

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Atherosclerosis is a sustained inflammatory disease in which the inside of arteries narrows due to the build-up of an atheromatous plaque. Several studies have shown that systemic inflammation and pro-inflammatory cytokines characterize early atherogenesis [1]. Oxidized low-density lipoproteins (OxLDLs) are reported to increase a series of pro-inflammatory reactions via different mediators (tumor necrosis factor [TNF]-α, interleukin [IL]-6, high mobility group box [HMGB]-1, etc.) [2]. In the early period of atherogenesis, OxLDL prompts expression of the chemokine (C-X3-C-motif) ligand 1 (CX3CL1) in vascular smooth muscle and endothelial cells (ECs). CX3CL1 is produced as a membrane-bound long protein, with its chemokine domain being situated on a mucin-like stalk. Cleavage at the base of this stalk generates a TNF-α-converting enzyme, a soluble chemokine with roles as a strong chemoattractant for T cells and monocytes, while the cell-bound chemokine enhances the strong adhesion of leukocytes to activated ECs [3]. Furthermore, high levels of inflammatory factors result in monocyte adhesion and differentiation into macrophages and increase the production of several adhesion molecules, containing vascular cell adhesion molecule-1 (VCAM-1), intracellular adhesion molecule-1 (ICAM-1), and E-selectin [4]. The activated macrophages migrate into the intima, becoming lipid-laden and transforming into foam cells. Some studies have revealed non-traditional roles for cathepsins in extracellular matrix remodeling. Cysteine protease cathepsins have a critical role in extracellular matrix remodeling and have been involved in the development and progression of atherosclerosis-based cardiovascular diseases [5]. The most potent of these cathepsins is cathepsin S, an elastolytic enzyme expressed predominantly in macrophages and inflamed medial smooth muscle cells [6]. Thus, natural products that control vascular inflammatory mediators may be efficacious in atherosclerosis. Curcuma zedoaria R. (Zingiberaceae) is a traditional Korean medicinal plant used to treat symptoms such as fatigue and weakness after illness. The major chemical constituents of C. zedoaria include curcumenol and zedoarofuran [7], curcumenone [8], bisabolone-9-one, turmeronol B, bisdemethoxycurcumin, desmethoxycurcumin, and curcumin [9]. This plant has been reported to exhibit anti-oxidant [10], cancer [111213], inflammatory [1415] and fungal effects [16], to reduce drunkenness [17], and to exert analgesic [18] and antitrypanosomal activities [19]. However, the relationship between a C. zedoaria extract (CzE) and atherosclerosis has not been fully elucidated. A previous report showing an anti-inflammatory activity of C. zedoaria encouraged us to study the potential anti-atherogenic effects of CzE. Currently, an apolipoprotein E-deficient (ApoE−/−) mouse model is the most frequently used animal model in the study of atherosclerosis. The ApoE−/− mouse model's main advantage is that the rate of atherogenesis can be markedly hastened by a high-fat, high-cholesterol diet (HCD), which results in significant increases in plasma lipid levels. Provision of an HCD is the best method for inducing systematic lesions in ApoE−/− mice. Thus, we investigated the effects of CzE on atherosclerosis and associated inflammatory mediators in ApoE−/− mice fed an HCD.
Go to : 
MATERIALS AND METHODS
Plant material
Dried C. zedoaria material was obtained from a commercial supplier (Kwangmyung-Dang, Ulsan, Korea). A voucher specimen was deposited in the Herbal Medicine Resources Research Center (Korea Institute of Oriental Medicine). The rhizome (3.0 kg) was extracted by using distilled water under reflux for 3 h. The obtained product was evaporated to dryness under vacuum and finally freeze-dried, producing the CzE (103.3 g). The lyophilized extract used in the animal experiments was stored at −20°C. The chemical components contained in CzE were zedoarondiol, isozedoarondiol, procurcumadiol, procurcumenol, and curcumenol. An high-pressure liquid chromatography analysis was performed using a Shimadzu system with ultraviolet detection at 265 nm and a LC-18 column (150 mm × 4.6 mm, 5 mm) from Phenomenex. The solvent system used was water:acetonitrile (95:5; v/v -> 30:70 -> 1:10) injected at a flow rate of 1 mL/min and an injection volume of 10 µL.
