

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Obesity is defined as the abnormal or excessive accumulation of fat in the body from the World Health Organization (WHO). It is a significant risk factor for metabolic diseases such as cardiovascular disease, nonalcoholic fatty liver disease, hypertension, type 2 diabetes, and a specific type of cancer [1]. Adipose tissue, a critical organ for energy homeostasis, expands to store excess energy in obesity [23]. Adipocytes, the primary cell type in adipose tissue, play a crucial role in adipose tissue enlargement [4]. Adipose tissue enlarges by increases of preexisting mature adipocytes in size (hypertrophy) and/or of numbers of new adipocytes (hyperplasia) [56]. Hypertrophy appears in the progression of obesity to meet the need for additional fat storage and then trigger hyperplasia in adipose tissue [25]. The enlargement of adipocytes arises from increased lipogenesis to promote lipid storage [7]. Adipogenesis, a process of preadipocytes differentiate into mature adipocytes, plays a crucial role in obesity. The process is tightly regulated by a network of transcription factors that activate a large number of adipogenesis genes [89]. The key players in the transcriptional cascade of adipocyte differentiation are peroxisome proliferator-activated receptor γ (PPARγ) and members of CCAAT/enhancer-binding protein (C/EBP) family [810]. PPARγ and C/EBP are the master regulators of adipogenesis and induce the genes involved in lipogenesis, lipolysis, and insulin sensitivity, such as glucose transporter 4 (GLUT4), adipocyte protein 2 (aP2), lipoprotein lipase (LPL), etc. [49]. Therefore, the inhibition of adipogenesis and triglyceride (TG) accumulation in the adipocytes is one of the winning strategies for the prevention of obesity [11].
Herbal medicines are claimed to effectively control obesity and its associated metabolic diseases [1213]. Ginger (Zingiber officinale) is one of the most commonly used spices and traditional medicine to treat arthritis, sprains, muscular pains, sore throats, nausea, and stroke in Asian countries [1415]. Ginger contains several bioactive phenolic compounds, including non-volatile and pungent compounds such as gingerol, shogaol, paradol, and zingerone [16]. The potential effects and its mechanisms actions of ginger and its active component, i.e., gingerol, shogaol, zingerone, etc., on the prevention of obesity have been reported in cell and animal studies [1415].
The composition and its functional properties of herbs or vegetables can differ from cultivation, storage, and processing methods. Ginger is heated or steamed when used as a spice or traditional medicine. Several studies reported that the heating or steaming process of herbs or root vegetables changes the chemical profiles [1718], and these changes can affect or improve their bioactivities [19202122]. Steamed ginger exerted different chemical profiles with enhanced anticancer effect [23] and adrenaline secretion [24]. In the previous study, the ethanolic extract of steamed ginger (SGE) showed antioxidative activity and lowered serum lipids in obesity induced by a high-fat diet (HFD) in C57BL/6J mice [25]. However, little is known about the protective effects and its underlying mechanisms of steamed ginger on adipogenesis in adipocytes and diet-induced obese mice. In the present study, we investigated the anti-obesity effect and its underlying mechanisms of the steamed ginger on 3T3-L1 cells and C57BL/6J mice fed a HFD.
Go to : 
MATERIALS AND METHODS
Cell culture and viability assay
The 3T3-L1 cells murine preadipocytes (ATCC, Manassas, VA, USA) were cultured in DMEM (Lonza, Walkersville, MD, USA) supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY, USA), 100 U/mL of penicillin, 100 μg/mL of streptomycin (Hyclone, Logan, UT, USA) in a 37°C humidified cell culture incubator with 5% CO2. The cytotoxicity of SGE was assessed with the cell viability of 3T3-L1 preadipocyte cells. The cells were seeded at a final concentration of 1 × 104 and treated with different concentrations of SGE for 24, 48, and 72 h. After incubation, the MTT [3(4,5-dimetyl2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide] EZ-Cytox, Daeil Lab Service, Seoul, Korea] and incubated for 2 h and read on the absorbance at 450 nm using a microplate reader (MRX II, Dynex Technologies, Chantilly, VA, USA).
Cell Differentiation and SGE treatment
Two days after post-confluency, the preadipocytes were differentiated in DMEM supplemented with 10% FBS and differentiation cocktail containing 0.5 mM IBMX, 1 μM DEXA, and 10 μg/mL insulin (Sigma-Aldrich Co., St. Louis, MO, USA). After differentiation, the cells were maintained in 10% FBS and 10 μg/mL of insulin in DMEM for another 6 days. 90% of the cells had transformed into mature adipocytes, with the formation of intracellular lipid droplets. SGE was treated every other day during the differentiation period of 8 days.
