

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The importance of diet for improving health and preventing chronic disease is widely recognized [1]. Notably, there is an increasing interest in studying the mechanism of action involved in the effects of food constituents, such as phytochemicals, in biological systems. However, because individual responses (e.g., bioavailability) to the diet are different, identifying the link between food intake and physiological outcome is not sufficient to solve the complex characteristics of diet-health associations [2].
To understand the health effects of certain foods, scientists have utilized new biomarkers to assess dietary intake and its biological effects [3]. Advances in analytical and data processing technologies have made it possible to use the metabolites present in biological fluids, such as blood and urine, as biomarkers of intake and exposure. Exposure biomarkers of various foods have been used to support associations between dietary exposure and disease risk [45].
Soy foods, the primary source of isoflavone exposure, are commonly consumed in their fermented form in Korea. Isoflavone patterns in soy foods differ based on the method of processing, such as fermentation. An accurate determination of dietary isoflavone intake is limited by the absence of a common database establishing the isoflavone composition of various food sources [6]. Additionally, although circulating and excreted levels of isoflavones have been reported to vary between individuals, this has not been studied extensively. Therefore, this review aims to: 1) summarize factors affecting the bioavailability of soy isoflavone and recent studies on its health effects, and 2) suggest a new research strategy for identifying an exposure biomarker of soy isoflavone intake by providing an overview of the recent development of exposure biomarkers.
Go to : 
EXPOSURE BIOMARKERS FOR VALIDATION OF FOOD INTAKE
Classification of biomarkers
Biomarkers provide objective measurements used to assess the exposure, effects, or susceptibility of humans or animals [3]. Gao et al. [3] suggested the following principles for the classification of dietary and health biomarkers based on their intended use: 1) exposure and intake biomarkers represent food intake, food compounds or nutrients, and dietary patterns in biological fluids; 2) effect (efficacy) biomarkers monitor changes in biochemical, physiological, or psychological states as a response to nutritional exposures; 3) Susceptibility (health state) biomarkers can be expressed as personal resilience, and are linked to various host factor-related risks based on their food and nutrition status. Among these 3 groups, exposure biomarkers are essential for linking dietary compounds to health outcomes and provide early signals of biological effects [7]. To apply biomarkers to future nutrition research, we should focus on the development of appropriate biomarkers by understanding the bioavailability and mode of action of bioactive food components, as well as conducting extensive population studies [7].
Research on exposure biomarkers
Biomarkers of food intake are mainly small molecules derived from the digestion and biotransformation of either the food itself, or of certain food-derived compounds [8]. Advances in metabolomics and statistical techniques have allowed exposure biomarkers to be used as objective indicators of specific food intake. For example, intervention studies have identified biomarkers of specific foods of interest, including coffee [9], fish and meat [10], cruciferous vegetables [11], and wine [12]. Exposure biomarkers have also been identified in cohort studies and correlated with a self-reported intake of meat [13], whole grain bread [1415] and a vegetarian diet [16]. Exposure biomarkers can provide evidence for associations between food intake and potential health effects.
Shi et al. [4] reported that a principal component analysis-derived metabolite pattern was correlated with fish intake (ρ = 0.37; P < 0.001), but was not associated with type 2 diabetes (T2D) risk. However, after removing the counteractive effects of co-exposure to persistent organic pollutants present in fish, the study showed that fish intake in fact lowered the risk of T2D (OR, 0.75; 95% CI, 0.54–1.02; P = 0.07) [4]. Metabolite markers of coffee exposure have been suggested to be strong predictors of liver cancer and liver disease mortality [5]. Trigonelline, a known coffee biomarker, was inversely associated with liver disease mortality. In contrast, 2 bile acids, namely glycochenodeoxycholic acid and glycocholic acid, were inversely associated with coffee intake but positively associated with both liver cancer and liver disease mortality. In a study that identified metabolites associated with high adherence to the Alternative Healthy Eating Index, circulating metabolites such as specific fatty acids were inversely correlated to the risk of cardiovascular disease (CVD) [17].
