although the endogenous expression of TAS2R16 was identified in HaCaT cells, its expression was upregulated by transfection. A SA-β-gal assay was performed to identify the effect of TAS2R16 overexpression on cellular senescence in D-gal-induced aged HaCaT cells. TAS2R16 overexpression reduced the levels of SA-β-gal-stained cells compared to those of the D-gal group (P < 0.05) (Fig. 2B). We investigated p53 and p21 as they are involved in the cell cycle regulation and are associated with aging [13]. Fig. 2C showed that D-gal treatment upregulated p53 and p21 expression (P < 0.05 for both), and that TAS2R16 overexpression significantly recovered this to the Ctrl level (P< 0.05 for both). In addition, D-gal treatment downregulated protein levels of antioxidant enzymes, SOD1 (P < 0.05), glutathione peroxidase 1 (P < 0.01), and catalase (P < 0.001) (Fig. 2D). However, TAS2R16 overexpression countered this suppression (P < 0.05 for all).
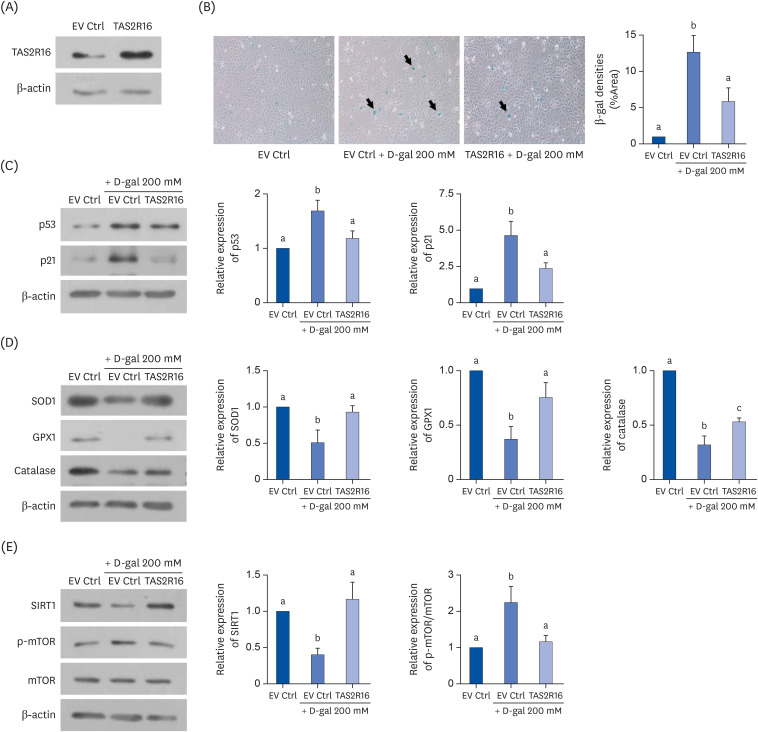 | Fig. 2Effects of TAS2R16 on aging in D-gal-induced aged HaCaT cells. (A) Transfection efficiency of TAS2R16 in HaCaT cells. β-actin was used as a loading control. (B) SA-β-gal assays to evaluate for cellular senescence. Representative photographs from each EV Ctrl, EV Ctrl + D-gal 200 mM and TAS2R16 + D-gal 200 mM group are shown in the left panel. The quantified result of SA-β-gal assay is shown in the right panel. (C) The protein levels of p53 and p21. (a) Representative bands (left panel) and quantified results of p53 and p21 (right panel) are shown. (D) The protein levels of antioxidant enzymes. Representative bands (left panel) and quantified results of SOD1, GPX1, and catalase (right panel) are shown. (E) Effects of TAS2R16 on the SIRT/mTOR pathway. Representative bands (left panel) and quantified results of SIRT1 and mTOR phosphorylation levels (right panel) are shown. β-actin was used as a loading control. The letters (a, b, and c) indicate statistical significance among the different groups.EV Ctrl, empty vector control; D-gal, D-galactose; SOD1, superoxide dismutase 1; GPX1, glutathione peroxidase 1; mTOR, mechanistic target of rapamycin; p-mTOR, phospho-mechanistic target of rapamycin; SA-β-gal, senescence-associated β-galactosidase.
|
Furthermore, the effect of TAS2R16 overexpression on a representative aging signaling, SIRT1/mTOR pathway was investigated in D-gal-induced aged HaCaT cells. Protein expression of SIRT1 was significantly downregulated (P < 0.05) and the level of mTOR phosphorylation was upregulated (P< 0.05) by D-gal treatment (Fig. 2E), but these expressions were recovered to the Ctrl level by TAS2R16 overexpression (P < 0.05 for both).