Oil-red O staining and glycerol-3-phosphate dehydrogenase (GPDH) assay
The cells were stained with Oil-Red O solution (Sigma-Aldrich Co.) to determine oil droplets in adipocytes as previously described [27]. Briefly, the differentiated cells were fixed with 10% formalin and then stained with filtered Oil-Red O solution for 30 min. The cellular TG contents were measured by the cell lysate (5% Triton X-100) at 580 nm. The GPDH activity was determined using GPDH assay kits from Takara (Takara Bio Inc., Otsu, Japan).
Animal care and diet
Eight-week-old male C57BL/6J mice (Charles River Laboratories, Tokyo, Japan) were randomly divided into four groups (n = 10 per group, total n = 40): a normal diet (ND; 10% fat by wt, Research Diets, InC., NJ, USA), a HFD (60% fat by wt, Research Diets, Inc.), and HFD supplemented with 40 mg/kg or 80 mg/kg of SGE by wt (SGD4 or SGD8, respectively). All animal procedures were approved by the Institutional Animal Care and Use Committee at the Chonbuk National University (CBU 2016). The mice were housed in a polycarbonate cage under 12 h light/dark cycles. The composition of the experimental diets fed to the mice for 12 weeks are shown in Table 1. Body weight and food consumption were recorded weekly. After 12 weeks, mice were fasted for 12 h and sacrificed. The serum was collected by centrifugation of blood at 1,000 × g for 15 min at 4°C. The epididymal adipose fat pads were snap-frozen in liquid nitrogen or fixed in 10% formalin. The serum and tissue samples were stored at −80°C until use.
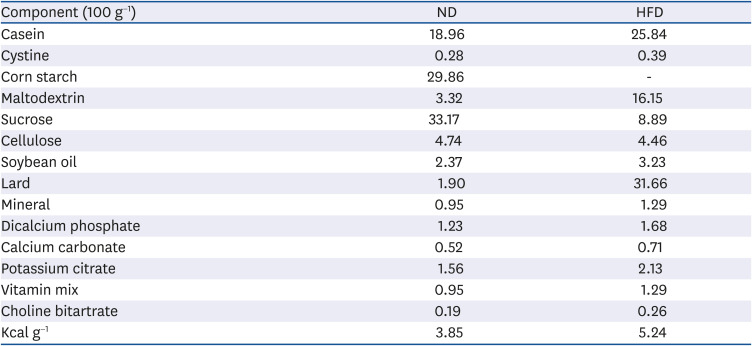
Table 1
Composition of experimental diets (unit: g)
ND: 10% of energy as fat (D12450B), HFD: 60% of energy as fat (D12492).
ND, normal diet; HFD, high-fat diet.
![]()
Serum chemistry and liver lipid measurements
Total cholesterol (TC) and TG concentrations in serum were determined by a commercial assay kit (Asan Pharmaceutical Co., Seoul, Korea). Serum alanine transaminase (ALT) and aspartate transferase (AST) activity were measured using a chemistry analyzer (Fuji Dri-Chem 3500, Fuji Photo Film Co., Tokyo, Japan). Serum glucose levels and insulin were measured by glucometer (Accu-check, Roche, Mannheim, Germany) and insulin ELISA kit (Alpeo, Sale, MA, USA), respectively. Leptin and adiponectin concentrations in serum were analyzed by assay kits (R&D Systems, Minneapolis, MN, USA), followed by the manufacturer's protocols. Hepatic TC and TG were measured enzymatically measured from the lipids extracted by Folch's methods, as we previously described [27].
Quantification of abdominal fat using micro-computed tomography (micro-CT)
The abdominal fat was imaged using high-resolution in vivo micro-CT (Skyscan, Konitch, Belgium) as previously described [2527]. The abdominal micro-CT scan images of anesthetized mice were captured at the level of L1-L5 intervertebral disk and analyzed using CT analyzer Version 1.16.4.1, Skyscan software (Skyscan).
Histology of epididymal adipose tissue
The formalin-fixed epididymal adipose tissue samples were embedded in paraffin. The paraffin-embedded tissues were cut at a thickness of 5 sµm and stained with hematoxylin-eosin. The tissue images were viewed using an Axiophot Zeiss Z1 microscope (Carl Zeiss, Gottingen, Germany) and analyzed with Analysis SIS3.2 software (Soft-Imaging System).