Animals
ApoE−/− mice [20] were acquired from Central Laboratory Animal Inc. (Seoul, Korea) and housed in cages (3 mice per cage) under specific pathogen-free conditions. All animal protocols were in accord with those of the Committee of Animal Care and Experiments of Chungnam National University, Korea (CNUH-015-A0017). Thirty-two male ApoE−/− mice were randomly separated into 4 dietary groups: normal diet (ND), n = 6; HCD, n = 10; HCD compensated with CzE, n = 10; and HCD supplemented with simvastatin, n = 6. The HCD included 15% fat (7.5% lard and 7.5% cacao butter), 1.25% cholesterol, and 0.5% cholic acid. The mice were given water and chow ad libitum. Mouse body weight was measured every 3 weeks for 12 weeks. After 12 weeks, parameters were analyzed by western blot analysis, immunohistochemistry, and bioluminescence imaging (PerkinElmer Inc., Waltham, MA, USA).
Histological examinations
The size of the aortic sinus lesions was quantified as described in a previous paper [21], with a slight modification. The heart and aorta were perfused with phosphate-buffered saline for 10 minutes and then with 3.5% paraformaldehyde for 5 minutes. The heart was then rapidly removed, fixed in 10% buffered neutral formalin for 24 h, and washed in tap water to remove the formalin. Atherosclerotic plaque samples were stained with Oil Red O. Six lesion areas were compared by using computer-supported morphometry (Image-Pro Plus; Media Cybernetics, Rockville, MD, USA) at 30-μm intervals, and the average lesion size was estimated (n = 22). To detect the cytokine level, frozen sections were stained individually with 1:200 dilutions of anti-TNF-α, CX3CL1, or HMGB-1 antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Harris hematoxylin was used as the counterstain.
Serum parameter analysis
Analysis of the serum parameters was performed according to methods described in a previous study [22]. Briefly, serum levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), triglyceride (TG), total cholesterol (TC), glucose, and high-density lipoproteins (HDLs) were determined using a clinical chemistry analyzer system (Dri-Chem 700i; Fujifilm, Tokyo, Japan). The serum low-density lipoprotein (LDL) content was determined using LDL Assay kits (KOMA Biotech, Seoul, Korea).
Ex vivo fluorescence imaging
Ex vivo fluorescence was evaluated by applying the method reported previously [23]. Briefly, mice were anesthetized with isoflurane/oxygen/nitrogen, and the aorta, liver, and lung were excised and subsequently underwent fluorescent signal analysis via imaging performed using an in vivo imaging system (IVIS Lumina XR; PerkinElmer Inc.). The fluorescent intensity of ProSense 750 (PerkinElmer Inc.) in the selected organ was analyzed.
Western blot analysis
Proteins obtained from mouse aortas were ultrasonically homogenized in lysis buffer that included protease inhibitors (Roche Diagnostics, Indianapolis, IN, USA). Protein concentrations were analyzed by using a Pierce BCA Assay Kit (Thermo Fisher Scientific, Rockford, IL, USA). Western blot analysis was undertaken as previously described [24]. The blotted membranes were developed by using an enhanced chemiluminescence system (Thermo Scientific), and the blots were identified using an LAS-4000 mini luminescent image analyzer (GE Healthcare, London, UK). Quantitative band density was determined by using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All measurements were obtained in triplicate, and the data are presented as mean ± standard deviation values. Analysis of variance in conjunction with Tukey's test was used to analyze group differences, and a P-value < 0.05 was considered to indicate statistical significance.