Effects of human bitter taste receptor TAS2R16 on wound healing in D-gal-induced aged HaCaT cells
The effect of overexpressed TAS2R16 on wound healing in D-gal-induced aged HaCaT cells was investigated using a scratch wound-healing assay. Relative wound widths were wider in the D-gal group both at 24 h (P < 0.001) and 48 h (P < 0.01) compared to the Ctrl group. TAS2R16 overexpression improved wound healing by narrowing the scratched wound compared to the D-gal group at both time points (P < 0.05) (Fig. 3A).
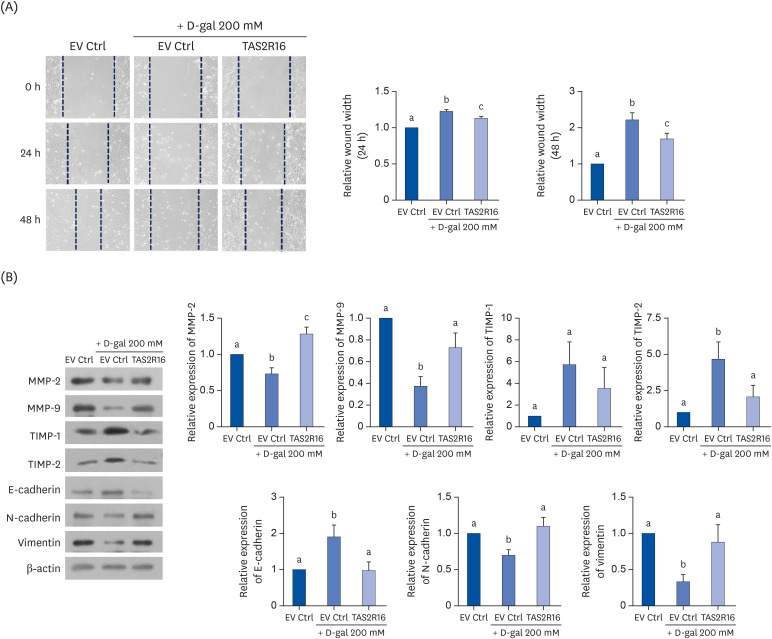 | Fig. 3Effects of TAS2R16 on wound healing in D-galactose-induced aged HaCaT cells. (A) Wound healing assays. Representative pictures are shown in the left panel. Quantified results of relative wound width at 24 h and 48 h are shown in the right panel. (B) Wound healing-related markers were examined by Western blot. Representative images (left panel) and quantified results of MMP-2, MMP-9, TIMP-1, TIMP-2, E-cadherin, N-cadherin, and vimentin (right panel). β-actin was used as a loading control. The letters (a, b, and c) stand for statistical significance among the different groups.EV Ctrl, empty vector control; MMP, matrix metalloproteinase; TIMP, tissue inhibitor of metalloproteinase.
|
EMT-related markers were analyzed as EMT contributes to the wound healing process. As shown in Fig. 3B, levels of MMP-2 (P < 0.05) and MMP-9 (P < 0.01) proteins were significantly lower after D-gal treatment compared to the Ctrl group, and TAS2R16 overexpression prevented downregulation against skin-aging (MMP-2, P < 0.01; MMP-9, P < 0.05). Expression of a metalloproteinase inhibitor, TIMP-2, was upregulated in the D-gal group (P < 0.05) compared to the Ctrl group, and their level suppressed in the TAS2R16 overexpressed group (P < 0.05). TIMP-1 showed a similar, nonsignificant trend.
