

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
A cluster of cases of pneumonia with an unknown cause in Wuhan was first reported to the World Health Organization (WHO) in December 2019; the number of confirmed cases exceeded one million in April 2020 and ten million in June 2020 [1]. According to the WHO coronavirus disease 2019 (COVID-19) dashboard, there are more than 127.8 million confirmed cases of COVID-19 infection and more than 2.7 million deaths worldwide as of March 29th 2021 with a fatality rate of 2.19%. The COVID-19 pandemic has not only affected the health of infected individuals but has also disrupted many aspects of our everyday life. People have become aware of the importance of immune function and interested in strategies to improve it. Optimal immune function is important for defenses against and recovery from viral infections. Nutritional status, immune function, and viral infection are closely interrelated. Undernutrition impairs immune function, which can lead to increased susceptibility to viral infection. Meanwhile, viral infection can result in changes in nutritional status that may further exacerbate the course of infection or delay viral clearance. Deficiency of any nutrient can cause an impaired immune response; however, several nutrients have been shown to play more significant roles than others in modulation of immune function and viral infection. Here, we review the nutrients reported to have a significant impact on immune responses, particularly the control of viral infection, and discuss the implications for COVID-19. The roles of vitamins A, C, D, and E, and zinc, iron, and selenium in immune function and viral infection and their relevance to COVID-19 will be reviewed.
Go to : 
COVID-19 AND IMMUNE RESPONSES
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which is responsible for COVID-19, is an RNA virus transmitted primarily via respiratory droplets. Among coronaviruses, SARS-CoV-2, SARS-CoV, and Middle East respiratory syndrome coronavirus replicate in the lower respiratory tract and cause substantial respiratory pathophysiology [2]. SARS-CoV-2 binds to angiotensin-converting enzyme 2 (ACE2) receptors and enters the cells in the airway of the host. Replicated viruses infect bronchial epithelial cells, type I and type II alveolar pneumocytes, and capillary endothelial cells. The proinflammatory cytokines and chemokines that are produced recruit monocytes, macrophages, and T cells to the infection site. The immune response of the host is one factor that determines the severity of COVID-19. With a healthy immune response, neutralizing antibodies inactivate viruses, macrophages clear neutralized viruses, CD8+ T cells eliminate virus-infected cells, and CD4+ T cells mediate efficient immune responses. However, when the immune response is dysfunctional, overproduction of pro-inflammatory cytokines occurs and leads to a cytokine storm and multi-organ damage [2]. Therefore, effective and controlled immune responses are critical for efficient early clearance of viruses and prevention of excessive inflammatory responses.
Although pulmonary manifestations due to aggressive inflammatory responses resulting in airway damage are the major pathophysiology of COVID-19, extrapulmonary symptoms including thrombotic complications, myocardial dysfunction, acute coronary syndrome, acute kidney injury, gastrointestinal symptoms, hepatocellular injury, hyperglycemia and ketosis, and neurologic illnesses have been reported [3].
Go to :
NUTRITIONAL CHANGES WITH COVID-19 AND VIRAL INFECTION
Changes in eating behavior and physical activity have been reported with the COVID-19 pandemic. In Poland, more eating and snacking was reported during the pandemic-associated nationwide quarantine, and 30% of the 1,097 studied subjects reported weight gain, while 18% of them reported weight loss [4]. On the other hand, the adoption of healthier dietary habits, reflected by a higher adherence to the Mediterranean Diet, in the studied population (n = 7,514) was reported in Spain during the COVID-19 confinement [5]. The purpose of the Spanish COVIDiet study was to evaluate the changes in dietary behavior during the COVID-19 confinement. Data related to the Mediterranean diet, intake of processed foods, changes in food choices, and weight gain were collected by an online questionnaire. The major difference in the surveyed individuals between the 2 studies was their age; 81.9% of the studied population was younger than 35 years in the Polish study, while only 34.0% of the studied population was younger than 35 years in the Spanish study. Changes in eating behavior affect nutrient intake, resulting in altered nutrition status.
Viral infection itself can cause alterations in the nutritional status of the host. This can be due to the changes in dietary intake related to infection and/or due to the physiological changes that occur with infection. In animals infected with influenza virus, levels of reduced glutathione and α-tocopherol were decreased in lung and liver tissues. The authors suggested that decreased antioxidant capacity could reduce the ability of tissues to protect against potential oxidative stress [6]. Among 1,020 hospitalized patients diagnosed with COVID-19 in Israel, folic acid levels were available from 333 patients, and 38 patients (11.4%) had low folate levels (< 5.9 ng/mL), but an association between low folate levels and clinical outcomes was not observed [7]. A retrospective analysis of 25-hydroxyvitamin D (25(OH)D) concentrations in plasma from patients from Switzerland showed that 25(OH)D levels were lower in patients with SARS-CoV-2 positive polymerase chain reaction results (median value, 11.1 ng/mL) than in those with negative results (24.6 ng/mL) [8]. Im et al. [9] reported that vitamin D levels were significantly lower in patients hospitalized with COVID-19 (mean value, 15.7 ng/mL, n = 50) than in a healthy control group matched by age and sex (25.0 ng/mL, n = 150). Overall, 76% of patients with COVID-19 were vitamin D deficient and 42% were selenium deficient.