Go to :
RESULTS
CzE regulates lipid profile in blood of ApoE−/− mice
The body weights of ApoE−/− mice fed the ND and those in the experimental diet groups were recorded periodically to observe changes related to CzE treatment. As listed in Table 1, the average body weights of the ND- and HCD-fed mice were significantly different. A significant reduction in body weight was observed in ApoE−/− mice when treated with HCD and CzE. In addition, as shown in Table 2, HCD-fed ApoE−/− mice had significantly elevated serum total TG, TC, glucose, and LDL levels compared to those in ND-fed ApoE−/− mice. Thus, treatment with CzE remarkably lowered both TG and HDL levels.

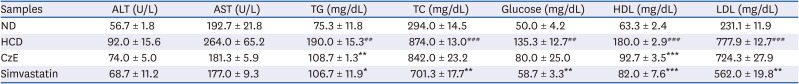
Table 1
Body weight changes in ApoE−/− mice after 12 weeks of CzE administration
Data are presented as mean ± standard error of mean values (n = 6, each group). ApoE−/− mice treated with simvastatin were considered positive controls.
ND, normal diet; HCD, high-cholesterol diet; CzE, Curcuma zedoaria extract (HCD with CzE treatment group), Simvastatin, HCD with simvastatin treatment group.
**P < 0.01 vs. HCD group; ##P < 0.01 vs. ND group.
![]()
Table 2
Effect of CzE on serum lipid profile levels in ApoE−/− mice
Data are presented as mean ± standard error of mean values (n = 3, each group). ApoE−/− mice treated with simvastatin were considered positive controls.
ND, normal diet; HCD, high-cholesterol diet; CzE, Curcuma zedoaria extract (HCD with CzE treatment group); Simvastatin, HCD with simvastatin treatment group; ALT, alanine aminotransferase; AST, aspartate aminotransferase; TG, triglyceride; TC, total cholesterol; HDL, high-density lipoprotein; LDL, low-density lipoprotein.
*P < 0.05, **P < 0.01, ***P < 0.001 vs. HCD group; #P < 0.01, ##P < 0.001 vs. ND group.
![]()
Anti-atherogenic effect of CzE on ApoE−/− mice
After 12 weeks of CzE administration, the effect of CzE on the formation of atherosclerotic lesions in the aortic sinuses of ApoE−/− mice was assessed. Fig. 1 shows sections of the aortic roots from ApoE−/− mice in the control, HCD, and HCD with CzE groups. Aortic roots of the control group mice showed no fatty streak lesions, whereas mice fed the HCD for 12 weeks showed considerable fatty streak lesions on the intima of the aortic sinuses (Fig. 1B). However, in the HCD with CzE group mice, the diet-induced aortic injury area was 41.6% smaller than that of the HCD-fed mice (5.50 ± 0.87 [× 106] μm2 vs. 3.21 ± 0.18 [× 106] μm2, respectively P < 0.01) (Fig. 1C and D).
 | Fig. 1Representative images of Oil Red O-stained gross atherosclerotic lesions in HCD-fed and control mice (A-C). Typical cross-sections of aortic valves (× 100) in hearts from control (ND) (A), HCD (B), and CzE-treated HCD (C) mice. Treatment with CzE (100 mg·kg−1·d−1) for 12 weeks affects atherosclerosis in ApoE−/− mice. (D) Mean atherosclerotic lesion area in the aortic sinus was determined; data are expressed as mean ± SD values (n = 6 for each group).ND, normal diet; HCD, high-cholesterol diet; CzE, Curcuma zedoaria extract; ApoE−/−, apolipoprotein E-deficient.