Gene expression analysis by quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was extracted using TRIzol reagent (Life Technologies, Inc., Carlsbad, CA, USA), and qRT-PCR analysis for gene expression was conducted as previously described using the SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and 7500 real-time PCR system (Applied Biosystems) [2829]. The primer sequences for qRT-PCR used in the present study are listed in Table 2. The glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control.
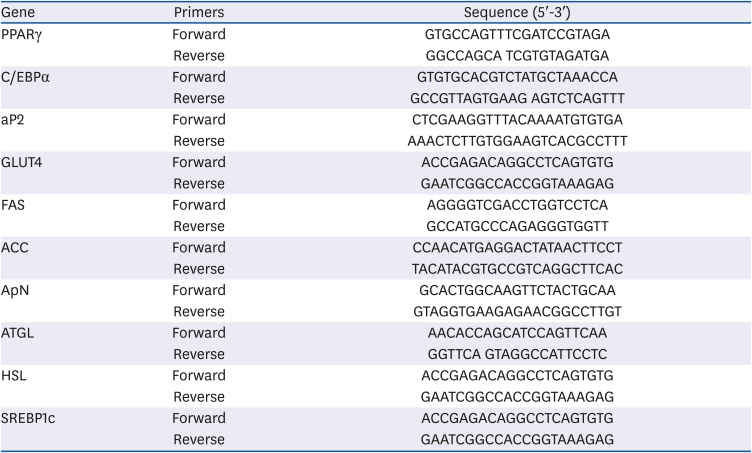
Table 2
Quantitative real-time polymerase chain reaction primer
PPARγ, peroxisome proliferator activated receptor gamma; C/EBPα, CCAAT/enhancer binding protein alpha; aP2, adipocyte protein 2; GLUT4, glucose transporter 4; FAS, fatty acid synthase; ACC, acetyl CoA carboxylase; ApN, adiponectin; ATGL, adipose triglyceride lipase; HSL, hormone sensitive lipase; SREBP1c, sterol regulatory element-binding protein1c.
![]()
Statistical analysis
One-way analysis of variance (ANOVA) and Newman-Keuls post hoc analysis was performed to detect the significance of mean differences between groups using GraphPad Prism 7 (GraphPad Software, Inc., La Jolla, CA, USA). The α-level of P < 0.05 was considered statistically significant, and all data are expressed as means ± SEM.
Go to :
RESULTS
Inhibitory effect of SGE on lipid accumulation in 3T3-L1 adipocytes
The safety of SGE was considered by measuring cytotoxicity levels before the differentiation of preadipocytes. The cell viability of 3T3-L1 cells determined the concentrations of SGE. The cells were incubated with increasing concentrations (25 μg/mL) of SGE for 24, 48, and 72 h. There was no significant reduction in the cell viability by SGE at the range under 10 μg/mL (Fig. 1A). Therefore, the following experiments were done with a concentration of fewer than 10 μg/mL of SGE on 3T3-L1 cells. The increased lipid accumulation in adipocytes is a critical feature in adipogenesis. The contents of TG were measured to investigate whether SGE can attenuate lipid accumulation induced during 3T3-L1 differentiations. SGE significantly decreased the TG contents in the cells compared to control (Fig. 1B). Furthermore, SGE reduced the lipid droplet size and suppressed oil droplet accumulation on differentiated cells (Fig. 1C). The activity of GPDH, a representative marker for adipocyte differentiation, was significantly decreased in SGE treated cells (Fig. 1D).