Go to :
SOY ISOFLAVONE METABOLISM
Soybean isoflavones consist of 3 aglycones (daidzein, genistein, and glycitein), 3 simple glucosides (daidzin, genistin, and glycitin), and 6 acylated glucosides (Fig. 1). The compositions of the acyl groups vary depending on the processing method, such as heating or fermentation [18]. Therefore, the absorption of soy isoflavones varies significantly between subjects or the provided food matrix [1819].
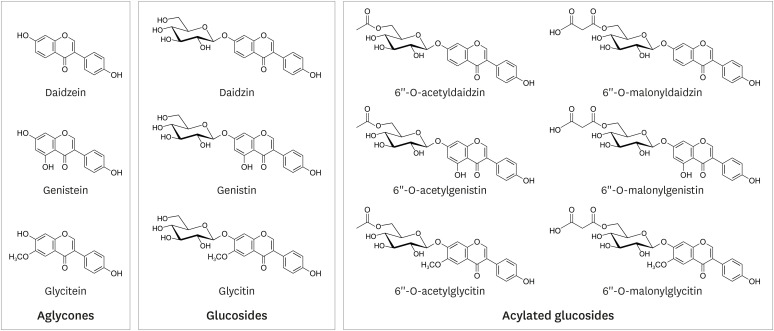
Isoflavone glucosides are not absorbed intact across the enterocyte, as they must first be deglycosylated [20]. Deglycosylation of isoflavone glucoside is carried out by the enterocyte brush boundary enzyme, lactase phlorizin hydrolase, before being absorbed into the small intestine, as shown by the appearance of a weak plasma peak after approximately 1 h [21]. Aglycones are conjugated in the intestine and liver during first-pass absorption and are then predominantly circulated in the plasma and urine as glucuronide conjugates, or to a lesser extent, sulfate conjugates [2022].
Isoflavone glucosides are hydrolyzed and metabolized by microflora in the colon. These microflora may also further degrade the isoflavone and its conjugates to reductive metabolites and ring-fission products [23]. For example, daidzein is metabolized by microbes into O-desmethylangolensin and equol. Dietary isoflavones are almost entirely conjugated with sulfates and glucuronides after intestinal absorption, whereas unconjugated isoflavones, the biologically active form, make up only 1–5% of the total isoflavone in the blood [24]. Recently, several studies have suggested that the conjugates may either have biological activity themselves [25] or serve as excellent sources of biologically active compounds within target cells through deconjugation [2627].
In order to establish conclusive evidence for the health benefits of isoflavones, it is essential to accurately determine the type and bioavailability of isoflavone in soy foods. Generally, released aglycone was quantified by the presence of glucuronidase and sulfatase enzymes in plasma and urine samples, rather than the presence of metabolites. Although this technique provided valuable insights, the results it produced were indirect and inaccurate [28].
Go to :
FACTORS AFFECTING THE BIOAVAILABILITY OF ISOFLAVONES
The bioavailability of dietary foods is affected by both food-related and human biological factors [28]. Food matrices, interactions with other compounds, and chemical structures all directly influence bioavailability. Following ingestion, bioavailability is affected by host-related factors including intestinal factors, such as enzyme activity or intestinal microflora, and systemic factors, such as gender, age, genetics, and physical condition [28].
Isoflavones have different intestinal absorption rates based on their chemical structure. In an animal study, acylated glucosides, such as malonyl glucoside, were hydrolyzed only in colon homogenates with high bacterial concentrations, whereas hydroxylation of simple glucoside occurred in jejunum, ileum, and colon homogenates [29]. This study also reported that although aglycone and simple glucosides are absorbed faster than malonyl glucoside, the absorption of malonyl-rich isoflavone in the colon contributes to a significant increase in plasma levels [29]. This result suggests that the gut microbiome produces variability of isoflavone absorption rates in the colon among different individuals.