E-cadherin levels were higher in the D-gal group compared to the Ctrl group (P < 0.05), but lower in the TAS2R16 overexpressed group compared to the D-gal group (P < 0.05). N-cadherin (P < 0.05) and vimentin (P < 0.05) expressions were downregulated in the D-gal group compared to the Ctrl group, but recovered to the Ctrl levels by TAS2R16 overexpression (P < 0.05 for both).
Effects of TAS2R10 on D-gal-induced aged HaCaT cells
To confirm the role of bitter taste receptors in skin aging, the bitter taste receptor, TAS2R10 was overexpressed in D-gal-induced aged HaCaT cells. We evaluated TAS2R10's transient transfection efficiency by Western blot assay (Fig. 4A). Like TAS2R16, TAS2R10 was endogenously expressed in HaCaT cells and its expression was upregulated by TAS2R10 transfection.
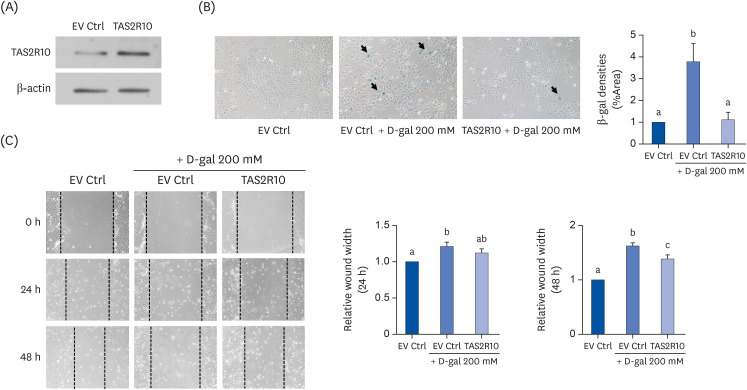 | Fig. 4Effects of TAS2R10 on D-gal-induced aged HaCaT cells. (A) Transfection efficiency of TAS2R10 in HaCaT cells. β-actin was used as a loading control. (B) SA-β-gal assays to evaluate for cellular senescence. Representative pictures from each EV Ctrl, EV Ctrl + D-gal 200 mM and TAS2R10 + D-gal 200 mM group are shown in the left panel. The quantified result of SA-β-gal assay is shown in the right panel. (C) Wound healing assays. Representative pictures are shown in the left panel. Quantified results of relative wound width at 24 h and 48 h are shown in the right panel. Letters (a, b, and c) stand for statistical significance among the different groups.EV Ctrl, empty vector control; SA-β-gal, senescence-associated β-galactosidase.
|
D-gal treatment increased the levels of SA-β-gal staining (P < 0.05), and TAS2R10 overexpression suppressed the percentage of SA-β-gal staining (P < 0.05) (Fig. 4B). In addition, wider wound widths were observed in the D-gal group both at 24 h (P < 0.05) and 48 h (P < 0.001) compared to the Ctrl group. This wound healing was increased in the TAS2R10 overexpressed group compared to the D-gal group at 48 h (P < 0.05) (Fig. 4C).
Go to :
DISCUSSION
Bitter taste receptors are generally expressed in oral tissues, mediating the taste sensation. However, this concept was overturned by recent research regarding the expression and functions of bitter taste receptors in extra-gustatory tissues. In this study, the expressions of TAS2R16 and TAS2R10 in human keratinocyte HaCaT were identified and found to be consistent with previous research on the location of these receptors in human skin tissues [5]. D-gal treatment significantly upregulated the expression of bitter taste receptors both in HaCaT cells and mouse skin tissue. Moreover, the levels of the taste signaling molecules TRPM5 and PLCβ2 increased in the D-gal group's cells. It was previously reported that LPS-treatment increased TAS2R7 and TAS2R38 levels in human lung macrophages, compared to the Ctrl group [35]. The messenger RNA expression of bitter taste receptors was also upregulated in the blood leukocytes of patients with asthma compared to healthy individuals, which suggests a protective role of upregulated bitter taste receptors against asthma [36]. In the present study, D-gal treatment induced cellular senescence and impaired wound healing activity in HaCaT cells. In addition, the expression of bitter taste receptors and their downstream molecules were upregulated by D-gal treatment. These results suggest the involvement of bitter taste receptors and their downstream signals in the regulation of cellular senescence and wound healing in skin. Previously, it was reported that the TAS2R16 agonist sinigrin accelerated wound healing in HaCaT cells. In addition, the TAS2R10 agonist caffeine ameliorated oxidative stress and apoptosis caused by UV radiation in HaCaT cells [23738]. Based on previous studies and our study results, we suggest that the upregulation of bitter taste receptors by D-gal treatment is a feedback mechanism to protect agaisnt skin aging induced by D-gal treatment. However, additional studies are required to confirm this hypothesis.