Go to :
VITAMIN C
Role of vitamin C
Vitamin C, also known as ascorbic acid, plays a crucial role as an antioxidant, directly quenching free radicals and restoring other cellular antioxidants such as α-tocopherol and glutathione, thus protecting the body from oxidative damage. In addition, vitamin C is required as a cofactor for a number of reactions such as collagen hydroxylation, biosynthesis of norepinephrine and carnitine, tyrosine metabolism, and histone demethylation.
Vitamin C affects both innate and adaptive immune responses [10]. Vitamin C can strengthen the epithelial barrier function against pathogens through synthesis of collagen and protect against oxidative stress by promoting scavenging activity. In addition, vitamin C can adjust gene expression in dermal fibroblasts, enhancing their proliferation and migration, which are important for tissue remodeling and wound healing [11]. Vitamin C enhances migration of neutrophils to infection sites in response to chemoattractants, enhances phagocytosis of microbes, and stimulates the production of reactive oxygen species (ROS) and destruction of microbes. It is also needed for neutrophil apoptosis from sites of infection to protect the host tissue from potential damage. For adaptive immunity, vitamin C promotes the differentiation and proliferation of B and T cells, possibly through its gene regulating effects [10]. Similar effects on proliferation and differentiation have been observed in natural killer (NK) cells [12]. Moreover, vitamin C may help inhibit age-related impairments in immune function. High vitamin C intake has been reported to suppress age-induced thymic atrophy and maintain T cell production in mice, which is crucial for preventing the deterioration of T cell function that occurs with age [13]. Therefore, vitamin C deficiency can impair immune function and result in increased susceptibility to infections.
Effects of vitamin C on viral infection
A systematic analysis of almost 150 animal studies on pure vitamin C and infections showed that vitamin C may mitigate or prevent bacterial or viral infections [14]. These protective effects include increased resistance of chick embryos and tracheal organs to avian coronavirus after ingestion of vitamin C. High plasma concentrations of vitamin C have been shown to be associated with enhanced immune function, such as antibody responses, neutrophil function, and antiviral activity in animal studies [10]. In a study of mice exposed to influenza A virus (H3N2), vitamin C was shown to be an essential factor for antiviral immune response by producing interferon (IFN)-α/β during the early stages of infection [15].
Despite the apparent effects of vitamin C against infections in diverse animal species, the effects of vitamin C on susceptibility to viral infections in humans are conflicting due to various study designs and dosages used. The most extensively studied human infection is the common cold, which is caused by a respiratory virus in most cases. A Cochrane review analyzing 29 randomized controlled trials (RCTs) found that regular supplementation with vitamin C (at least 200 mg) did not decrease the incidence of colds in the general population, but supplementation with 1 to 2 g of vitamin C per day had a consistent effect on the duration and severity of colds without adverse effects [16]. In addition, the study concluded that high-dose supplementation is not necessary for general community but may be reasonable during periods of heavy physical stress. A more recent meta-analysis of 9 RCTs found that the combination of extra doses of vitamin C (0.7 to 8 g/day) and a daily supplemental dose (no more than 1 g/day) reduced the duration of the common cold and relieved symptoms [17].
The host response to viral infections include the generation of ROS from activated phagocytes. ROS play a role in the process of killing pathogens. However, ROS can damage the host cells, and in some cases, they are implicated in the pathogenesis of infections. The oxidative stress caused by increased ROS during the immune response is related to the induction of inflammatory response genes, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1, IL-8, and intercellular adhesion molecule 1 (ICAM-1), through activation of the nuclear transcription factor nuclear factor κB (NF-κB) [18]. Excessive increases in inflammatory cytokines and chemokines at once, which is called a cytokine storm, is a cause of severity in some conditions, such as sepsis and acute respiratory distress syndrome (ARDS). Excessive generation of cytokines increases neutrophil infiltration, leading to neutrophil extracellular traps (NETosis). NETosis is a cell death pathway that leads to tissue injury, organ damage, and the development of multi-organ failure. Evidence suggests that vitamin C can regulate this process. Vitamin C infusion has been shown to attenuate NETosis induced by sepsis in vitamin C-deficient Gulo−/− mice model [19]. Moreover, vitamin C supplementation decreased serum levels of TNF-α and IL-6 and enhanced superoxide dismutase, catalase, and glutathione levels in a rat ARDS model [20].
During various infections, vitamin C levels decreased, and the requirement for vitamin C increased with the severity of the infection [14]. In severe cases, high-dose intravenous vitamin C may be required to achieve high enough levels to compensate for the turnover of the vitamin in the body. A meta-analysis of intravenous vitamin C supplementation (200 mg/kg body weight/day) in the critically ill patients found that it shortened the intensive care unit (ICU) stay and decreased the need for mechanical ventilation [21]. In addition, an RCT involving patients with sepsis-associated ARDS reported that administration of ~15 g/day of intravenous vitamin C for 4 days decreased 28-day all-cause mortality; however, it did not improve organ function scores or inflammatory markers [22].
Relevance to COVID-19
Many severely ill patients with COVID-19 have been reported to have elevated cytokine levels [23]. A cytokine storm can damage the lung tissue by uncontrolled inflammation and is regarded as a key factor in the progression to ARDS. Considering the beneficial effects of large doses of vitamin C in sepsis and ARDS, high-dose vitamin C may have beneficial applicability in patients with COVID-19. More than 20 clinical trials studying the therapeutic effects of vitamin C alone or in combination with other substances are registered on clinicaltrials.gov [24]. Results from these ongoing or just completed clinical trials will help to produce definitive evidence in the near future. A recent review proposed that much higher doses of intravenous vitamin C may be required to reduce the cytokine storm in ARDS that occurs during the late stage of COVID-19 [25].
