

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The incidence of obesity has increased notably in adults and children owing to an increase in pervasive obesogenic lifestyle changes [1]. The increased incidence and progression, as well as mortality resulting from cancers have been considered to be associated with obesity [2]. Global research focusing on the prevalence of musculoskeletal diseases and disorders has increased owing to the increased prevalence of obesity [3,4]. Obesity can induce fat infiltration of skeletal muscle, mitochondrial dysfunction, or muscle atrophy, related to physical performance [5]. Studies have shown that people with higher muscle mass and strength have a lower mortality risk and body mass index [6,7].
Extreme diets that are either high-fat or high-fat and high-sucrose liquid (HFHS) lead to mitochondrial dysfunction, neuromuscular dysfunction, and lower exercise (EX) capacity of skeletal muscle [8-10]. In particular, sugar-sweetened beverages (SSBs) are associated with an increased mortality rate owing to circulatory diseases [11]. An increase in the total consumption of SSBs is correlated with an increase in the incidence of type 2 diabetes mellitus [12]. Sucrose has been reported to increase intermuscular fat [13], and this fat formation is further associated with decreased muscle function [14]. Chronic sugar consumption is related to autophagy regulation and lipid accumulation in skeletal muscle [15].
Autophagy can participate in mitochondrial metabolism by regulating lipid droplets (LD) [16]. Autophagy plays an essential role in maintaining muscle mass and healthy skeletal muscle [17,18] by removing old and damaged cellular components, breaking down unutilized nutrients, and remodeling cellular structures. Meanwhile, over-nutrition has been shown to inhibit autophagy, thereby impairing various cellular homeostatic processes [19]. Even in a single EX, autophagy is involved in EX-induced metabolic regulation and is related to physical performance changes in the obesity model [20].
The prevention of muscle dysfunction in obese patients may provide a method for suppressing the development of obesity-related diseases. More studies on developing intervention methods and understanding the mechanism of muscle dysfunction caused by an unhealthy diet in obese individuals are essential. Thus, the purpose of this study was to verify whether obesity-related muscle dysfunction caused by a high-fat diet or high-sucrose liquid feeding could be restored via SSB restriction (SR). This study demonstrated the combined effects of SR and EX in obesity-related muscle dysfunction via autophagy signaling pathways.
METHODS
Animals
Male C57BL/6 mice (3-week-old, n=61) were purchased from Central Lab. Animal Inc. Three to four mice were housed per cage under constant temperature (22°C±2°C) and humidity (55% to 60%) with a 12-hour-light/12-hour-dark cycle in a specific pathogen-free animal facility. All groups were allowed to eat ad libitum. Food intake and body weight were recorded twice per week. After a week of acclimatization, all mice were randomly assigned to the following six groups: CON (normal diet+plain water, n=6), CON+EX (normal diet+plain water+exercise, n=6), HFHS (n=16), HFHS+EX (high-fat diet+high-sucrose liquid+exercise, n=11), SR (change high-sucrose liquid to plain water, high-fat diet+plain water, n=11), and SR+EX (high-fat diet+plain water+exercise, n=11) (Fig. 1).
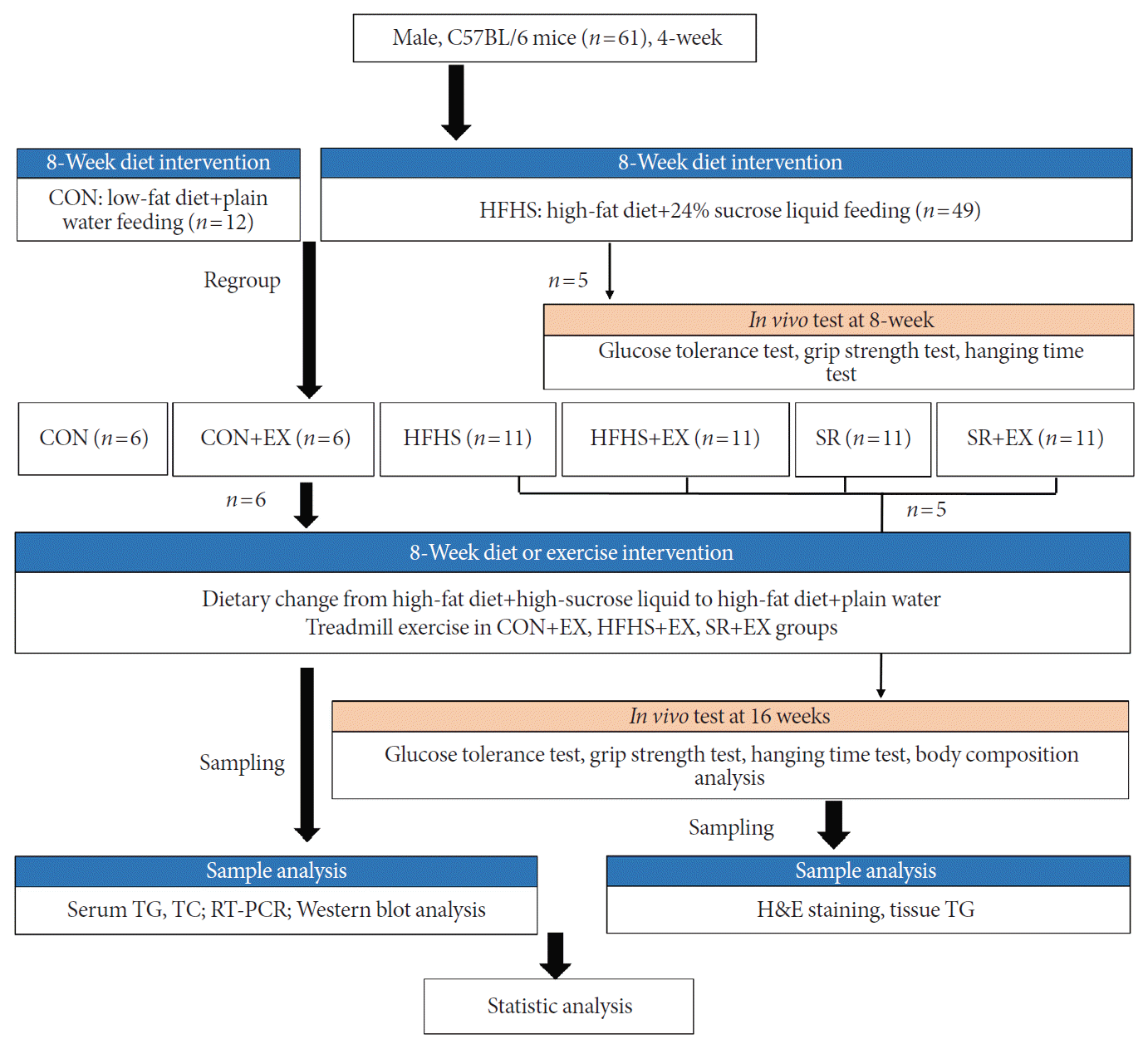
Fig. 1.