In addition, the fermentation process can cause structural changes in soy isoflavone, which consequently affects its bioavailability and possibly optimizes its biological effect [22]. These results may help explain the discrepancies between clinical findings regarding soy foods in Asia versus the West. However, few studies have been conducted to develop biomarkers for fermented soybean (FS) intake.
Here, we outline the effects of fermentation on the bioavailability of soy isoflavone from 3 human clinical studies, as shown in Table 1. In the first study, Okabe et al. [30] reported that the isoflavone of aglycon-rich FS powder was absorbed faster and in higher amounts than those of glucoside-rich non-fermented soybean (NFS) powder in postmenopausal women. The serum concentrations of total isoflavone (calculated as the sum of genistein, daidzein, and glycitein) 1–4 h after oral consumption was significantly higher in the aglycone-rich FS than the glucoside-rich NFS group; however, there was no difference between the 2 groups 6–24 h after administration. The total isoflavone concentration in urine samples was significantly increased after intake of FS compared to that after intake of NFS in samples collected at 2–4 h and 24–48 h after administration. Genistein, daidzein, and glycitein had trends similar to that of total isoflavone in blood and urine samples, but the gut-mediated metabolite, equol, did not. A similar study by Kano et al. [31] also investigated the effects of fermentation on bioavailability of isoflavones after ingesting soymilk. The initial absorption rate of total isoflavone was greater after ingesting FS milk than after ingesting NFS milk. However, concentrations of isoflavone did not differ between the 2 kinds of milk in both blood and urine after 6–24 h and 12–48 h, respectively [31]. A third study revealed that the consumption of FS milk did not enhance urinary isoflavone excretion at any isoflavone dosage (from low to high) on any of the 3 collection days (day 4, 13, and 14) over a period of 2 weeks [32].
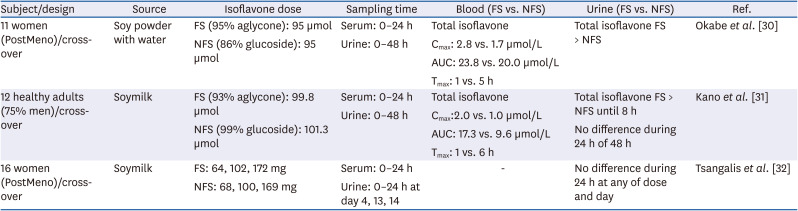
Table 1
Studies on the bioavailability of isoflavone after intakes of FS and NFS
| Subject/design | Source | Isoflavone dose | Sampling time | Blood (FS vs. NFS) | Urine (FS vs. NFS) | Ref. |
|---|---|---|---|---|---|---|
| 11 women (PostMeno)/cross-over | Soy powder with water | FS (95% aglycone): 95 µmol | Serum: 0–24 h | Total isoflavone | Total isoflavone FS > NFS | Okabe et al. [30] |
| NFS (86% glucoside): 95 µmol | Urine: 0–48 h | Cmax: 2.8 vs. 1.7 µmol/L | ||||
| AUC: 23.8 vs. 20.0 µmol/L | ||||||
| Tmax: 1 vs. 5 h | ||||||
| 12 healthy adults (75% men)/cross-over | Soymilk | FS (93% aglycone): 99.8 µmol | Serum: 0–24 h | Total isoflavone | Total isoflavone FS > NFS until 8 h | Kano et al. [31] |
| NFS (99% glucoside): 101.3 µmol | Urine: 0–48 h | Cmax:2.0 vs. 1.0 µmol/L | No difference during 24 h of 48 h | |||
| AUC: 17.3 vs. 9.6 µmol/L | ||||||
| Tmax: 1 vs. 6 h | ||||||
| 16 women (PostMeno)/cross-over | Soymilk | FS: 64, 102, 172 mg | Serum: 0–24 h | - | No difference during 24 h at any of dose and day | Tsangalis et al. [32] |
| NFS: 68, 100, 169 mg | Urine: 0–24 h at day 4, 13, 14 |
FS, fermented soybean; NFS, non-fermented soybean; PostMeno, post menopause; Cmax, maximum plasma concentration; AUC, area under the curve; Tmax, time of the maximal plasma concertation.