Given the potential role of vitamin C as an antioxidant and immune enhancer in various infectious conditions and its lack of associated serious adverse effects, an oral low dose (1–2 g/day) may be effective for prophylactic use.
Go to :
VITAMIN D
Role of vitamin D
Vitamin D is a fat-soluble vitamin that can be taken from the food or supplements and is the secosterol prohormone that can be synthesized by skin 7-dehydrocholesterol via UVB exposure. Absorbed vitamin D3 (cholecalciferol) is bound to vitamin D-binding protein and then transported to the liver and hydroxylated by 25-hydroxyase to 25(OH)D, the major circulating form of vitamin D. Serum 25(OH)D concentration is a marker of vitamin D status, and it reflects the amount of vitamin D synthesized in the skin and consumed from the diet. Then, 25(OH)D is converted to 1,25-dihydroxyvitamin D (1,25(OH)2D) via the action of 1-hydroxylase (CYP27B1), which is mainly expressed in kidney. 1,25(OH)2D exerts its effects by binding to the vitamin D receptor (VDR) and modulating the transcription of target genes with the vitamin D response element (VDRE) in their promoters [26]. Upon discovery of CYP27B1 and VDR expression in immune cells, including macrophages, T cells and B cells, the role of vitamin D in immunity has been highlighted [2728]. Overall, vitamin D has been reported to have inhibitory effects on the adaptive immune system by inhibiting Th1 and Th17 responses and decreasing the proliferation of T and B cells; in contrast, it has relatively stimulatory effects on innate immune cells specifically monocytes and macrophages. Vitamin D has been reported to suppress the maturation of dendritic cells (DCs). The effects of vitamin D on NK cell activity remain unclear because enhanced lytic activity of NK cells, inhibited NK cytotoxicity, or no changes in NK cytotoxicity have been reported with in vitro vitamin D treatment. In particular, vitamin D induces monocyte proliferation and regulates the expression of antimicrobial peptides (AMPs), such as cathelicidin (LL37), beta-defensin2 (DEFB4), and IL-1, thereby contributing to innate immune responses in respiratory tract infections (RTIs) [29].
Effects of vitamin D on viral infection
The complement system and antibacterial responses by neutrophils and macrophages are essential parts of immune responses to viral infection. Monocytes and macrophages are involved in both pathogen phagocytosis with AMPs and pathogen-associated molecular pattern recognition by pattern recognition receptors [30]. AMPs, also known as host defense peptides, function as an important first line of defense against bacterial, viral, or fungal infections of eukaryotic organisms by killing microorganisms directly or by affecting other innate immune cells. 1,25(OH)2D and VDR complex directly induce the antibiotic protein LL37 in monocytes and neutrophils by binding to the VDRE in LL37 [3132]. Peripheral blood monocyte LL37 expression has been reported to be positively correlated with variations in serum 25(OH)D levels and significantly enhanced by exogenous 25(OH)D treatment [32]. Human DEFB4, which is expressed in monocytes, macrophages, and DCs, also contains a VDRE in the proximal promoter site [31]. Human DEFB4 is modestly induced by 1,25(OH)2D and enhances protection against viral infection [3133].
Many epidemiological studies have reported a close relationship between vitamin D deficiency and RTIs. An association between serum 25(OH)D concentration lower than 16 ng/mL at baseline and the risk of acute RTI in young Finnish men and significant correlations between increased numbers of RTIs and low 25(OH)D level in Bangladeshi and Indian children have been observed [343536]. A retrospective analysis of the Third National Health and Nutrition Examination Survey of 18,883 patients showed an increased prevalence of upper RTIs in people with serum 25(OH)D levels less than 30 ng/mL (75 nmol/L) when compared with a subgroup of individuals with 25(OH)D levels higher than 30 ng/mL [37]. Furthermore, according to a meta-analysis that assessed the preventive effect of vitamin D intake on RTIs, vitamin D supplementation showed a protective effect against RTIs and this effect was stronger with daily doses than with a bolus dose [38]. Although several studies have reported contradictory results showing no significant difference in 25(OH)D levels between patients with RTIs and control subjects [3539], vitamin D may have a beneficial role in the respiratory tract in response to viruses by induction of AMPs.
Relevance to COVID-19
Vitamin D is one nutrient that has been investigated the most in relation to COVID-19. A study evaluating the correlation of vitamin D levels with COVID-19 cases in UK Biobank (502,624 participants) showed that circulating vitamin D levels were not significantly correlated to COVID-19 infection [40]. On the other hand, a negative correlation between vitamin D levels and COVID-19 cases [41] and negative correlations between the mean vitamin D levels and the number of COVID-19 cases/1 M population were observed in 20 European countries [42]. Kaufman et al. [43] reported that there is a strong negative correlation between circulating 25(OH)D levels and SARS-CoV-2 positivity, regardless of latitude, ethnicity, sex, and age. A total of 191,779 patients from all 50 states with SARS-CoV-2 results and matching 25(OH)D results from the preceding 12 months were included in the study. Patients with vitamin D deficiency (25(OH)D < 20 ng/mL) showed a significantly higher SARS-CoV-2 positivity rate than patients in subgroups with higher vitamin D levels (30–34 ng/mL and > 55 ng/mL).