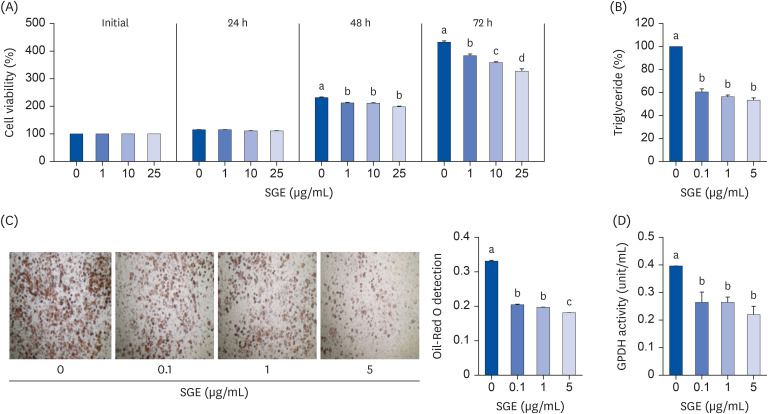 | Fig. 1Effects of SGE on (A) cell viability (B) Triglyceride contents (C) Oil-Red O staining and (D) GPDH activity in 3T3-L1 cells. Cells were treated with different concentrations of SGE. Quantitative real-time-polymerase chain reaction analysis was conducted to measure mRNA abundance. Data are expressed as relative expression to GAPDH control. Bars with different letters are significantly different (P < 0.05). Values are means ± SEM; n = 6.SGE, ethanolic extract of steamed ginger; GPDH, glycerol-3-phosphate dehydrogenase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
|
Effect of SGE on the gene expression in 3T3-L1 adipocytes
To investigate the underlying mechanisms of SGE on 3T3-L1 adipocyte differentiation, we measured the genes involved in adipogenesis and lipogenesis. The expression of PPARγ, the master transcription factor for adipocyte differentiation, was significantly and dose-dependently decreased by SGE. There was a marked reduction of C/EBPα, a well-known regulator for adipogenesis, in SGE treated 3T3-L1 cells. During adipocyte differentiation, the expressions of aP2, GLUT4, fatty acids synthase (FAS), acetyl-CoA carboxylase (ACC), adiponectin (ApN) are regulated by the actions of PPARγ and C/EBPα. The mRNA abundance of aP2, GLUT4, FAS, ACC, and ApN was significantly decreased by SGE treatment. We also measured genes for lipolysis in SGE treated mature adipocytes (Fig. 2).
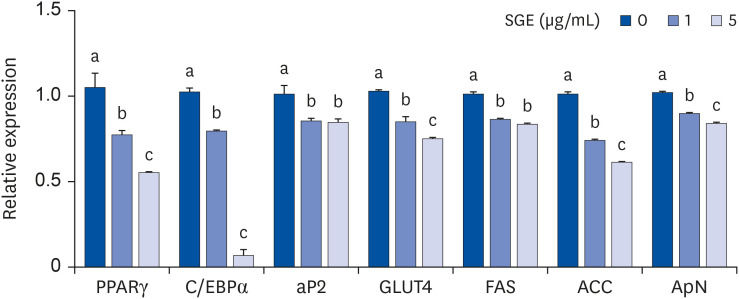 | Fig. 2Effects of SGE during differentiation of 3T3-L1 cells. Cells were treated with 1 or 5 μg/mL of SGE during differentiation. Quantitative real-time-polymerase chain reaction analysis was conducted to measure mRNA abundance. Data are expressed as relative expression to GAPDH control. Bars with different letters are significantly different (P < 0.05). Values are means ± SEM; n = 6.SGE, ethanolic extract of steamed ginger; PPARγ, peroxisome proliferator-activated receptor gamma; C/EBPα, CCAAT/enhancer binding protein alpha; aP2, adipocyte protein 2; GLUT4, glucose transporter 4; FAS, fatty acid synthase; ACC, acetyl CoA carboxylase; ApN, adiponectin; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
|
Effect of SGE on body and tissue weights and serum chemistry in HFD-induced obese mice
The inhibitory effect of SGE on adipocyte differentiation in 3T3-L1 preadipocytes suggested the anti-obesity effect of SGE. We next examined the effects of SGE on DIO mice. The HFD significantly increased body, epididymal adipose fat, and mesenteric adipose fat weight compared to ND, whereas SGE supplementation significantly attenuated the increases. The increased liver weight and the elevation of hepatic TC and TG concentrations were observed in HFD-fed groups. However, SGE supplementation (40 mg/kg or 80 mg/kg) significantly attenuated the weight gain and the hepatic TC and TG concentrations compared to those of HFD (Table 3).
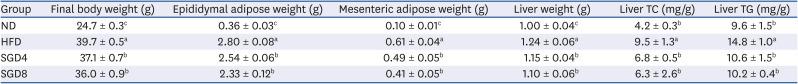
Table 3
Body weight, adipose weight, liver weight and lipid contents of C57BL/6J mice fed high-fat diet supplemented with SGE for 12 weeks
Data represent mean ± SEM (n = 10). Values with different letter in a column are significantly different (P < 0.05).
SGE, ethanolic extract of steamed ginger; TC, total cholesterol; TG, triglyceride; ND, normal diet; HFD, high-fat diet; SGD4, HFD with 40 mg/kg of SGE; SGD8, HFD with 80 mg/kg of SGE.