Schematic diagram of the experimental design. Male C57BL/6 mice, 4-week-old (n=61) were randomly assigned to six groups. Five mice from the high-fat and high-sucrose liquid (HFHS) group were subjected to glucose tolerance test and muscle performance test before re-grouping. After diet or exercise intervention, five mice from each of the eight groups were subjected to glucose tolerance test, muscle performance test, morphological observation, and tissue lipid quantification. Moreover, the other six mice from the groups as mentioned above were subjected to biological detection. Control (CON), normal diet+plain water; EX, exercise; SR, sugar-sweetened beverages (SSB) restriction, high-fat diet+plain water; TG, triglyceride; TC, total cholesterol; RT-PCR, reverse transcription-polymerase chain reaction.
![]()
Five HFHS mice were subjected to a glucose tolerance test after 8 weeks of feeding to evaluate the obese model. After another 8 weeks of diet or EX intervention, five mice from each of the six groups were subjected to glucose tolerance test, muscle performance test, body composition analysis, morphological observation, and tissue lipid quantification. The other six mice from the groups as mentioned above were subjected to biological detection. The high-fat diet and sucrose were purchased from Research Diets (D12492; New Brunswick, NJ, USA) and Central Lab. Animal Inc. (Seoul, Korea), respectively (Supplementary Table 1).
Exercise program
The mice in the EX groups were acclimatized and trained on an Exer-3/6 open treadmill (Columbus Instruments, Columbus, OH, USA) with 12° uphill incline for 2 days. The mice ran for 5minutes at 8m×min−1, and for 5minutes at 8m×min−1 on day 1, followed by another 5minutes at 10m×min−1 on day 2. On day 3, the mice were subjected to and were then trained on the treadmill with 12° uphill incline for 60minutes×day−1, 5 days×week−1 at 12 m×min−1 for 8 weeks (Supplementary Table 2) [21].
Glucose tolerance test
The intraperitoneal glucose tolerance test was performed after a 16-hour fast following the 2-day wash-out period via EX intervention. Glucose water solution at a concentration of 250 mg× mL−1 was injected at a dose of 2.5 g×kg−1 body weight. The blood glucose level in tail vein blood was determined using a glucometer (Accu-Chek Go, Roche Diagnostic, Mannheim, Germany). The blood glucose levels at 0, 30, 60, and 120 minutes were measured after glucose water injection. Tail snipping method was used to obtain blood.
Grip strength test
The mice were placed on a tension grid of grip strength meter while restrained manually by the scruff of the neck and base of the tail. After visual confirmation of firm gripping, the mouse was gently pulled back until it released its grip from the grid. Three trials were performed on each mouse with the most significant force used for analysis. The grip strength test values were normalized to body weight [22].
Hanging test
Mice were placed on the grip 25 cm above soft bedding and then placed the mouse on the grid so that it grasps it with its four paws. The test session ended for mice that could hang for a duration of 600 seconds. Mice fall grid and give up to two more tests before 600 seconds. The maximum hanging time was used for the analysis. The results from the hanging tests were multiplied by body weights to adjust the potential effects of body weight on test performance [23].
Body composition analysis
Total body composition (fat mass, lean mass) was evaluated using the Mini spec Contrast Agent Analyzer (Bruker Optik, Ettlingen, Germany) immediately before killing the mice.
Sample preparation
After an 8-week EX and/or dietary intervention, the mice were subjected to fasting and only plain water was provided. After 16 hours of fasting, the mice were anesthetized using isoflurane. All serum blood samples were obtained using cardiac puncture in serum separator tubes (BD Microtainer, Franklin Lakes, NJ, USA). After incubating the samples at room temperature for coagulation to occur, the tubes were centrifuged at 3,000 ×g for 10 minutes at 4°C, and the transparent supernatant was carefully separated. The tissues of muscles such as quadriceps (QUAD), gastrocnemius (GAS), soleus (SOL), tibialis anterior (TA), and extensor digitorum longus (EDL), and adipose tissues such as inguinal white adipose tissue (iWAT; subcutaneous fat), epididymal white adipose tissue (eWAT; visceral fat), and interscapular brown adipose tissues were collected and weighed. The tissues were frozen in liquid nitrogen and stored at −80°C, post collection.
Lipid analysis
The concentration of triglycerides (TG), extracted from the GAS muscle, was analyzed using reagent kits according to the manufacturer’s protocol in a serum separator tube (BD Microtainer, Franklin Lakes, NJ, USA) [24]. The serum TG and total cholesterol (TC) levels were measured using kits (Asan Pharm. Co. Ltd., Seoul, Korea).
Total RNA extraction
The total RNA was extracted from muscles using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s instructions. AccuPower CycleScrip RT PreMix (dN6) (Bioneer, Daejeon, Korea) was used to synthesize cDNA. The CFX9 Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) and SensiFAST SYBR Lo-ROX Kit (BIO-94020) were used to amplify target genes. The gene expression levels were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The gene expression levels of the autophagy pathway-related gene, Parkin, were analyzed using primers.