In addition, a recent study demonstrated a protective effect of 1,25(OH)2D against SARS-CoV-2 in human nasal epithelial cells (with a reduction of 0.69 log10 in viral titers) [44]. The authors screened 4 compound libraries: a United States Food and Drug Administration (FDA)-approved drug library, an ACE2-targeted compound library, a flavonoid compound library, and a natural product library. Among them, only 1,25(OH)2D mediated inhibition of SARS-CoV-2.
In conclusion, vitamin D status is closely associated with SARS-CoV2 infection, and vitamin D supplementation may have a protective role against SARS-CoV2 infection; however, there is not enough evidence of vitamin D's effect on COVID-19 severity. To prevent vitamin D deficient or insufficient states, one of the potential risk factors for COVID-19, daily supplementation with moderate doses of vitamin D3 could be recommended. However, vitamin D supplementation in patients with severe SARS-CoV2 should be cautiously approached because vitamin D can inhibit adaptive immune cell responses such as B and T cell proliferation.
Go to :
VITAMIN A
Role of vitamin A
Vitamin A is a fat-soluble vitamin that plays an important role in maintaining vision, promoting growth and development, and protecting the epithelium and mucus integrity in the body. Vitamin A exists as retinol, retinal, and retinoic acid, and the most biological activity is shown with retinoic acid. Retinoid acid regulates the differentiation, maturation, and function of cells of the innate immune system including CD169+ macrophages, neutrophils, and NK cells and plays a role in transformation of M1 macrophages into M2 macrophages in the bone marrow [45]. Retinoic acid enhances cytotoxicity and T cell proliferation in part by enhancing IL-2 secretion and promotes Th2 differentiation by inducing Il4 gene expression and Th2-cell promoting transcription factors, such as GATA3. Retinoic acid blocks the differentiation of Th17 cells and enhances the induction of regulatory T cells (Treg cells) [29]. Retinoic acid promotes immunoglobulin A secretion and upregulates gut-homing receptors [2945]; therefore, vitamin A is especially important for gut immunity and integrity, which explains why vitamin A deficiency puts children at risk for diarrhea.
Effects of vitamin A on viral infection
When the effects of vitamin A supplementation for preventing morbidity and mortality in children aged six months to five years old were assessed based on the results from 47 RCTs and cluster RCTs, nine trials reported mortality due to diarrhea and showed a 12% overall reduction with vitamin A supplementation (risk ratio [RR], 0.88; 95% confidence interval [CI] 0.79–0.98). There was no significant effect of vitamin A supplementation on mortality due to measles, respiratory disease, or meningitis. Vitamin A supplementation reduced the incidence of diarrhea (RR, 0.85; 95% CI, 0.82–0.87; 15 studies) and measles (RR, 0.50; 95% CI, 0.37–0.67; 6 studies) but not the incidence of respiratory disease [46].
Vitamin A deficiency resulted in excessive inflammation, more severe liver pathology, and increased death in a mouse model of persistent lymphocytic choriomeningitis virus infection. When retinoic acid was supplemented, the hyperactive T-cell response was downregulated. Retinoic acid may modulate T-cell activation by regulating T cell receptor signals and nuclear factor or activated T cells transcriptional factors [47]. Vitamin A deficiency is also associated with dysregulated immune responses to influenza virus and increased mortality rates after bacterial coinfections. Vitamin A-deficient mice failed to recruit CD4+ T cells to the lungs and their viral titers were higher than the vitamin A-sufficient mice and viral clearance was delayed [48].
Relevance to COVID-19
Changes in vitamin A status with COVID-19 are not available to date. Stephensen and Lietz [49] reviewed the relevance of vitamin A to SARS-CoV-2 infection. The authors suggested that vitamin A may play a role in COVID-19 by maintaining innate and adaptive immunity to promote clearance of primary infection and to reduce the risk of secondary infections. In addition, it may play a role in the respiratory tract by minimizing inflammation, supporting repair of the respiratory epithelium, and preventing fibrosis. Vitamin A deficiency can develop during COVID-19 due to specific effects on lung and liver stores caused by inflammation and impaired kidney function, which may necessitate vitamin A supplementation to restore its status. However, vitamin A status and the levels and duration of vitamin A supplementation should be carefully considered because excessive vitamin A intake is associated with toxicity and because vitamin A storage in the body varies among individuals. Furthermore, vitamin A supplementation may restrain the adverse effects of SARS-CoV2 on the angiotensin system as well as counterbalance the adverse effects of COVID-19 therapies. However, evidence specific to COVID-19 is needed to support these suggestions.
Considering the specific effects of vitamin A on the immune system and viral infections, benefits of vitamin A supplementation for COVID-19 are possible. However, high doses of vitamin A as retinol (exceeding 20,000 IU/day over short periods) can lead to intoxication. Furthermore, inflammation affects retinoid metabolism, and sequestration of retinol in tissues may result in decreased serum retinol levels [45]. Therefore, the use of serum retinol during inflammation to assess the vitamin A status may be problematic, and vitamin A status in individuals with COVID-19 should be interpreted in the context of each individual's inflammation status.
Go to :
VITAMIN E
Role of vitamin E
Vitamin E is a fat-soluble vitamin that functions as an antioxidant protecting polyunsaturated fatty acids in the membrane from oxidation. Vitamin E inhibits protein kinase C (PKC) activity and may directly affect enzymes involved in the generation of lipid mediators [50].