![]()
Serum TC and TG levels were significantly elevated in HFD compared to ND, whereas SGE significantly and dose-dependently attenuated the increases. Elevated serum glucose and insulin levels in the HFD groups, compared with the ND, were significantly (P < 0.05) decreased in SGE supplemented groups. Furthermore, the increased levels of serum adiponectin and leptin were markedly attenuated in SGD4 and SGD8 groups compared to those of HFD. Moreover, ALT and AST were significantly decreased in the serum of SGE supplemented DIO mice (Table 4).
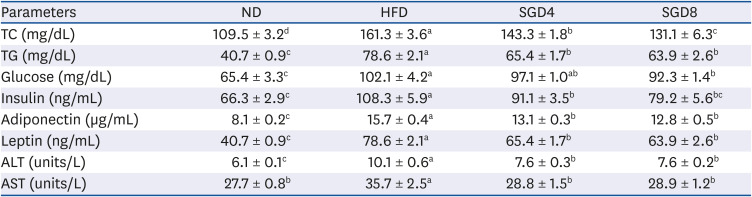
Table 4
Serum chemistry of C57BL/6J mice fed high-fat diet supplemented with SGE for 12 week
Data represent mean ± SEM (n = 10). Values with different letter in a raw are significantly different (P < 0.05).
SGE, ethanolic extract of steamed ginger; ND, normal diet; HFD, high-fat diet; SGD4, HFD with 40 mg/kg of SGE; SGD8, HFD with 80 mg/kg of SGE; TC, total cholesterol; TG, triglyceride; ALT, alanine transaminase; AST, aspartate transferase.
![]()
Effect of SGE on the epididymal adipose tissue of DIO mice
The effects of SGE on adipogenesis in the epididymal adipose tissue of DIO mice were examined by histology and micro-CT images. SGE supplementation attenuated the size and length of the adipocyte in epididymal adipose tissue compared to those of HFD (Fig. 3A and 3B). The micro-CT also showed that SGE significantly decreased the fat mass in HFD induced obesity (Fig. 3C and D).
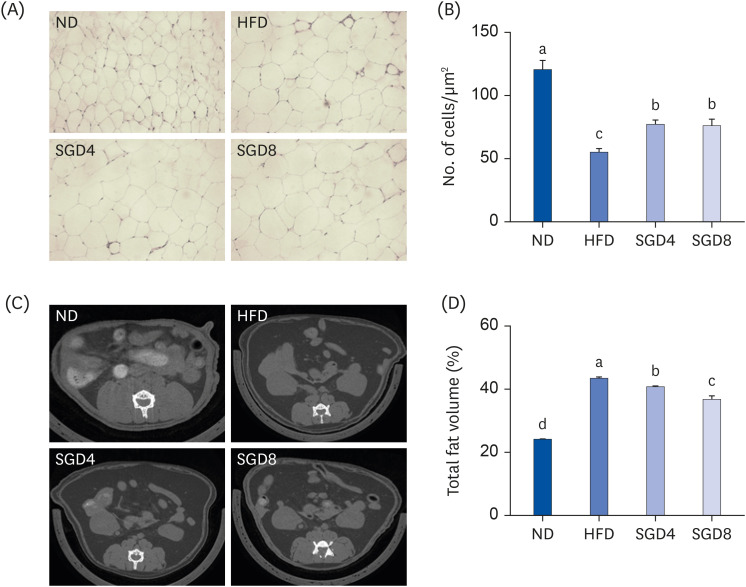 | Fig. 3Epididymal adipose tissue in SGE supplemented diet-induced obesity mice for 12 weeks. (A) Hematoxylin and eosin stained adipose tissue (B) Adipocyte number (C) The representative images of micro-CT (D) The fat volume of mice using in vivo micro-CT image analysis. Bars with different letters are significantly different (P < 0.05). Values are means ± SEM; n = 10.SGE, ethanolic extract of steamed ginger; ND, normal diet; HFD, high-fat diet; SGD4, HFD with 40 mg/kg of SGE; SGD8, HFD with 80 mg/kg of SGE; CT, computed tomography.