![]()
In summary, fermentation was observed to increase the initial concentration of isoflavones in the blood after soy food intake. However, urinary total isoflavone was not affected by fermentation at 24 or 48 h after ingestion. Isoflavones are absorbed in the form of aglycone in the small intestine, and the composition of various glucosides can affect the absorption and metabolism of isoflavones. Unfortunately, the effects of chemical structure on bioavailability and biological benefits have not yet been assessed in detail. Thus, further studies are needed to develop isoflavone exposure biomarkers.
Go to :
STUDIES ON ISOFLAVONE EXPOSURE BIOMARKERS
Exposure biomarkers provide objective exposure measures that explain bioavailability and metabolism after ingestion and are not vulnerable to bias reporting. Therefore, biomarkers can be used to improve the accuracy of assessments of isoflavone exposure [33]. Typically, high isoflavone levels in urine and blood are associated with soy consumption, indicating dietary isoflavone intake.
Table 2 shows recent studies on isoflavone exposure biomarkers in blood and urine. Recent isoflavone intakes estimated by 24-h recall showed a high correlation (r = 0.46–0.97) [343536373838] with urinary isoflavones. However, the food frequency questionnaire (FFQ), which reflects habitual dietary intake, showed a weak correlation (r = 0.29; P < 0.01) with isoflavone concentrations in urine [36]. Chávez-Suárez et al. [37] reported that total isoflavone and individual isoflavone intakes estimated by FFQ showed no correlation to urinary levels (r = 0.002; P = 0.983), whereas 24-h recalls of total isoflavones (r = 0.460; P < 0.001) and genistein (r = 0.374; P = 0.002) were correlated to their urinary levels. Exposure biomarkers of isoflavones, as well as those of carotenoids, folate, cruciferous vegetables, and fruits showed moderately valued correlations (r = 0.30–0.49) with dietary intake [39]. The correlations between biomarkers and FFQ were slightly lower than those of the corresponding 24-h dietary recalls [39]. Further, the concentrations of isoflavone in urine or blood can be used to provide relative validation of isoflavone intake estimates from a self-reported FFQ [3940414243].
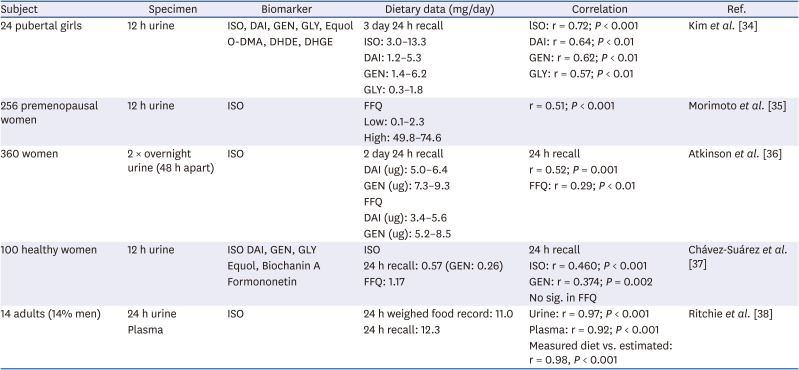
Table 2
Studies on dietary isoflavones as biomarkers in blood or urine
| Subject | Specimen | Biomarker | Dietary data (mg/day) | Correlation | Ref. |
|---|---|---|---|---|---|
| 24 pubertal girls | 12 h urine | ISO, DAI, GEN, GLY, Equol | 3 day 24 h recall | lSO: r = 0.72; P < 0.001 | Kim et al. [34] |
| O-DMA, DHDE, DHGE | ISO: 3.0–13.3 | DAI: r = 0.64; P < 0.01 | |||
| DAI: 1.2–5.3 | GEN: r = 0.62; P < 0.01 | ||||
| GEN: 1.4–6.2 | GLY: r = 0.57; P < 0.01 | ||||
| GLY: 0.3–1.8 | |||||
| 256 premenopausal women | 12 h urine | ISO | FFQ | r = 0.51; P < 0.001 | Morimoto et al. [35] |
| Low: 0.1–2.3 | |||||