The immunomodulatory effects of vitamin E are evident in aged animals and humans experiencing age-associated decline of immune function. Increased lymphocyte proliferation, immunoglobulin levels, antibody responses, NK cell activity, and IL-2 production and decreased IL-6 and prostaglandin E2 production have been reported with vitamin E supplementation [5051]. In vitro treatment with vitamin E enhanced the cell division and IL-2 producing capacity of naïve T cells, increased the percentage of CD4+ T cells capable of forming an effective immune synapse, and reversed age-associated defects in the phosphorylation of linker for activation of T cells in CD4+ T cells from old animals [5253].
Effects of vitamin E on viral infection
Vitamin E deficiency has been shown to result in increased viral titers and IL-1β, TNF-α, and RANTES in the brains of mice intranasally inoculated with herpes simplex virus-1. However, no difference was observed between mice fed the supplemented level (384 mg/kg diet) of vitamin E and those fed an adequate level (38.4 mg/kg diet). Vitamin E supplementation (500 mg/kg diet) was shown to provide enhanced resistance to influenza virus infection as evidenced by decreased viral titers. Modest improvement in NK cell activity and increased Th1 cytokine (IL-2 and IFN-γ) production seemed to contribute to viral clearance as an inverse correlation was observed between IFN-γ levels and viral titers [5054]. Vitamin E alleviated the excessive production of TNF-α in old mice [54].
In humans, the effects of vitamin E on resistance to infection cannot be determined by induced infections; therefore, the natural incidence of infectious diseases among vitamin E supplemented population has been determined in several studies. The incidence of the common cold or pneumonia in male smokers, respiratory infections in nursing home residents or non-institutionalized individuals, and malaria in human immunodeficiency virus (HIV)-infected pregnant women has been investigated. The magnitudes of the effects of vitamin E on decreasing incidence of infectious diseases were rather small, and in some studies, positive effects were only observed in subgroups of subjects or no beneficial effect was observed. These discrepancies among studies can be attributed to the differences in supplemental doses, single nutrient vs. combination of nutrients, and the age, sex, and nutritional status of the subjects [5051].
Relevance to COVID-19
In a prospective case-control study, vitamin E levels were compared between 60 pregnant women with COVID-19 infections and 36 age-matched pregnant women without COVID-19 infections. Serum vitamin E levels were significantly lower in the COVID-19 group (median value, 2.57 μg/mL) than in the control group (6.73 μg/mL). Pregnant women with COVID-19 had lower neutrophil and lymphocyte counts, and their levels of C-reactive protein (CRP) and ferritin were higher than pregnant women without COVID-19, indicating increased oxidative stress [55].
Considering the antioxidant, anti-inflammatory, and immunomodulatory functions of vitamin E, preserving vitamin E levels by supplementation may confer a beneficial effect in individuals with COVID-19. The enhanced T-cell function and reduced production of inflammatory cytokines achieved by vitamin E may potentially reduce the severity of the disease.
Go to :
ZINC
Role of zinc
Zinc is an essential trace element involved in many biological processes such as activation of more than 300 enzymes and expression of 2,000 transcription factors [56]. Nearly 2 billion individuals in the developing world are estimated to be affected by a nutritional zinc deficiency. Zinc deficiency is related not only to inadequate intake but also to malabsorption of zinc from the diet. Nutritional zinc deficiency is caused by a phytate, a potent inhibitor of zinc absorption, which is present in staple foods such as unrefined cereal in developing countries [57]. Since zinc plays critical roles in multiple aspects of the immune system, zinc deficiency affects the development and function of immune cells, including neutrophils, NK cells, and macrophages, and negatively affects the proliferation and function of T and B cells [58]. According to the World Health Report by the WHO [59], zinc deficiency is responsible for approximately 16% of lower RTIs.
Effects of zinc on viral infection
Zinc can affect viral infections including common flu caused by rhino- and coronaviruses. A systematic review of RCTs reported that adults and children receiving zinc had a shorter duration of cold symptom [60]. Furthermore, zinc administration within 24 h after the onset of common flu symptoms reduced the duration of the flu [61]. According to a review by Overbeck et al. [62], these beneficial effects of zinc on viral infection are related to inhibition of viral replication and modulation of immune cells by reducing cytokine production. First, binding of positively charged zinc ions (Zn2+) with negatively charged regions of rhinovirus can inhibit viral binding of ICAM-1. Second, IFN-α production induced by zinc can contribute to its antiviral action. In addition, the inhibitory effect of zinc on prostaglandin metabolism has been suggested as a potential mechanism of action of zinc against the common cold; however, the authors concluded that more studies are needed to explain how in vivo zinc treatment works for the common flu. The level of Zn2+ in cells is tightly controlled, by binding to cellular proteins, in particular to metallothioneins, and by involving zinc transporters such as SLC39 and SLC30 [63]. However, some compounds such as pyrrolidinedithiocarbamate or pyrithione (PT), which stimulate cellular import of zinc, can induce high intracellular Zn2+ levels; thereby, zinc can inhibit the replication of influenza and RNA rhino viruses [6465].
Zn2+ has been also demonstrated to inhibit the RNA-synthesizing activity of the replication and transcription complex of SARS-coronavirus in Vero-E6 cells when treated with PT. Specifically, RNA polymerase elongation and template binding are blocked by Zn2+ [66]. Furthermore, Zn2+ uptake into cells has been shown to be enhanced by chloroquine, which has emerged as a potential candidate treatment for COVID-19; furthermore, increased intracellular Zn2+ has been shown to increase the cytotoxicity and mediate the antiviral effect of chloroquine against COVID-19 [67].