|
Effect of SGE on the mRNA expression in epididymal adipose tissue of DIO mice
We investigated whether the alterations in the expressions of genes involved in adipogenesis and TG synthesis are responsible for decreasing weight and adipocyte size in epididymal adipose tissue of DIO mice by SGE supplementation. Interestingly, the mRNA abundance of transcription factors for adipogenesis and lipogenesis, i.e., PPAR, C/EBPα and sterol regulatory eleme0nt-binding protein1c (SREBP1c), was significantly reduced in epididymal adipose tissue of SGE groups compared to HFD. HFD exerted the decreased expression of GLUT4 and ApN in the epididymal adipose tissue, whereas the expressions were increased in both SGD4 and SGD8 groups. Furthermore, the increased expression of aP2, FAS, ACC in the adipose tissue of HFD were significantly decreased by SGE supplementation. In contrast, the expression of genes for lipolysis, ATGL and HSL, were significantly induced in the epididymal fat by SGE supplementation (Fig. 4A and B).
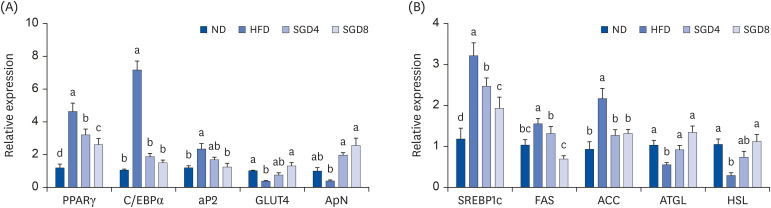 | Fig. 4Effect of SGE in the epididymal adipose tissue of mice. mRNA expression of gene involved in (A) adipogenesis and (B) lipid metabolism. Bars with different letters are significantly different (P < 0.05). Values are means ± SEM; n = 10.SGE, ethanolic extract of steamed ginger; ND, normal diet; HFD, high-fat diet, SGD4, HFD with 40 mg/kg of SGE; SGD8, HFD with 80 mg/kg of SGE; PPARγ, peroxisome proliferator-activated receptorγ; C/EBPα, CCAAT/enhancer-binding proteinα; aP2, adipocyte protein 2; GLUT4, glucose transporter type 4; ApN, adiponectin; SREBP1c, sterol regulatory element-binding protein1c; FAS, fatty acid synthase; ACC, acetyl-coA carboxylase; ATGL, adipose triglyceride lipase; HSL, hormone-sensitive lipase.
|
Go to :
DISCUSSION
Obesity and its associated metabolic diseases are the major health problem worldwide, and herbal medicines are claimed to control these diseases effectively. Ginger, one of the widely used spices and traditional medicines, is on the United States Food and Drug Administration (FDA)'s Generally Recognized as Safe (GRAS) list. The beneficial effects and mechanisms of ginger and/or its bioactive phenolic components on obesity are well-known. However, most of the studies focused on the effects of raw, dried, or frozen ginger. Ginger may undergo a heating or steaming process when used as a spice or traditional medicine. The steaming process changed the chemical profiles of root vegetables and herbs, i.e., Panax notoginseng, Codonopsis lanceolate and garlic, etc., [1718] and affected their bioactivities [19202122]. Furthermore, Cheng et al. [23] reported that the steaming process affected the chemical profile of ginger, and the steamed ginger exerted a higher antiproliferative effect than that of dried and fresh ginger in Hela cells. The effects of ginger on obesity are well-known. However, to our knowledge, the present study is the first study investigating whether steamed ginger affects adipogenesis and adiposity in both cell and epididymal adipose tissue of the DIO mouse model.
In the present study, we investigated the effect of steamed ginger on adipogenesis in adipocytes and the DIO mice model. Adipogenesis and TG accumulations in adipocytes are associated with obesity and are essential to avoid ectopic fat deposition in non-adipose tissue [30]. In obesity, adipose tissue grows by hyperplasia (cell number increase) and/or hypertrophy (cell size increase). One winning strategy for the prevention of obesity is targeting adipogenesis [31].