Total protein extraction and Western blot analysis
QUAD tissue (50 g) was added to 500 µL of lysis buffer (RIPA buffer, Thermo Fisher Scientific, Waltham, MA, USA; #89900), protease (Roche, #4693159001), and phosphatase (Sigma-Aldrich, St. Louis, MO, USA; #4906845001). All samples were homogenized using an ultrasonic homogenizer with the following sonication cycle: 30 seconds ‘on’ and 30 seconds ‘off,’ under cold conditions. The samples were centrifuged at 12,000 rpm for 15 minutes at 4°C, and the supernatant was transferred into a new tube. The total protein concentration present in the sample was quantified using a bicinchoninic acid assay kit, and was then subjected to Western blot analysis using antibodies against proteins involved in autophagy—anti-microtubule-associated proteins 1A/1B light chain 3 (LC3; Sigma, 1:500), anti-P62 (Cell Signaling, Danvers, MA, USA; 1:500), anti-BCL2 interacting protein 3 (BNIP3; Cell Signaling, 1:500), anti-AMP-activated protein kinase (AMPK; Cell Signaling, 1:500), anti-p-AMPK (Cell Signaling, 1:500), anti-mechanistic target of rapamycin (mTOR; Cell Signaling, 1:500), anti-p-mTOR (Cell Signaling, 1:500), and anti-Beclin1 (Cell Signaling, 1:500) and mitochondrial biosynthesis—anti-peroxisome proliferator-activated receptor γ coactivator-1α (PGC1α; Cell Signaling, 1:500). Anti-GAPDH antibodies (Sigma, 1:3,000) were also used for the analysis.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 8 (GraphPad Software Inc., La Jolla, CA, USA), and the data are represented either as mean±standard deviation or mean±standard error of the mean. P values <0.05 were considered as statistically significant. Two-way analysis of variance was used to analyze the data for examining the independent and combined effects of EX and SR on mice. Multiple comparisons were used for two interventions between different groups.
RESULTS
Effects of SR and exercise on body weight and food intake
The body weight of the SR group mice continued to rise after the restriction, as shown in the images of mice captured at the time of sacrifice (Fig. 2A and B). Even though the food intake was significantly (P<0.01) increased after the SR intervention (Fig. 2D), the total calorie intake of the SR group during 8 weeks was significantly decreased (P<0.05) when compared with the HFHS group (Fig. 2C). SR showed a significant increase in the wet weight of the subcutaneous fat, iWAT, and visceral fat, eWAT (Fig. 2E), but not when the adipose mass was normalized (adipose tissue mass [mg]/body weight [g]) (Fig. 2F). The body composition analysis confirmed that SR did not affect the total fat mass (Fig. 2G).
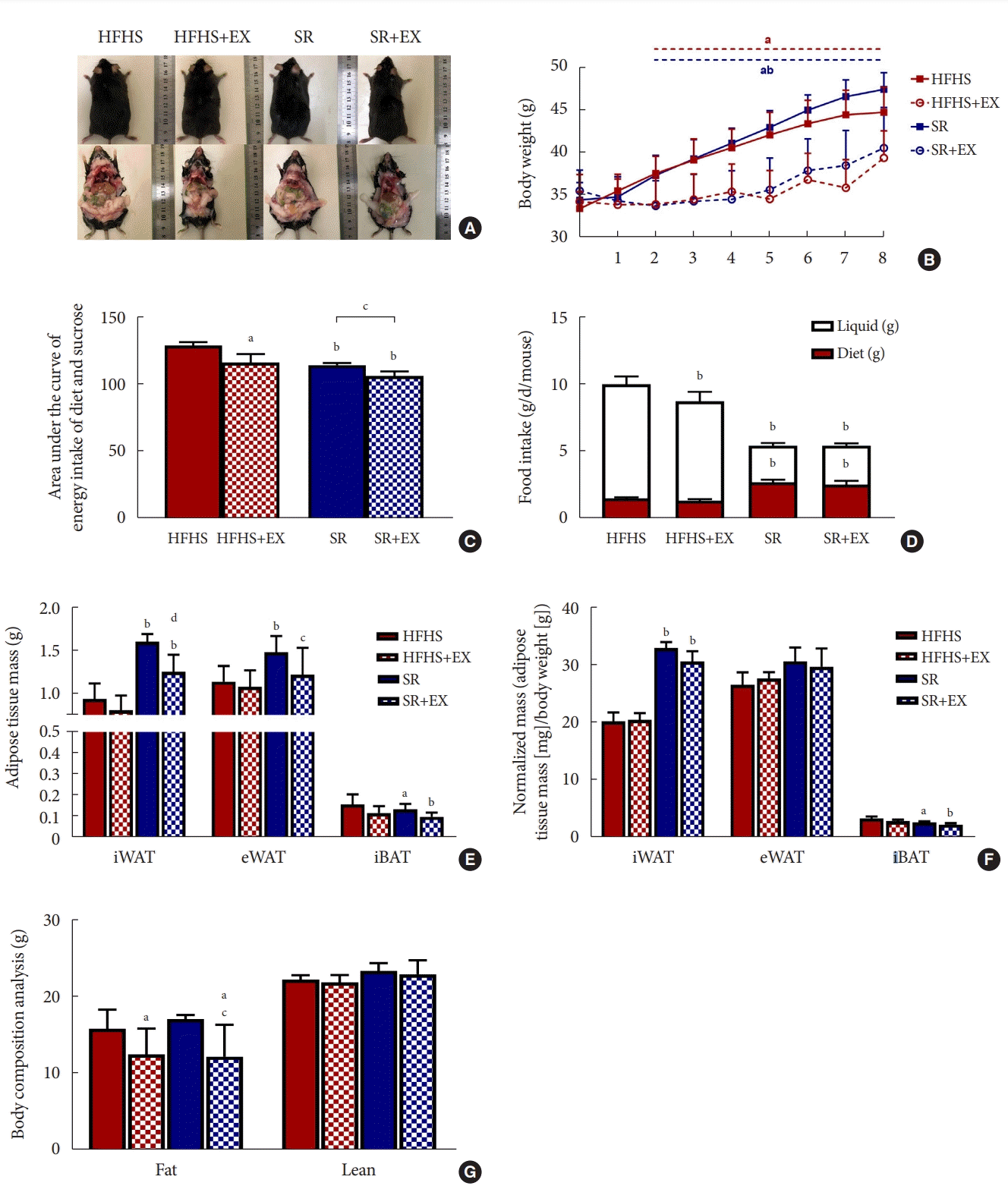
Fig. 2.
Effects of SR and exercise on body weight and food intake. (A) Images of mice. (B) Change in the body weight (n=5). (C) The area under the curve of energy intake (n=5). (D) Diet or liquid intake (n=5). (E) Wet weight of adipose tissues (n=6).
(F) Normalized wet weight of adipose tissues (n=6). (G) Results of body composition analysis (n=5). All values are shown as the mean±standard deviation. HFHS, high-fat and high-sucrose liquid; EX, exercise; SR, sugar-sweetened beverages (SSB) restriction, high-fat diet+plain water; iWAT, inguinal white adipose tissue; eWAT, epididymal white adipose tissue; iBAT, interscapular brown adipose tissue. aP<0.05, bP<0.01 vs. the HFHS group and cP<0.05, dP<0.01 vs. the SR group.