Relevance to COVID-19
Indirect evidence indicates a role of Zn2+ against COVID-19 infection. According to the review by Wessels et al. [68], zinc is closely associated with the activity of ACE2, which is known to be the receptor SARS-CoV-2 utilizes for entry into target cells, since ACE2 is a zinc-metalloenzyme that requires zinc for its enzymatic activity. The authors [68] suggested that zinc may decrease ACE2 expression through downregulation of Sirt-1 activity, thereby reducing viral entry into cells. In addition, zinc deficiency is associated with the production of pro-inflammatory cytokines and ROS in the lungs, which is one of the characteristics of COVID-19, and this inflammatory response is reversible by zinc supplementation [69]. Zinc modulates immunity through inhibition of NF-κB, a cytoplasmic transcription factor, is critical for the expression of many pro-inflammatory cytokines, and is required for T-cell proliferation, the balance between CD4+ and CD8+ T cells, and the secretion of T-cell dependent ILs and cytokines including IL-1, IL-2, IL-4, and IFN-γ [70]. Therefore, the immunomodulatory effects of zinc may be beneficial for the prevention and treatment of COVID-19, including limiting the cytokine storm associated with COVID-19.
Few epidemiology studies have assessed zinc status in patients with COVID-19. According to one recent observational study involving 47 Indian patients with COVID-19, patients with COVID-19 showed low zinc levels compared to normal healthy people, and zinc-deficient COVID-19-positive patients had increased complication rates (P=0.009), increased rates of ARDS, received corticosteroid therapy, prolonged hospital stays, and increased mortality [71]. Another study of supplementation of 440 mg of zinc sulfate in patients with COVID-19, found no significant casual association between zinc and the risk of in-hospital mortality; however, the authors concluded that their results must not be used to confirm the clinical benefits of zinc for COVID-19 management because of the limitations of their study design (short period of observation) [72].
In conclusion, zinc can contribute to mitigating viral infections of the respiratory tract and might have beneficial effects for the prevention and treatment of COVID-19 infection. A list of zinc supplementation studies in respiratory infections depending on the zinc compound and intake duration has been published by Wessels et al. [68]. Zinc supplementation for preventing respiratory infection was more effective than therapeutic procedures. Therefore, it is recommended for vulnerable people to Covid-19 infection to take zinc supplementation. However, large dose of zinc can cause the suppression of immune system and interfere with absorption of other minerals. The tolerable upper intake level established by the Food and Nutrition Board of the US institute of Medicine is 40 mg a day and tolerable upper intake level from dietary reference intakes (DRIs) for Koreans is 35 mg/day.
Go to :
IRON
Effects of iron on immune function and viral infection
Iron is an essential trace element that functions in diverse metabolic processes including oxygen transport, electron transport chain, and DNA synthesis. Iron deficiency is associated with suboptimal health including impairment of immune function, leading to an increased risk of infections [73]. Considering that ribonucleotide reductase, which is involved in DNA synthesis, requires iron, iron deficiency can lead to decreased lymphocyte proliferation [74]. In addition, iron modulates lymphocyte function by regulating cytokine production and PKC activation. Moreover, iron is required for myeloperoxidase activity, which is involved in the bactericidal process through the formation of highly toxic hydroxyl radicals. Prolonged iron deficiency has been shown to reduce antibody production in mice exposed to influenza virus [75].
However, therapeutic doses of oral iron supplementation have been linked to increased rates of malaria and increased morbidities with other infectious diseases [73]. These detrimental effects may be because supplementation provides iron-requiring microorganisms with a suitable environment to grow and replicate. Therefore, both iron deficiency and overload above a threshold level are associated with unfavorable immune responses.
Relevance to COVID-19
In a retrospective study involving 50 hospitalized patients with COVID-19, those with severe COVID-19 had higher hepcidin and serum ferritin levels than those with mild diseases and healthy people [76]. Hyper-ferritinemia has been associated with increased mortality risk [23]. The serum ferritin levels in COVID-19 non-survivors were two-fold those in survivors. Ferritin is a major intracellular iron storage protein in its ferric state (Fe3+). Hepcidin is a peptide hormone secreted by the liver and plays a role in the regulation of iron metabolism. It controls serum iron by binding to ferroportin, an iron transporter in iron-absorptive enterocytes and iron-recycling macrophages. Binding of hepcidin causes internalization and degradation of ferroportin, which prevents entry of iron into plasma [77].
During inflammation, cytokines, particularly IL-6, increase ferritin and hepcidin levels [78]. This response is important for immune function to limit the available iron for proliferating pathogens and for preventing excessive oxidative damage. Evidence suggests that a ‘cytokine storm’ is a hallmark of the hyper-inflammatory state in patients with COVID-19 and contributes to deterioration of the patients [23]. Decreased iron influx into plasma results in low transferrin saturation, and less iron is delivered to the developing erythroblasts, leading to anemia of inflammation. A modest decrease in hemoglobin levels has been observed in patients with COVID-19 [79].
The increased serum ferritin levels in patients with severe COVID-19 are strongly associated with hyper-inflammation. In addition, it is possible that dysregulated iron status plays a role in the pathogenesis of COVID-19. Excess intracellular iron due to increased ferritin and hepcidin can contribute to oxidative damage of cellular components in various organs and lead to ferroptosis, a type of programmed cell death mediated by iron-dependent peroxidation. Ferroptosis has been found to be associated with neurological disorders, agueusia and anosmia (taste and smell loss), which are manifested in patients with COVID-19 [80]. Iron overload can also increase blood coagulation by interacting with proteins of the coagulation cascade, which is a major concern in the pathogenesis of COVID-19. Therefore, altered iron homeostasis in association with hyper-inflammatory status may be a key modulator of the pathogenesis of COVID-19.