**P < 0.01 vs. HCD group.
|
Expressions of TNF-α, CX3CL1, and HMGB-1 induced by CzE in normal intima and atherosclerotic lesions of ApoE−/− mice
Frozen aortic sinus sections were stained with antibodies for TNF-α, CX3CL1, and HMGB-1 in accordance with the methods described previously [25]. Consistent with our western blot analysis results, ApoE−/− mice demonstrated reduced expression of the inflammatory markers TNF-α, CX3CL1, and HMGB-1 (P < 0.05) compared with those of HCD-fed ApoE−/− mice. Both TNF-α and CX3CL1 were highly expressed in the HCD-fed group, but that increased expression was remarkably suppressed by the combined HCD–CzE treatment (Fig. 2). Compared with wild-type mice, the vascular HMGB-1 expression level was notably amplified in ApoE−/− mice (P < 0.01) (Fig. 2). However, this was reversed by CzE treatment. Additionally, the western blot analysis results showed a pattern similar to that for atherosclerotic lesions (Fig. 4). In addition, TNF-α, and CX3CL1 were expressed not only in ECs, but also in the intima. However, HMGB-1 was detected primarily in the intima of the atherosclerotic lesions (Fig. 2).
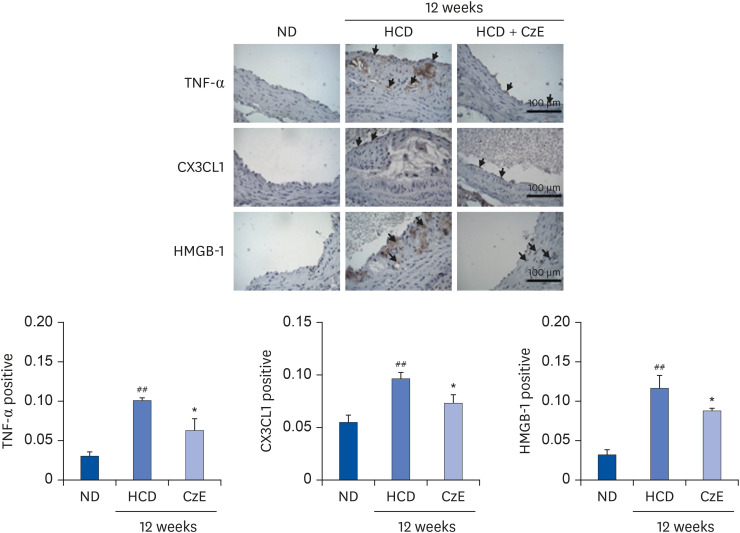 | Fig. 2Immunohistochemical identification of inflammatory molecules in ApoE−/− mouse aortas. ApoE−/− mice were fed an HCD for 12 weeks, after which, inflammatory molecules in plaques in the aortic sinuses were assessed histologically and immunohistochemically. In the control aorta, TNF-α, CX3CL1, and HMGB-1 showed negligible staining (brown staining). HCD-fed ApoE−/− mice showed over-expression of TNF-α, CX3CL1, and HMGB-1 not only in endothelial cells, including in some intimal areas, but also in smooth muscle cells (dark-brown staining: black arrows). However, in HCD–CzE-fed ApoE−/− mice, the expressions of these inflammatory molecules were significantly inhibited. Arrowheads indicate TNF-α-, CX3CL1-, or HMGB-1-positive cells. All results are shown as mean ± SD values.ND, normal diet; HCD, high-cholesterol diet; CzE, Curcuma zedoaria extract; ApoE−/−, apolipoprotein E-deficient; TNF-α, tumor necrosis factor-α; CX3CL1, chemokine (C-X3-C-motif) ligand 1; HMGB-1, high mobility group box-1.
*P < 0.05 vs. HCD group; ##P < 0.01 vs. ND group.