![]()
In Fig. 2B, the SR+EX group showed a slow pattern of body weight growth (P<0.05) than the HFHS and SR mice with total lower energy intake (P<0.01) (Fig. 2C). It consistently showed a change in body weight when compared to the HFHS and HFHS+EX groups. Moreover, it demonstrated a significant (P<0.01) difference starting from the first to the seventh week of intervention. In addition, the reduction of fat in the SR+EX group could be visually observed (Fig. 2A), consistent with the results suggesting that the SR+EX group showed a significant (P<0.05) decrease in fat mass in comparison to the HFHS and SR groups (Fig. 2G). The iWAT and eWAT mass decreased when compared to the SR and SR+EX groups; however, there was no significant difference observed when the mass was normalized using body weight (Fig. 2E and F). Therefore, the results indicated that EX could prevent the increase in body weight, energy intake, and fat mass in the absence of SSB consumption. However, no combined effect of EX and SR on the mentioned parameters were observed (Table 1).
Table 1.
Independent and combined effects of exercise and sugar-sweetened beverage restriction on obesitya
| Variable |
Diet |
Exercise |
Diet×exercise |
|||
|---|---|---|---|---|---|---|
| F | P value | F | P value | F | P value | |
| BW, g | 1.644 | 0.218 | 16.61 | 0.001b | 0.257 | 0.620 |
| AUC for energy intake | 29.17 | 0.001b | 20.42 | 0.002b | 0.984 | 0.350 |
| Fat mass from body composition analysis, g | 0.140 | 0.714 | 9.670 | 0.067 | 0.350 | 0.563 |
| Adipose tissue mass, g | ||||||
| iWAT | 56.03 | <0.001b | 10.58 | 0.005b | 2.094 | 0.167 |
| eWAT | 5.676 | 0.030b | 2.532 | 0.131 | 1.022 | 0.327 |
| iBAT | 1.972 | 0.179 | 6.582 | 0.021b | 0.047 | 0.832 |
| Normalized adipose tissue mass, mg/g | ||||||
| iWAT/BW | 63.32 | <0.001b | 0.515 | 0.483 | 0.789 | 0.387 |
| eWAT/BW | 1.721 | 0.207 | 0.001 | 0.971 | 0.188 | 0.670 |
| iBAT/BW | 5.80 | 0.028b | 2.175 | 0.159 | 0.081 | 0.779 |
| Fasting blood glucose, mg/dL | 24.48 | <0.001b | 1.746 | 0.205 | 0.261 | 0.616 |
| AUC of ipGTT | 11.43 | 0.004b | 13.43 | 0.002b | 9.495 | 0.007b |
| Serum TC | 41.00 | <0.001b | 6.160 | 0.018b | 2.593 | 0.090b |
| Serum TG | 3.857 | 0.031b | 9.321 | 0.005b | 0.349 | 0.708 |
| QUAD TG | 4.244 | 0.031b | 3.755 | 0.069 | 3.841 | 0.041b |
| Normalized skeletal muscle mass, mg/g | ||||||
| QUAD/BW | 73.67 | <0.001b | 2.135 | 0.153 | 1.603 | 0.216 |
| GAS/BW | 54.74 | <0.001b | 0.001 | 0.981 | 1.236 | 0.303 |
| TA/BW | 115.6 | <0.001b | 0.707 | 0.405 | 1.040 | 0.364 |
| SOL/BW | 12.49 | <0.001b | 7.092 | 0.008b | 0.329 | 0.722 |
| EDL/BW | 3.153 | 0.055 | 2.183 | 0.149 | 2.474 | 0.099 |
| Hanging time (sec)×BW(kg) | 0.525 | 0.480 | 27.87 | <0.001b | 1.272 | 0.277 |
| Grip strength (g)/BW(kg) | 0.211 | 0.652 | 9.440 | 0.007b | 0.993 | 0.334 |
| Autophagy-related proteins | ||||||
| LC3I | 14.49 | <0.001b | 2.877 | 0.105 | 1.137 | 0.340 |
| LC3II | 4.877 | 0.018b | 21.39 | 0.001b | 3.517 | 0.048b |
| LC3II/LC3I ratio | 6.512 | 0.006b | 6.642 | 0.018b | 2.528 | 0.104 |
| P62 | 4.189 | 0.028b | 1.742 | 0.200 | 0.943 | 0.404 |
| BNIP3 | 0.843 | 0.445 | 6.079 | 0.022b | 1.860 | 0.180 |
| Beclin1 | 4.385 | 0.024b | 0.017 | 0.899 | 2.587 | 0.096 |
| Glucose metabolism | ||||||
| p-AMPK/AMPK ratio | 0.499 | 0.614 | 7.387 | 0.013b | 2.487 | 0.106 |
| p-mTOR/mTOR ratio | 11.51 | 0.001b | 3.000 | 0.095 | 0.555 | 0.580 |
| GLUT4 | 8.250 | 0.002b | 5.249 | 0.032b | 0.280 | 0.758 |
| Mitochondria metabolism | ||||||
| PGC1α | 2.632 | 0.094 | 8.683 | 0.007b | 0.224 | 0.801 |
| Myogenesis | ||||||
| MyoD | 5.478 | 0.011b | 8.916 | 0.007b | 0.739 | 0.489 |
BW, body weight; AUC, area under the curve; iWAT, inguinal white adipose tissue; eWAT, epididymal white adipose tissue; iBAT, interscapular brown adipose tissue; ipGTT, intraperitoneal glucose tolerance test; TC, total cholesterol; TG, triglyceride; QUAD, quadriceps; GAS, gastrocnemius; TA, tibialis anterior; SOL, soleus; EDL, extensor digitorum longus; LC3, microtubule-associated proteins 1A/1B light chain 3; BNIP3, BCL2 interacting protein 3; AMPK, AMP-activated protein kinase; mTOR, mechanistic target of rapamycin; GLUT4, glucose transporter type 4; PGC1α, peroxisome proliferator-activated receptor γ coactivator-1α; MyoD, myogenic differentiation.