Recently, iron chelators have been proposed as an adjunctive treatment to improve clinical outcomes of patients with COVID-19 [81] based on the understanding that iron chelation can inhibit viral replication and endothelial inflammation by decreasing iron availability. However, there is a concern about the application of iron chelators. Withholding iron may be counterproductive and should not be employed until there is evidence that elevated iron levels exist and have relevance in COVID-19 [82]. There are few studies in this regard. In a study with 50 hospitalized patients with COVID-19, serum iron deficiency was observed in these patients, and the severity of COVID-19 was correlated with low serum iron levels [83]. Other therapeutic approaches may include ferroptosis inhibitors, hepcidin modulators, and erythropoietin. However, large RCTs and data analyses are needed to elucidate the efficacy and safety of therapeutic treatments for COVID-19. It is important to maintain an optimal iron level for immune function and to mitigate risk factors associated with COVID19 infection.
Go to :
Selenium
Role of selenium
Selenium is an essential trace element that has a variety of physiological effects, including antioxidant and immune-modulating properties [84]. Dietary selenium is incorporated into selenoproteins during their synthesis in the form of selenocysteine. Selenocysteine is an analogue of cysteine in which sulfur is substituted with selenium. The biological function of selenium is performed primarily through selenoproteins. To date, 25 selenoproteins have been identified in humans, including enzymes that play crucial roles in oxidative stress response such as glutathione peroxidases (GPXs) and thioredoxin reductases (TXNRDs). GPXs have important roles in protecting against ROS damage, and TXNRDs participate in a redox homeostasis and signaling. In addition, a series of selenoproteins are present in the endoplasmic reticulum where they function in protein folding and activation of various signaling pathways [8485].
Effects of selenium on immune function and viral infection
Selenium deficiency contributes to poor immune function, increased susceptibility to infection, and increased risk of mortality [84]. Selenium influences both the innate and adaptive immune systems [86]. During infection, excessive ROS production by immune cells, especially macrophages, at the site of infection triggers oxidative damage to the tissues and initiates signaling cascades that lead to inflammatory responses. Selenium can contribute to epithelial barrier maintenance by protecting against ROS through the antioxidant function of selenoproteins such as GPXs. In addition, T-cell proliferation and NK cell activity are impaired, and the production of immunoglobulins is decreased in selenium deficiency [87]. A prospective study involving 83 patients with respiratory disease found that serum selenium levels were 28% lower in the ICU group than in the general ward group [88]. Low serum selenium status has been shown to be associated with decreased lymphocyte counts and albumin concentrations as well as increased CRP, which is a marker of inflammation. In addition, selenium deficiency can increase the virulence of viruses. The first evidence came from the study of Keshan disease, an endemic cardiomyopathy in the Keshan area in China that leads to congestive heart failure and death. The disease is caused by a combination of selenium deficiency and infection with coxsackievirus B3 (CVB), a single-stranded RNA virus [89]. Sequencing of the viral genome revealed that CVB and influenza viruses mutated into more virulent forms in selenium-deficient mice that were unable to produce adequate GPX [90]. Selenium deficiency has been linked to increased mortality in patients infected with some other RNA viruses, such as HIV-1, hepatitis, and hantavirus [91]. Therefore, low selenium status not only affects the host response to infection but also affects the viral mutations and pathogenicity.
These results suggest that selenium supplementation improves immune responses. In influenza virus-infected mice, selenium supplementation reduced mortality and improved the response to infection, by stimulating the production of TNF-α and IFN-γ [92]. Selenium supplementation (50-100 μg/day of selenium selenite) for 15 weeks in adults with marginally low selenium status (< 95 ng/mL) increased plasma selenium concentrations and enhanced cellular immune responses to a live polio vaccine, leading to more rapid clearance of the virus with earlier peak T-cell proliferation and increased numbers of T-helper cells [93]. Similarly, a 12-week RCT involving healthy adults with a marginal selenium status (< 110 ng/mL) supplemented with selenium (50, 100 μg/day of selenomethionine in a yeast matrix) after flu vaccination resulted in beneficial effects for cellular immunity, such as increased T-cell proliferation, IL-8, and IL-10; however, humoral immune responses were unaffected. However, higher doses of supplementation (200 μg/day of selenomethionine in a yeast matrix) resulted in reduced granzyme B content of CD8 cells, suggesting a detrimental effect of high supplemental doses [94]. Therefore, optimizing plasma selenium may be required to improve immune responses.
Relevance to COVID-19
Given the evidence that selenium or selenoproteins protect against viral infection and support immune responses, selenium status could influence COVID-19 infection and disease outcomes. A recent ecological analysis in China indicated that COVID-19 cure rates in different regions were positively associated with the mean regional selenium status as previously measured by hair selenium concentrations [95]. In addition, a German study found markedly low levels of serum selenium and selenoprotein P in patients with COVID-19 compared with reference data from the European EPIC cross-sectional study [96]. Moreover, selenium status was higher in COVID-19 survivors than in non-survivors.