|
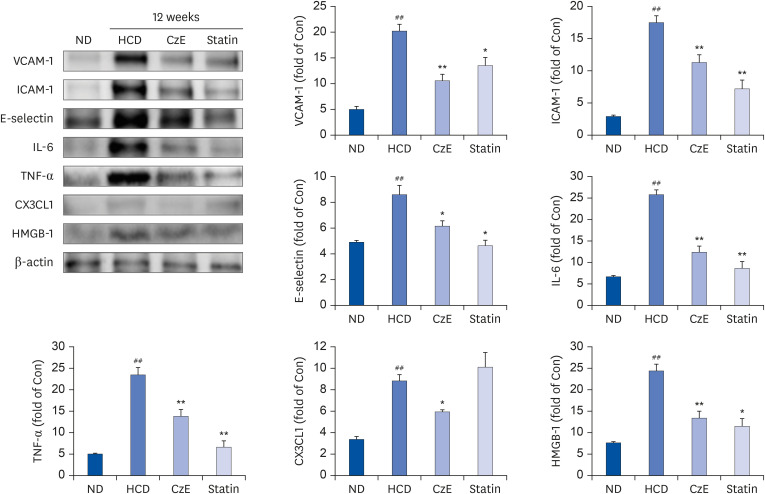 | Fig. 4Effect of CzE on adhesion molecules and inflammatory mediators in the aortas of ApoE−/− mice. The data were collected from ND, HFD with CzE (100 mg/kg), and HFD with simvastatin (10 mg/kg) fed mice (treated orally for 12 weeks). Representative photomicrographs of western blot results and densitometric analysis show the relative protein expression levels of adhesion molecules (ICAM-1, VCAM-1, and E-selectin) and inflammatory mediators (TNF-α, IL-6, CX3CL1, and HMGB-1), normalized to β-actin. Values are mean ± SD.ND, normal diet; HCD, high-cholesterol diet; CzE, Curcuma zedoaria extract; ApoE−/−, apolipoprotein E-deficient; ICAM-1, intracellular adhesion molecule-1; VCAM-1, vascular cell adhesion molecule-1; TNF-α, tumor necrosis factor-α; IL-6, interleukin-6; CX3CL1, chemokine (C-X3-C-motif) ligand 1; HMGB-1, high mobility group box-1.
*P < 0.05, **P < 0.01 vs. HCD group; ##P < 0.01 vs. ND group.
|
CzE treatment decreases arterial cathepsin activity in HCD-fed ApoE−/− mice
The HCD-fed group exhibited an increased proteolytic fluorescence level in the carotid arteries compared with that in control mice (P < 0.01); however, CzE reduced the cathepsin activity in the aorta and carotid arteries (Fig. 3). The ApoE−/− mice fed the HCD with simvastatin added showed markedly reduced cathepsin activity in the aorta (Fig. 3). We also detected a significant reduction in ProSense 750 signals in mice fed the HFD with CzE (P < 0.01) compared to that in the HFD-fed group (Fig. 3).
 | Fig. 3Visualization of cathepsin activity in atherosclerotic mouse aortas. Visible light images show that the aortas of ApoE−/− mice appear normal, whereas those of ApoE−/− mice fed an HCD exhibit increased proteolytic fluorescence in the carotid arteries. The fluorescent density distributed from the ascending aorta to the infrarenal aorta in each group indicates the location of cathepsin activity. In the ApoE−/− mice fed the HCD with CzE diet (100 mg·kg−1·d−1), the inflammatory cathepsin expression level was significantly inhibited. The presence of internally quenched fluorogenic peptides containing Abz-Dnp, which are activatable by pan-cathepsin, confirmed the inhibition of inflammatory cardiovascular disease by CzE treatment in ApoE−/− mice. Values presented are mean ± SD.ND, normal diet; HCD, high-cholesterol diet; CzE, Curcuma zedoaria extract; ApoE−/−, apolipoprotein E-deficient.
**P < 0.01 vs. HCD group; ##P < 0.01 vs. ND group.
|
Effect of CzE on vascular inflammatory molecules in HCD-induced ApoE−/− mice
We also compared the expressions of inflammatory and adhesion molecules in the atherosclerotic aorta of HCD-fed ApoE−/− mice by examining the western blot results. The expression levels of VCAM-1, ICAM-1, and E-selectin in HCD-fed mice were notably increased compared with those of control mice. In contrast, the CzE- and simvastatin-treated mice had expression levels of VCAM-1, ICAM-1, and E-selectin lower than those of the HFD-fed mice (Fig. 4). In addition, we determined the levels of the inflammatory molecules TNF-α, IL-6, CX3CL1, and HMGB-1, which induce atherosclerotic inflammation in the aorta tissue. Administration of CzE in HFD-fed ApoE−/− mice markedly decreased the expressions of TNF-α, IL-6, CX3CL1, and HMGB-1 in the aorta (Fig. 4). These results suggest that CzE may inhibit the expression of inflammatory and adhesion molecules in the aorta.