![]()
Effects of SR and exercise on glucose and lipid metabolism
An 8-week intervention of SR resulted in a significant increase in the fasting blood glucose levels in the HFHS group as compared to before intervention (0 week) (Table 1, Fig. 3A). Compared to the HFHS group, the SR group had a more impaired (P<0.05) glucose tolerance (Fig. 3B) and increased blood glucose area under the curve (AUC; P<0.01) than the HFHS group (Fig. 3C). The EX (P<0.01) improved glucose tolerance at both 90 and 120 minutes in the SR group indicating a significant (P<0.01) improvement in the blood glucose AUC values (Fig. 3B and C); however, these effects were not observed on the HFHS group.
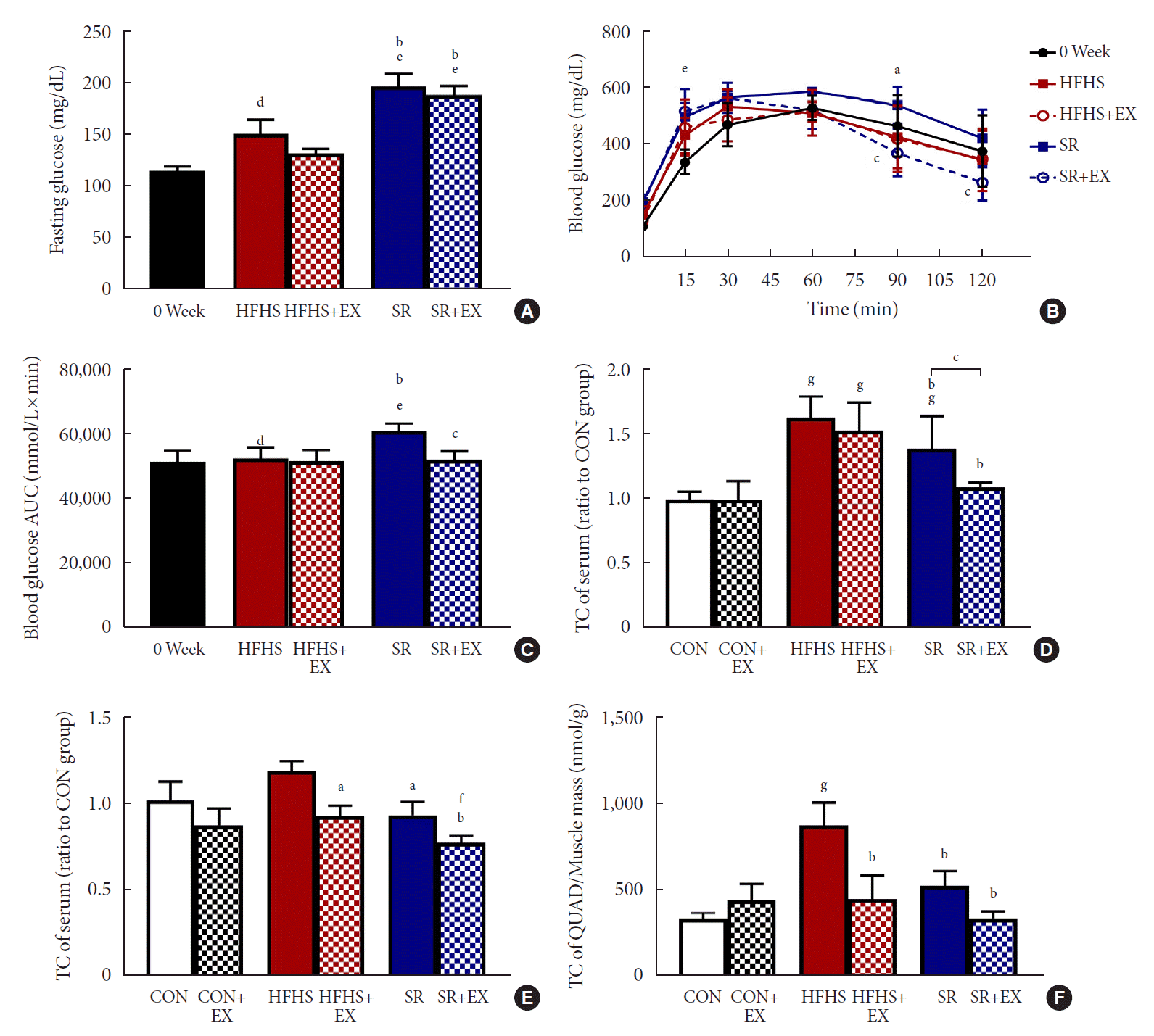
Fig. 3.
Effects of SR and exercise on glucose and lipid metabolism. (A) Fasting glucose (n=5). (B) Intraperitoneal glucose tolerance test (ipGTT) curve (n=5). (C) The area under the curve (AUC) of ipGTT (n=5). (D) Total cholesterol (TC) (n=6). (E) Triglyceride (TG) concentration (n=6) of serum. (F) TG concentration (n=6) of quadriceps (QUAD). All values are shown as the mean±standard deviation. 0 week, point before intervention; HFHS, high-fat and high-sucrose liquid; EX, exercise; SR, sugar-sweetened beverages (SSB) restriction, high-fat+plain water. aP<0.05, bP<0.01 vs. the HFHS group, cP<0.01 vs. the SR group, dP<0.05, eP<0.01 vs. 0 week, fP<0.01, and gP<0.01 vs. the control (CON) group
![]()
For the analysis of lipid metabolism, an 8-week SR intervention significantly decreased the TC (P<0.05) and TG (P<0.01) levels in both serum and muscle samples (Fig. 3D-F). For the effect of EX, the serum TC level was (P<0.01) decreased in the SR group when compared with the SR+EX group, but not in the HFHS group when compared with the HFHS+EX group (Fig. 3D). These results indicated that SR along with EX prevented the regulation of serum TC levels.
Effects of SR and exercise on skeletal muscle performance
The normalized muscle mass of QUAD, GAS, TA, SOL, and EDL remained unaffected in the SR group as compared to in the HFHS group (Fig. 4A-E). Meanwhile, both the hanging time and grip strength also remained unaffected in the SR group as compared to in the HFHS group (Fig. 4F and G). EX improved (P<0.05) the muscle mass of QUAD in both the HFHS and SR groups (Fig. 4A). EX also improved SOL and EDL muscle mass, but this improvement was not significant in the SR group (Fig. 4D and E). EX also improved the poor hanging performance of the SR+EX group, similar to the HFHS+EX group (Fig. 4F). Moreover, EX significantly improved the grip strength of the SR+EX group, but not of the HFHS+EX group (Fig. 4G). These results indicated that EX could improve the hanging performance with improved muscle mass. However, the SR intervention alone could not improve muscle mass despite improving grip strength without EX training.