| High: 49.8–74.6 | |||||
| 360 women | 2 × overnight urine (48 h apart) | ISO | 2 day 24 h recall | 24 h recall | Atkinson et al. [36] |
| DAI (ug): 5.0–6.4 | r = 0.52; P = 0.001 | ||||
| GEN (ug): 7.3–9.3 | FFQ: r = 0.29; P < 0.01 | ||||
| FFQ | |||||
| DAI (ug): 3.4–5.6 | |||||
| GEN (ug): 5.2–8.5 | |||||
| 100 healthy women | 12 h urine | ISO DAI, GEN, GLY | ISO | 24 h recall | Chávez-Suárez et al. [37] |
| Equol, Biochanin A | 24 h recall: 0.57 (GEN: 0.26) | ISO: r = 0.460; P < 0.001 | |||
| Formononetin | FFQ: 1.17 | GEN: r = 0.374; P = 0.002 | |||
| No sig. in FFQ | |||||
| 14 adults (14% men) | 24 h urine | ISO | 24 h weighed food record: 11.0 | Urine: r = 0.97; P < 0.001 | Ritchie et al. [38] |
| Plasma | 24 h recall: 12.3 | Plasma: r = 0.92; P < 0.001 | |||
| Measured diet vs. estimated: r = 0.98, P < 0.001 |
ISO, total isoflavone; GEN, genistein; DAI, daidzein; GLY, glycitein; O-DMA, O-desmethylangolensin; DHDE, dihydrodaidzein; DHGE, dihydrogenistein; FFQ, food frequency questionnaire.
![]()
Although the association between dietary isoflavone intake and urinary isoflavone excretion is weak at a low intake, urinary isoflavones can be used to discriminate between low- and high-soy diets across the population [35]. Following the study of relative validity and reproducibility of FFQ using 24-h dietary recalls and urinary biomarkers, the correlation between soy food intake and isoflavone levels in urine was different among ethnic groups in Asia [43]. Depending on the source and processing methods of soy food that contribute to isoflavone exposure, bioavailability may vary, leading to differences in urine or blood isoflavone levels.
Go to :
RECENT STUDIES ON THE HEALTH EFFECTS OF SOY ISOFLAVONES
Five studies investigated the association of the biomarkers for dietary isoflavone with chronic disease and cancer risk (Table 3). Urinary isoflavone levels were not associated with CVD-related markers such as low-density lipoprotein-, high-density lipoprotein-cholesterol, and blood pressure [44]. A null association was also observed between urinary isoflavone and the risk of ischemic stroke [45]. The authors determined this was because urinary isoflavone excretion reflected short-term exposure, and participants showed differences in biological metabolism such as gut microbiota [45]. On the other hand, the levels of total isoflavone in urine showed no association with T2D, whereas higher daidzein levels in urine were associated with decreased risk of T2D (OR, 0.71; 95% CI, 0.55–0.93) [46]. Plasma isoflavone levels have been used as exposure biomarkers to predict the risk of prostate or liver cancer. Levels higher than the median of plasma genistein (> 640.2 nmol/L) were associated with a low risk of prostate cancer (OR, 0.31; 95% CI, 0.13–0.71) [47]. However, plasma genistein, daidzein, glycitein, and equol showed no association with primary liver cancer [48].