Go to :
DISCUSSION
C. zedoaria was shown to be an effective anti-inflammatory agent in several earlier studies. In the present study, we established that CzE, a water extract of C. zedoaria in which sesquiterpenes and polyphenols are the main constituents, exerts anti-atherogenic effects in an ApoE−/− mouse model of atherosclerosis. CzE (100 mg/kg) notably decreased atherosclerotic lesion formation in the aortic sinus of ApoE−/− mice, and inhibited expression of the inflammatory mediators TNF-α, IL-6, and HMGB-1, and the chemokine CX3CL1. The expressions of the adhesion molecules VCAM-1, ICAM-1, and E-selectin in the aortic sinus of CzE-fed ApoE−/− mice were significantly decreased from those of HCD-fed ApoE−/− mice. The reduction of adhesion molecule expressions was accompanied by down-regulation of cysteine protease cathepsin activity in the aorta of ApoE−/− mice. In this study, the body weight of the CzE mice was lower than that of the HCD-fed ApoE−/− mice (Table 1). Serum analysis showed that ALT and AST levels tended to be lower in CzE-treated mice but the differences were not significant; in contrast, total TG and HDL levels were significantly improved (Table 2). Furthermore, we observed that the 12-week HCD induced lesion formation in the aorta of ApoE−/− mice, but the HCD supplemented with CzE resulted in a remarkable reduction in lesion area, similar to that induced by the commercially available anti-lipid-lowering simvastatin administered in ApoE−/− mice fed an HCD (data not shown). TNF-α, a pro-inflammatory cytokine, has been shown to play a significant modulatory role in the chronic inflammation that characterizes atherosclerosis [26]; moreover, TNF-α elevates matrix degradation, thus promoting an influx of inflammatory cells into the vessel wall. TNF-α also up-regulates the expression of cell adhesion molecules (ICAM-1, VCAM-1, and E-selectin) and the chemokine CX3CL1. The expressions of inflammatory cytokines were significantly induced by the 12-week HCD treatment, whereas treatment with HCD and CzE diminished the expression levels of TNF-α, ICAM-1, VCAM-1, E-selectin, and CX3CL1. In addition, CzE significantly inhibited the expression of HMGB-1 (a ubiquitous nuclear protein). HMGB-1 has been described as an effective mediator of tissue remodeling in experimental atherosclerosis, and there is a significant association between HMGB-1 and coronary plaque composition [27]. In this study, immunohistochemical staining and western blot results showed that the high expression levels of HMGB-1 in HCD-fed ApoE−/− mice were decreased significantly by the inclusion of CzE in the feed. In nonatherosclerotic arteries, the expression of many cathepsins, including cathepsins F, S, and K, is negligible [5], and the expression level is also low in the vessel walls of healthy arteries [28]. In atherosclerotic lesions, however, cathepsin activity is enhanced. Among cathepsins B, L, and S, which are expressed in the intima of atherosclerotic lesions, cathepsin S is the most strongly expressed in medial smooth muscle cells [5]. Thus, cathepsin activity is an important indicator of atherosclerosis. We conclude that CzE can inhibit cathepsin activity in atherogenic aortic plaques. Further, our results demonstrate that CzE (100 mg/kg) suppresses vascular inflammation and atherosclerosis in ApoE−/− mice, presumably through the down-regulation of TNF-α, IL-6, HMGB-1, and CX3CL1 in atherosclerotic plaques in conjunction with the down-regulation of adhesion molecules VCAM-1, ICAM-1, and E-selectin. Therefore, the reducing effect of CzE on potential inflammatory cytokines could be an important mechanism contributing to reducing the risks of vascular inflammation and atherosclerosis.
Go to :
 XML Download
XML Download