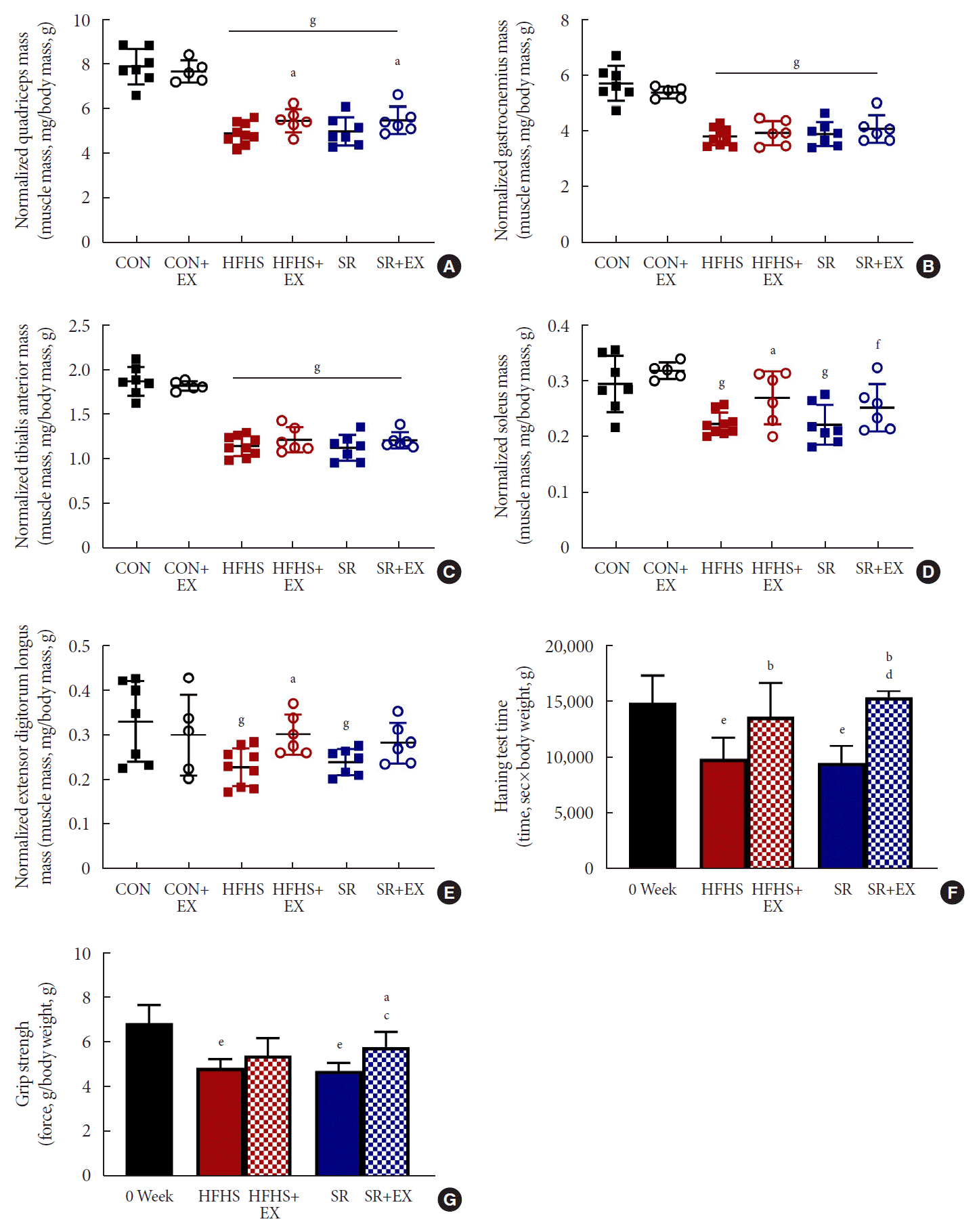
Fig. 4.
Effects of SR and exercise on skeletal muscle performance. (A) Normalized quadriceps mass (n=6). (B) Normalized gastrocnemius mass (n=6). (C) Normalized tibialis anterior mass (n=6). (D) Normalized soleus mass (n=6). (E) Normalized extensor digitorum longus mass (n=6). (F) Normalized hanging time (n=5). (G) Normalized grip strength (n=5). All values are shown as the mean±standard deviation. 0 week, point before intervention; control (CON), normal diet+plain water; EX, exercise; HFHS, high-fat and high-sucrose liquid; SR, sugar-sweetened beverages (SSB) restriction, high-fat+plain water diet. aP<0.05, bP<0.01 vs. the HFHS group, cP<0.05, dP<0.01 vs. the SR group, eP<0.01 vs. 0 week, and fP<0.05, gP<0.01 vs. the CON group.
![]()
Effects of SR and exercise on the expression of the genes involved in the autophagy pathway
The expression of genes involved in the autophagy pathway were showed in the images of immunoblotting (Fig. 5A). The ratio of LC3II/LC3I in the SR group remained unaffected as compared to in the HFHS group (Table 1, Fig. 5B). EX increased the activation of LC3II, but not the LC3II/LC3I ratio. In addition, SR reduced the intracellular levels of P62 accumulated in the HFHS+EX group (P<0.05) as compared to in the HFHS group (Fig. 5C). SR also significantly improved the expression of BNIP3 (P<0.01), which was consistent with the effects of EX observed on the HFHS group (Fig. 5D). There was no combined effect of EX and SR on the expression of BNIP3 and was no effect of SR or EX on the expression of Beclin1 (Table 1, Fig. 5E).