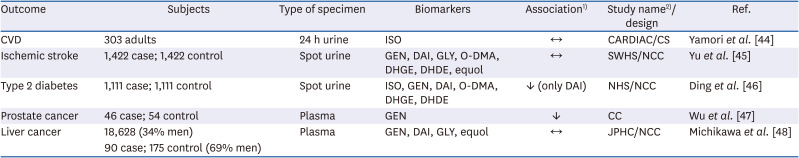
Table 3
The observational studies on the disease risk of dietary isoflavones by using level in biological samples as biomarker
| Outcome | Subjects | Type of specimen | Biomarkers | Association1) | Study name2)/design | Ref. |
|---|---|---|---|---|---|---|
| CVD | 303 adults | 24 h urine | ISO | ↔ | CARDIAC/CS | Yamori et al. [44] |
| Ischemic stroke | 1,422 case; 1,422 control | Spot urine | GEN, DAI, GLY, O-DMA, DHGE, DHDE, equol | ↔ | SWHS/NCC | Yu et al. [45] |
| Type 2 diabetes | 1,111 case; 1,111 control | Spot urine | ISO, GEN, DAI, O-DMA, DHGE, DHDE | ↓ (only DAI) | NHS/NCC | Ding et al. [46] |
| Prostate cancer | 46 case; 54 control | Plasma | GEN | ↓ | CC | Wu et al. [47] |
| Liver cancer | 18,628 (34% men) | Plasma | GEN, DAI, GLY, equol | ↔ | JPHC/NCC | Michikawa et al. [48] |
| 90 case; 175 control (69% men) |
CVD, cardiovascular disease; ISO, total isoflavone; GEN, genistein; DAI, daidzein; GLY, glycitein; CS, cross-sectional; FFQ, food frequency questionnaire; Q, quartile or quintile; NCC, nested case-control; HR, hazard ratio; CI, confidence interval; O-DMA, O-desmethylangolensin; DHGE, dihydrogenistein; DHDE, dihydrodaidzein; WHO, World Health Organization; CARDIAC, Cardiovascular Disease and Alimentary Comparison; SWHS, The Shanghai Women's Health Study; NHS, Nurses' Health Study; JPHC, The Japan Public Health Center-based prospective.
1)↑ positive association, ↓ inversely association, ↔ no association; 2)WHO-coordinated CARDIAC, SWHS, NHS, JPHC.
![]()
The exposure biomarkers for isoflavones proposed in recent studies were presented as total isoflavone or aglycone (parent isoflavone compound) equivalent doses. Most of the conjugated forms of dietary soy isoflavones in blood or urine after absorption are sulfates or glucuronides [24]. Circulating isoflavone conjugates may act as a source of aglycone at specific target tissues [26]. However, the health effects and mechanisms of action of conjugated metabolites in humans remain unclear. In addition, recent studies may be limited due to possible measurement errors, variable food sources, and differing levels of isoflavone intake among populations. The effects of isoflavone on chronic disease and cancer risk merit further study using both biomarkers and data on dietary intake.
Go to :
CONCLUSION
The composition of isoflavone in soy foods differs based on the method of processing, fermentation conditions, and the microorganisms present. Additionally, bioavailability may vary depending on the soy food source, processing methods, and individual differences in biological metabolism. These factors lead to differences in urine or blood isoflavone concentrations. Human metabolites of isoflavone can be used as biomarkers to improve the accuracy of exposure assessments for dietary isoflavone. However, the effects of chemical structures on bioavailability and biological benefits have not yet been assessed in detail. Therefore, these factors are rarely considered in epidemiological and clinical studies, and their health impacts remain unclear.
Most studies on isoflavone showed that the biologically active molecules are either aglycones or their unconjugated metabolites. However, the conjugates may be considered either biologically active themselves or serve as excellent sources of biologically active compounds within target cells by deconjugation. Data on human metabolites without enzymatic hydrolysis of conjugates in blood and urine are not yet sufficient to understand the exposure biomarkers of soy isoflavone intake. However, direct quantification of conjugates in biological samples without enzymatic hydrolysis would pose a major challenge. It will be valuable to develop exposure biomarkers in future studies on soy isoflavones for different types of soy foods in order to fully understand the association between various soy foods and risks of disease.
Go to :
 XML Download
XML Download