The deterioration of patients with COVID-19 has been attributed to a cytokine storm, which involves an excessive release of pro-inflammatory cytokines [23]. This cytokine storm can be caused by unregulated ROS production through NF-κB signaling [97]. A recent murine study found that selenium deficiency promoted NFκB-mediated inflammation and inhibited macrophage phagocytosis [98]. Based on these results, selenium supplementation might be beneficial for patients with COVID-19 with moderate to severe symptoms. In a recent RCT study, critically ill patients with COVID-19 and ARDS were administered intravenous selenium (1 mg for 3 days and 1 mg/day for 6 successive days as sodium selenite). Serum selenium concentrations were associated with serum GPXs and inversely related to serum levels of IL-1ß and IL-6, indicating that selenium restored antioxidant activity and moderated inflammatory responses [99]. However, there were no apparent effects on survival or the duration of mechanical ventilation or ICU stay in that study. Further studies are needed to elucidate the effect of selenium supplementation on NF-κB expression and survival benefit in patients with COVID-19. Nevertheless, selenium status may be a determinant of COVID-19 disease outcome, and selenium supplementation could be beneficial for patients with COVID19 with a sub optimal or low selenium status.
Selenium is known to have a narrow range of beneficial and adverse effects. In general, selenium supplementation is not recommended for those with an adequate selenium status to avoid selenium toxicity due to high selenium intake. Selenium requirements are based on intake that maximizes activities of the plasma selenoproteins, which are the criteria used in the DRIs of many countries. However, the cure rate of patients with COVID-19 continued to increase beyond the intake of selenium required to optimize selenoprotein activity [95]. In addition, daily doses of 1 mg of intravenous selenium (as selenite) have been used in critically ill patients with sepsis or ARDS without causing adverse effects [99].
In conclusion, dietary selenium supplementation (50–100 μg/day) may be beneficial as an adjuvant therapy for viral infections, in particular for those at risk of selenium deficiency. Supranutritional doses of selenium might be beneficial in patients with COVID-19 with moderate to severe symptoms.
Go to :
CONCLUSION
Decreased levels of vitamin D, folate, vitamin E, zinc, iron, and selenium have been reported in different populations of patients with COVID-19 compared with healthy individuals. Identifying the nutritional status affected by viral infection and restoring these decreased levels are important since deficiencies of vitamins A, D, E, and C, zinc, iron and selenium have been reported to impair immune function and increase the risk of infectious diseases. However, supplementations with these nutrients should be approached cautiously considering other physiological effects, toxicity with high doses, and interactions with other nutrients. Even without toxicities, high levels of these nutrients may adversely affect immune function. Vitamins A, D, E, and C, zinc, iron and selenium have been shown to modulate different arms of the immune system in animals and humans, and evidence suggests the potential benefits of their use for COVID-19 (summarized in Table 1); however, clinical research data are needed to support the therapeutic use of these nutrients. Currently many clinical trials designed to investigate the effects of vitamins D, C, A, and B12, and zinc as single components or in combination in relation to COVID-19 are registered in the clinical trial registry of the National Institutes of Health (https://clinicaltrials.gov/) [100]. Findings from these studies will provide the insight into efficacy of supplemental doses of vitamins and minerals on the progression, clinical outcomes, and treatment of COVID-19.
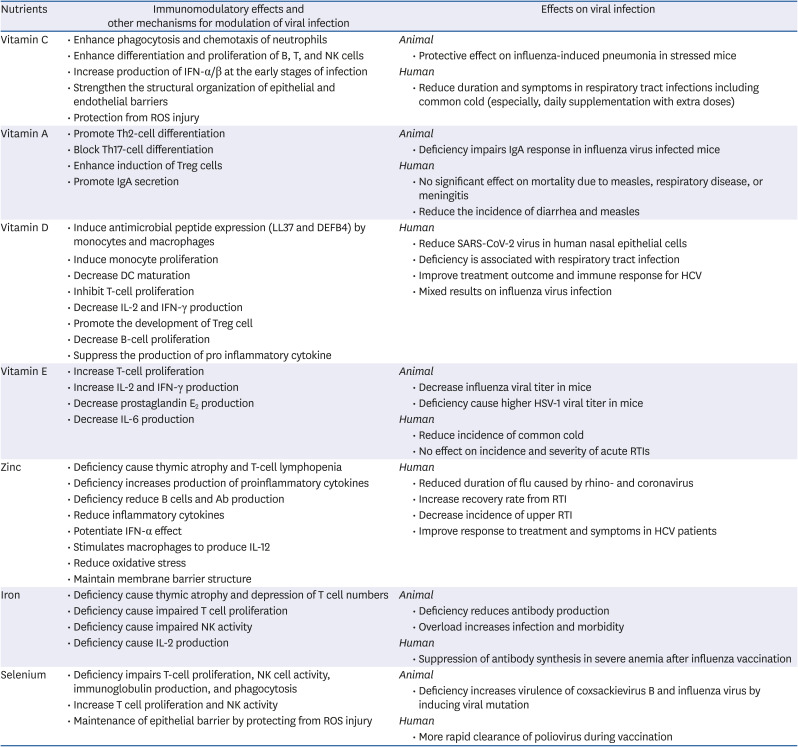
Table 1
Impact of nutrients on immune responses and viral infections
DC, dendritic cell; HCV, hepatitis C virus; HSV, Herpes simplex virus; IFN, interferon; IL, interleukin; NK cell, natural killer cell; ROS, reactive oxygen species; IgA, immunoglobulin A; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2; RTI, respiratory tract infection; Th, T helper; Treg cell, regulatory T cell.
![]()
Go to :
 XML Download
XML Download