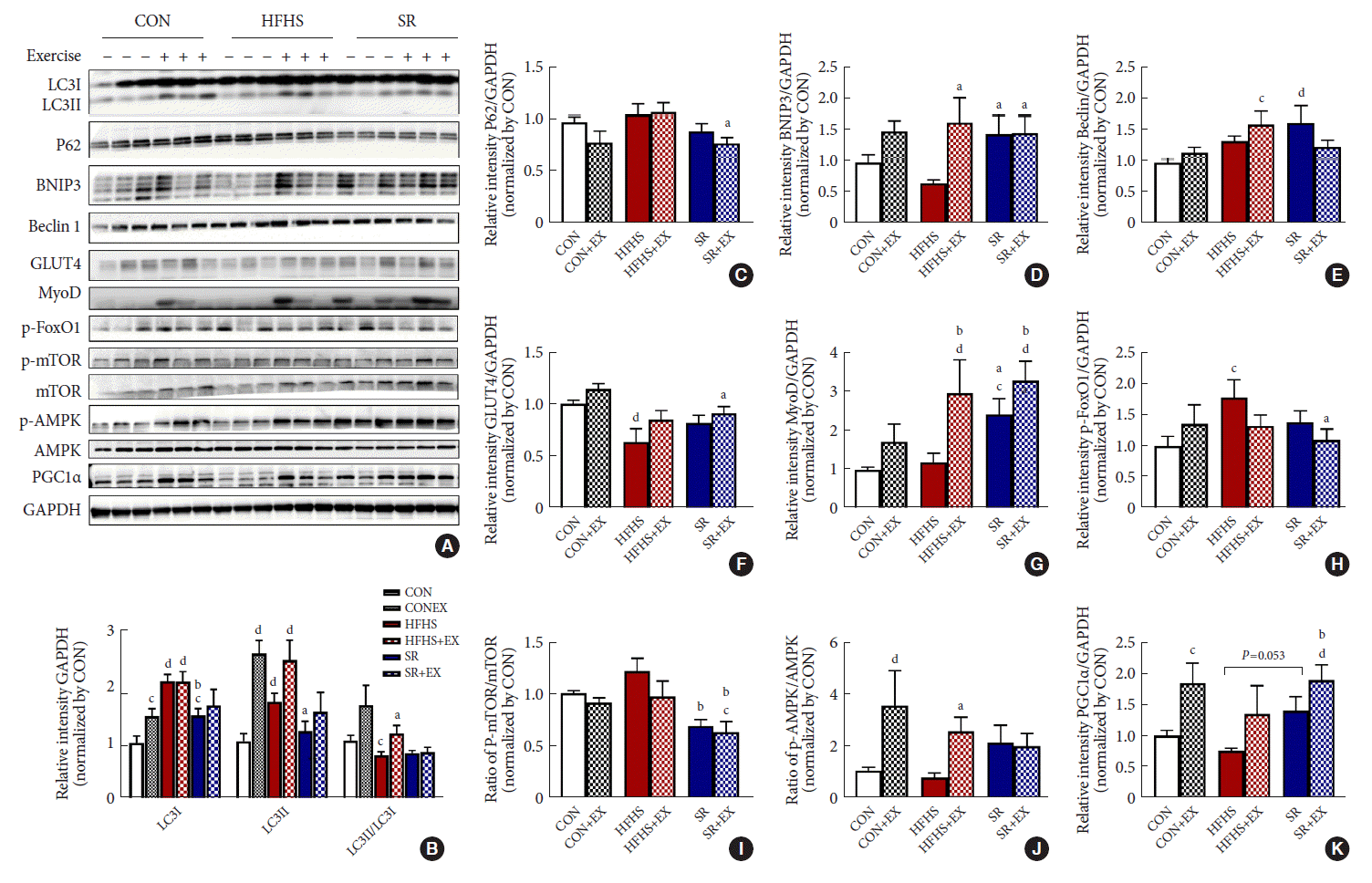
Fig. 5.
Effects of SR and exercise on the expression of the autophagy pathway-related proteins. (A) Images of immunoblotting results. (B) The graph shows the expression of microtubule-associated proteins 1A/1B light chain 3 (LC3I), LC3II, and LC3II/LC3I ratio, (C) P62, (D) BCL2 interacting protein 3 (BNIP3), (E) beclin1, (F) glucose transporter type 4 (GLUT4), (G) myogenic differentiation (MyoD); (H) p-forkhead box O1 (FoxO1), (I) p-mechanistic target of rapamycin (mTOR)/mTOR ratio, (J) p-AMP-activated protein kinase (AMPK)/AMPK ratio, (K) peroxisome proliferator-activated receptor γ coactivator-1α (PGC1α), determined by densitometric quantification. All values are shown as the mean±standard error of the mean (n=6). GAPDH, glyceraldehyde-3-phosphate dehydrogenase; Control (CON), normal diet+plain water; EX, exercise; HFHS, high-fat and high-sucrose liquid; SR, sugar-sweetened beverages (SSB) restriction, high-fat+plain water diet. aP<0.05, bP<0.01 vs. the HFHS group, cP<0.05, and dP<0.01 vs. the CON group.
![]()
Both EX and SR intervention significantly decreased glucose transporter type 4 (GLUT4) expression by long-term HFHS feeding (P<0.05) in the SR+EX group (Fig. 5F). Significant (P<0.05) increase in the expression of myogenic differentiation (MyoD) was observed in the SR groups, similar to the effect of EX on the HFHS group (Fig. 5G). Both EX and diet intervention decreased the phosphorylation of forkhead box O1 protein that was increased by HFHS, especially in the SR+EX group (P<0.05) (Fig. 5H). Therefore, this indicated a negative regulation of adipogenesis in the SR+EX group.
SR significantly decreased the p-mTOR/mTOR ratio that was increased by HFHS feeding (Table 1, Fig. 5I). SR increased the ratio of p-AMPK/AMPK, though not significantly, which was consistent with the effect of EX observed on the CON group when compared to the CON+EX group and the HFHS group when compared to the HFHS+EX group (Fig. 5J). Both EX and SR intervention increased the expression of PGC1α, especially in the SR+EX group (P<0.01) (Fig. 5K). SR had no effect on the mRNA expression of Parkin, which was decreased by long-term HFHS feeding, but recovered by EX, though not significantly (Supplementary Fig. 1).
DISCUSSION
Excessive sucrose and fat intake causes obesity and is the most frequent cause of death among diabetic patients [25]. Obesity has been reported to induce muscle dysfunction, metabolic syndrome, and other diseases [4]. Previous studies have indicated that increased fat in muscles decreased muscle mass in obese patients [5], and subsequent decrease in muscle function, in particular, has been proved to increase mortality rate [6].
The dangers of SSB consumption are widely publicized, and some countries have formulated regulations concerning SSB consumption [26]. Studies have shown that SSBs are the cause of weight gain, and therefore reducing the intake of SSBs might be useful for weight loss [27]. However, our results showed that SR did not decrease body weight (Fig. 2B) or fat mass (Fig. 2G) as expected, which was consistent with a previous animal study demonstrating that sucrose had little effect on body weight [28]. SR increased the wet weight of iWAT and eWAT in comparison with the HFHS group (Fig. 2E and F). The fasting glucose levels and glucose tolerance were not improved by the intervention of SR (Fig. 3A-C), which contradicted the results of a previous study showing that a carbohydrate-restricted diet might reduce the risk of muscle damage in patients with a disease of muscle metabolism [29].