Consistent with our study, the bioactive component of gingers showed an inhibitory effect on adipogenesis and lipid accumulation in 3T3-L1 cells [32333435363738]. The GPDH plays a significant role in TG accumulation in adipocytes by providing the glycerol for the TG synthesis. The phytochemicals of ginger, i. e., 6-shogaol, 6-gingerol, galanolactone, and gingerenone A, exerted inhibitory effects on intracellular lipid contents and GPDH activity in 3T3-L1 cells. In the present study, SGE significantly suppressed TG content and GPDH activity in 3T3-L1 cells. Adipogenesis is strictly regulated by the master switch of crucial transcription factors that activate adipogenesis genes [3940]. PPARγ and C/EBPα, the master regulators among the cascade of adipogenic transcription factors, are involved in the final stage of adipogenesis. These transcription factors induce their downstream genes that promote preadipocyte differentiation into mature adipocytes [394041]. Activation of PPARγ by its ligands also leads to adipocyte differentiation [42]. PPARγ alone or with C/EBPα induces genes involved in lipid storage, lipogenesis, and insulin sensitivity, e.g., aP2, cluster of differentiation 36 (CD36), LPL, GLUT4, and ApN [39404142]. SREBP-1c is the key transcription factor for lipogenesis and induces enzymes involved in lipogenesis such as ACC, FAS, and stearoyl-CoA desaturase-1 (SCD-1) [43]. The transcription factor also stimulates C/EBPα and PPARγ and plays an important role in adipogenesis [41]. SGE significantly inhibited adipocyte differentiation and lipid accumulation in 3T3-L1 cells by altering adipogenesis-related genes. The expressions of PPARγ and C/EBPα significantly, and dose-dependently decreased in SGE treated 3T3-L1 cells. Several studies reported that 6-gingerol, the primary active component of ginger, significantly reduced both mRNA and protein levels of PPARγ and C/EBPα with concomitant decreased adipogenesis-related genes during differentiation. [3334]. 6-gingerol inhibited adipocyte differentiation and cytoplasmic lipid accumulation by regulating the expression of adipogenesis-related genes via activating Wnt/b-catenin signaling pathway [35], or by inhibiting Akt/GSK3β-mediated signaling that suppresses the signaling cascades of PPARγ and C/EBPs during differentiation of 3T3-L1 cells [33]. Compared to 6-gingerol, 6-shogaol, another major active component of ginger, exerted higher inhibitory effects on the adipogenic process of 3T3-L1 preadipocytes by suppressing adipogenic/lipogenic marker proteins [36]. 6-shogaol and its major metabolite, 6-paradol, showed potent activity in stimulating glucose utilization by 3T3-L1 cells by increasing 5′ adenosine monophosphate-activated protein kinase (AMPK) phosphorylation [37]. Galanolactone isolated from ginger exerted a significant reduction in fat accumulation in adipocytes with downregulation of PPARγ and C/EBPα and aP2 mRNA expression [38]. The major bioactive components of ginger are 6-gingerol and 6-shogaol [44]. SGE contained the highest contents of 6-shogaol, among other extracts (data not shown). Therefore, these active compounds present in SGE may have potent inhibitory effects on adipocyte differentiation by altering genes involved in adipogenesis and lipogenesis.
In 3T3-L1 cells, SGE exerted strong inhibitory effects on differentiation by regulating adipogenesis related gene. Therefore, we next investigated whether SGE can have protective effects on DIO mice. In the present study, HFD significantly increased body weight gain, weights of epididymal adipose tissue, mesenteric adipose tissue, and liver compared to those of ND in C57BL/6J mice. Furthermore, HFD induced the elevation of serum TC and TG levels compared to those of ND. However, SGE supplementation (40 mg/kg or 80 mg/kg) significantly attenuated the weight gains and the increased serum TC and TG levels compared to those of HFD. Consistent with our results, studies reported the hypolipidemic and hypocholesterolemic effects of ethanolic extracts of ginger by improving TC, LDL-C, and TG levels in high-fat DIO C57BL/6J mice [45], apolipoprotein E-deficient mice [46], and goldthioglucose-induced obese mice [47]. Furthermore, ethanolic extract of ginger exerted hypolipidemic and hypocholesterolemic effects in several animal models such as HFD-fed rats [48], high-fat high-carbohydrate diet-fed rats [49], streptozotocin (STZ)-induced diabetes rats [50], and cholesterol-fed rabbits [51]. The lipid metabolism in hypercholesterolemic rats was improved by aqueous extract of ginger [52] and ginger powder [53]. Several studies reported the protective effects of bioactive compounds in ginger against dyslipidemia. 6-gingerol attenuated dyslipidemia in HFD-fed mice [54], obese db/db mice [55], and high-fat-diet fed rat [56]. In the HFD-fed golden hamster, 6-gingerol supplementation decreased the plasma TC, TG, and LDL-C levels, with concomitant inhibition of hepatic lipogenic genes [57]. Gingerol and shogaol-enriched extract decreased plasma TC and liver cholesterol via alteration of genes involved in cholesterol absorption and bile acid secretion in hamsters fed a high-cholesterol diet [58].