Several bitter-tasting foods possess beneficial human health properties, including managing skin aging. Bitter melon extract and citrus fruits exerted a protective effect against skin aging [3940]. The actual role of bitter taste receptors on skin aging has not been verified, however, in this present study, overexpressed TAS2R16 significantly inhibited cellular senescence by suppressing cell cycle arrest, enhancing antioxidant enzymes, and regulating the SIRT1/mTOR pathway in D-gal-induced aged HaCaT cells. TAS2R16 overexpression also reduced β-galactosidase staining. These results suggest that TAS2R16 exerts anti-aging effects on D-gal-induced aging by regulating cellular senescence in HaCaT cells. Several GPCRs besides bitter taste receptors participate in diverse biological processes. For example, administration of α2-adrenoceptor agonist downregulated reactive oxygen species (ROS) generation in adrenoceptor-expressing retinal tissues of rat [41]. Moreover, the lysophosphatidic acid receptor LPA3 suppressed ROS and prevented cellular senescence [42], suggesting that GPCRs play a role in the aging process.
Consistent with previous studies, D-gal treatment significantly hindered wound healing and inhibited EMT process in HaCaT cells [43]. We found that overexpressed TAS2R16 enhanced wound healing and EMT process. Agonists of the adenosine A2A receptor, and P2Y purinoceptor enhanced wound healing ability in in vivo murine skin tissues [4445], implying that GPCRs located in the extra-gustatory tissues have potential to accelerate wound healing.
TAS2R10 was further analyzed to clarify the role of bitter taste receptors play in skin aging. TAS2R10 was chosen as it participates in the wound healing process, as revealed by bioinformatic analysis [46]. Moreover, the TAS2R10 agonist, caffeine prevented cellular senescence in human epidermal keratinocytes [47]. We demonstrated that TAS2R10 also exerts a protective effect against D-gal induced skin aging in HaCaT cells by suppressing the SA-β-gal staining and improving wound healing.
Currently, the mechanisms by which D-gal-induced aging is impacted by bitter taste receptors are poorly understood. In intestinal epithelial cells, a subunit of sweet taste receptor, TAS1R3 knockdown alleviated ROS generation induced by the artificial sweetener aspartame [48]. A heterodimeric sweet taste receptor, TAS1R1/TAS1R3 knockdown in pancreatic β cells downregulated mTOR phosphorylation [49]. These results indicate that TAS1Rs knockdown inhibited oxidative stress and mTOR phosphorylation in extra-gustatory organs. Conversely, we found that overexpression of the bitter taste receptor, TAS2R16 enhanced antioxidant enzymes expression and downregulated mTOR phosphorylation in D-gal-induced HaCaT cells. Opposing roles of TAS1Rs and TAS2Rs in the extra-gustatory tissues were reported in previous research as well. These taste receptors in the nasal solitary chemosensory cells had contrasting roles in regulating upper-airway immunity [3]. Extra-gustatory expressing type 1 taste receptors such as sweet taste receptor might induce aging, and type 2 taste receptors, which are the bitter taste receptors, might exert the opposite effects.
In conclusion, the bitter taste receptors, TAS2R16 and TAS2R10 are protective against skin aging in D-gal-induced aged HaCaT cells. This study demonstrated the endogenous expression of bitter taste receptors in HaCaT cells and their upregulation by D-gal treatment. The anti-aging effects of bitter taste receptors are mediated by regulating cellular senescence and wound healing capabilities. As information about the role of bitter taste receptors in skin aging has been limited thus far, these findings provide evidence of a novel therapeutic role for bitter taste receptors in human skin aging and other skin disorders.
Go to :
 XML Download
XML Download