Similarly, SR had no effect on the muscle performance related to endurance and maximum grip, which was consistent with the findings of previous research that obesity affects muscle function, and the skeletal muscle becomes similar to aged mice muscle [30]. Similarly, at the molecular level, SR increased the protein expression of GLUT4 (Fig. 5F) decreased by HFHS feeding despite no significant. It reduced TC and TG levels in serum and tissues compared to the HFHS group. This indicated that SR promoted the assimilation of glucose and lipids in blood and tissues. Combined with the aforementioned regulatory effect of SR, we know that it was consistent with the previous findings that sucrose intake could increase TG and TC levels in the blood [31]. However, a meta-analysis study showed that the replacement of sucrose with fructose in beverages could reduce postprandial glucose and insulin levels without raising TG concentrations [32]. We found that although the total energy intake was reduced in the SR group, the intake of a high-fat diet was considerably increased because of the ad libitum intake, which could be the possible reason why the muscle function of the SR group did not recover. These results are consistent with the findings of the previous studies that liquid sucrose consumption might be an essential contributor to insulin resistance [33,34].
As previously mentioned, body weight had a remarkable influence on EX performance. However, EX significantly decreased body weight and fat mass in the EX groups as compared to in the HFHS and SR groups (Fig. 2B and G), which was consistent with improved performance during the hanging test performed on the EX groups (Fig. 4F). Moreover, the visceral fat quality was proved to be inversely related to the muscle mass [35]. In this study, the significant effects of EX on the mass of eWAT (Fig. 2E) of the SR group were not evident in the HFHS group. This was consistent with the result that EX could improve grip strength in the SR+EX group, but not in the HFHS+EX group (Fig. 4G).
Moreover, EX improved glucose tolerance and serum TC levels in the SR+EX group, but not in the HFHS+EX group, which was consistent with the change in GLUT4 expression. The results mentioned above suggest that weight and fat mass affect the hanging performance. Simultaneously, sucrose intake blocked the positive effects of EX on lipid metabolism and affected the improved grip strength due to EX.
According to the results as mentioned above, EX performance had not improved with the intervention of SR. Moreover, we found that with the intervention of SR, LC3II was decreased, and the ratio of LC3II/LC3I did not change, which was consistent with previous studies showing that excessive calorie intake could inhibit autophagy [19]. However, relative to HFHS, SR decreased the intracellular levels of P62 and increased the expression of Beclin1 and BNIP3 significantly, suggesting that SR could cause an increase in mitophagy. Previous studies have shown that mitochondria can regulate lipid metabolism through autophagy [16]. In addition, excessive sugar intake has been shown to increase ectopic fat deposits in muscles [13]. Therefore, it is speculated that SR inhibits ectopic fat deposition by promoting mitophagy. The changes in P62 were consistent with the effects of EX on high blood TC level, impaired glucose tolerance, and low maximum grip strength induced by HFHS only in the SR+EX group and not in the HFHS+EX group. P62 was associated with LD protein and adipose-differentiation associated protein in regulating lipid phagocytosis and intramyocardial lipid accumulation [36]. P62 expression affects adipogenesis, insulin signaling, and brown adipose tissue thermogenesis in patients with metabolic diseases and cancers [37]. Our results were consistent with accumulated P62 levels in the HFHS group which resulted in the failure of muscle function recovery.
SR groups decreased P62 levels, especially in the SR+EX group. SR significantly increased the expression of BNIP3, which was consistent with the study that a high-sucrose diet could damage mitochondria [38,39]. In addition, SR+EX significantly increased the expression of PGC1α, and the mRNA levels of Parkin and MyoD, which could restore mitochondrial activity and muscle growth in the SR+EX group. Additionally, the improved EX performance of the SR group was observed. It is speculated that SR could enhance the quality of muscle, but due to SSB interruption, the change in calorie intake resulted in a large intake of the high-fat diet, increasing blood glucose and body weight. In addition, sugar has been shown to induce autophagy through the regulation of the mTOR pathway [40]. In this study, SR increased the p-AMPK/AMPK ratio, though not significantly, and decreased the p-mTOR/mTOR ratio, which was consistent with the change observed in autophagic response. These results showed that only SR+EX improved glucose tolerance, which was consistent with improved muscular performance.
Concerning the limitations of this study, studies have shown that restricting dietary intake can cause changes in autophagy. This study aimed to explore the effect of SSB deprivation on changes in the autophagic response in obese mice to determine the relationship between muscle function and metabolism. It is difficult to control the dietary intake of the SR group. Therefore, it may be weak to explain whether the results of this study were caused by SR intervention or excessive high-fat diet intake. Although SR induced mitophagy, it did not affect muscle function or blood glucose level as compared to HFHS. To explain this result, we investigated the total calorie intake. SR significantly increased fat intake and decreased calories simultaneously as compared to HFHS (Fig. 2C). Therefore, it is speculated that excessive fat intake is not the cause of unchanged muscle function. We can be sure that replacing SSB with water increased the appetite of mice for diet and thereby reduced water intake, resulting in increased blood sucrose, white adipose, and body weight. Second, we investigated the expression of autophagy-related genes only in the QUAD, which is a mixed fiber-type muscle. However, in a previous study, oxidative muscle and glycolytic muscle were proved to have different basal autophagic responses [41]. We speculated that the autophagic response with intervention might vary in different muscles and could be related to muscle function; thus, lacking the detection of autophagy-related protein expression in different muscles is another limitation of this study.
In conclusion, SR promoted TG assimilation in muscle, serum TG, and TC, and restored the expression of mitophagy-related proteins in QUAD, but had no effect on muscle function, which might be associated with an excessively high-fat diet. The combined effect of SR and EX resulted in the activation of mitophagy-related proteins and restoration of muscle function by enhancing grip strength, mitochondrial oxidative function, and the assimilation of fat and glucose in the muscle. Finally, we found that sucrose liquid intake limited the effect of EX on glucose tolerance and muscle strength. Therefore, based on the results, we suggest that limiting the intake of sugary beverages and controlling high-fat diet intake is an effective way to reduce body weight and restore glucose tolerance and muscle function.
 XML Download
XML Download