In the present study, SGE supplementation exerted hypoglycemic effects in DIO mice model. Several studies have reported the hypoglycemic properties of ginger and its bioactive compounds in animal models. Ethanolic extract of ginger attenuated hyperinsulinemia induced by liquid fructose in rats [59] and HFD-fed rats [48]. Furthermore, improved glucose and lipid profile by ethanolic extract of ginger in HFD-fed rats was associated with the alteration of hepatic genes involved in lipid metabolism [60]. Aqueous extract of ginger (500 mg/kg) exerted antihyperglycemic effects in STZ-induced diabetic rats [6162] and alloxan-induced diabetes rats [63]. Gingerol with HFD significantly decreased glucose, leptin, insulin, lipids levels in plasma, and liver lipids compared to control [56]. Wei et al. [37] compared the hypoglycemic effects of different active components of ginger, namely gingerols, shogaols, zingerone, and paradols. Among the active components, 6-paradol, the major metabolite of 6-shogaol, significantly reduced blood glucose and cholesterol in high-fat-diet-fed mice. These active compounds may responsible for the protective effect of SGE on increased serum lipid chemistry and hyperglycemia induced by a HFD.
Both adipocyte hypertrophy and hyperplasia contribute to adipose tissue expansion during the HFD challenge. In epididymal adipose tissue, hypertrophy is the prominent phenomenon contributing to adipose tissue expansion during the early stages of a HFD (within 1 month), and adipogenesis is apparent at the late stages of HFD [64]. In obese individuals and rodents, hypertrophy is closely related to the pathological expansion of adipose tissue. In the present study, SGE significantly reduced the weight and adipocyte size in epididymal adipose tissue than those of HFD. Similar to our results, ginger decreased adipocyte size and increased adipocyte number followed by suppressed macrophage infiltration and macrophage-associated cytokine in adipose fats of fructose-induced hyperinsulinemia in rats [59].
Differentiated adipocytes are post-mitotic; therefore, hyperplasia represents an increase in de novo adipocyte formation (adipogenesis). Adipocyte hypertrophy is closely linked to adipose dysfunction: this pathological expansion of white adipose tissue (WAT) is a significant component of the metabolic syndrome in obese individuals [264]. Therefore, we investigated whether the alterations in the expressions of genes involved in adipogenesis and TG synthesis are responsible for decreasing weight and adipocyte size in epididymal adipose tissue by SGE in DIO mice. The mRNA abundance of the adipogenic and lipogenic transcription factors and their downstream genes were significantly increased in epididymal adipose tissue in DIO mice. Interestingly, SGE supplementations attenuated the increases of epididymal gene expressions for adipogenesis and lipogenesis, i. e. PPARγ, C/EBPα, SREBP1-c, FAS, and ACC in DIO mice. Consistent with our findings, gingerenone A, a polyphenol in ginger, decreased the size of adipocyte in HFD-fed mice and reduced the expression of SREBP1 and FAS in epididymal adipose tissue [32]. The hot water extract of ginger ameliorated obesity and obesity-associated inflammation by lowering the mRNA levels of adipogenic genes and pro-inflammatory cytokines in white adipose tissue of rats fed a HFD [65]. Wang et al. [59] reported the ethanolic extract of ginger suppressed the expression of macrophage filtration marker but did not alter the adipogenic and lipogenic in adipose tissue of liquid fructose-fed rats. These results indicate extraction method may affect the different underlying mechanisms of the protective effect of ginger against obesity.
The lipolysis, the TG breakdown process, is essential in lipid metabolism and frequently altered with obesity [66]. In adipose tissue, the activity of lipases such as lipoprotein lipase (LPL), ATGL, and HSL are known to increase in obesity [6768]. Adipocyte lipolysis is a complex process mainly controlled by ATGL and HSL [68]. The mRNA abundance of ATGL and HSL in epididymal adipose tissue was significantly increased in SGE groups compared to the HFD group. Gingerenone A supplementation showed higher expression of ATGL and HSL in HFD-fed mice [32].
In the present study, the steamed ginger inhibited adipogenesis by regulating genes involved in adipocyte differentiation and lipogenesis in 3T3-L1 cells. Furthermore, steamed ginger exerted protective effects against adiposity in the epididymal adipose tissue of DIO mice by attenuating adipogenic and lipogenic genes. Further study is warranted to investigate which components of steamed ginger are responsible for these effects. In conclusion, the present study provides and supports that steamed ginger protects against high-fat DIO and metabolic disorders.
Go to :
 XML Download
XML Download