

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Despite a decrease in incidence in the recent decade, gastric cancer is still one of the most common causes of cancer death worldwide [1]. In South Korea, gastric cancer is the most frequent cancer and the fourth most common cause of cancer death. In countries with a high incidence of gastric cancer, such as Korea and Japan, a nationwide early screening system was introduced to improve the early detection rate of gastric cancer, thereby increasing its 5-year survival rate compared to other cancers [2]. However, because stomach pain is seen only in advanced stages of gastric cancer, the cure rate decreases sharply if not detected early. The probability of metastasis increases as gastric cancer progresses; more than 80% of advanced gastric cancer patients have metastasis to lymph nodes [3]. Patients with early gastric cancer can increase their survival rate compared to other cancers with conventional treatment methods such as surgery, chemotherapy and radiation therapy. However, these conventional therapies are not entirely appropriate for advanced gastric cancer patients as well. The cause of death of an advanced gastric carrier patient is metastasis, so it is the measures to address the metastasis that is needed [4].
Metastasis is associated with more than 90% of cancer-associated mortality [5]. Cancer cells of the primary tumor invade the surrounding tissues locally to intravasation of the lymphatic and vascular system, where they survive and migrate to the microvessels of the distant tissues through the bloodstream (extravasation). There, they adapt to the unfamiliar environment of these tissues, survive and produce secondary tumors through cell proliferation (colonization) [6]. Thus, it is a valid approach to prevent metastasis in order to effectively treat cancers, including gastric cancer. Bioactive compounds obtained naturally have been noticed as new agents for metastasis prevention.
Rhus verniciflua Stokes (RVS) belongs to the Anacardiaceae family, usually known as the lacquer tree. RVS has been used as a traditional herbal medicine for the relief of pain caused by various diseases, such as cancer, as well as the treatment of digestive system diseases in East Asian countries, such as Korea, Japan, and China [78]. RVS contains numerous useful compounds, such as quercetin, fustin, fisetin, sulfuretin, and butein. Previous research has shown that Rhus verniciflua Stokes extract (RVSE) has antioxidant, anti-proliferative, anti-inflammatory, and anti-tumor effects [89101112131415161718]. Several in vivo and in vitro studies demonstrated antitumor effects of RVSE and/or its compounds in gastric, breast, liver, lymphoma, and osteosarcoma [131920]. The mechanism of the antitumor effect of RVS is still uncertain, but possible mechanisms include inhibition of the phosphoinositol-3-kinase (PI3K)-Akt/protein kinase B pathway [20], activation of AMP-activated protein kinase (AMPK) [21], cell cycle arrest [22], decreased manganese superoxidase activity or glutathione content [23], activation of caspase, downregulation of Bcl-2 and Mcl-1, upregulation of Bax, P53 hyperphosphorylation, and S6 hypophosphorylation [7].
Despite the pharmacological effects of RVS, its use has been limited because it contains an allergenic substance, urushiol (a mixture of several derivatives of catechol). Therefore, removing urushiol from RVSE is critical for its safe use. Allergen-free RVSE, as well as common RVSE, have been reported to have growth inhibitory and apoptosis-inducing effects in A549 human lung cancer cells [7] and have been shown to be useful in the treatment of advanced or metastatic cancers [19]. However, the components contained in allergen-free RVSE and their subsequent effects have been shown to differ depending on the specific method of detoxification (removing urushiol). Thus, it is possible to maximize the specific effect by modifying the detoxification method [17]. We modified the existing extract method to create a new, concentrated and purified allergen-free RVSE. Analysis of the compound showed that this RVSE contained 69.8% and 48.6% more fustin and fisetin, respectively, compared to those extracted by the conventional method. This study was conducted to investigate the effect of this new RVSE on metastasis in AGS cells, a human gastric cancer cell line.
Go to : 
MATERIALS AND METHODS
Materials
The materials used in this research were purchased from the following suppliers: RPMI1640 and fetal bovine serum (FBS) from Welgene (Daegu, Korea); 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), anti-β-actin antibody from Sigma-Aldrich (St. Louis, MO, USA); antibodies against signal transducer and activator of transcription (STAT) 3 (PY705) and STAT3 from Cell Signaling Technology (Beverly, MA, USA); antibodies against matrix metalloproteinase (MMP)-9 and vascular endothelial growth factor (VEGF) from Santa Cruz Biotechnology (Santa Cruz, CA, USA); antibodies against urokinase-type plasminogen activator (uPA) and plasminogen activator inhibitor 1 (PAI-1) from Calbiochem (Darmstadt, Germany); Unless noted otherwise, all other materials were purchased from Sigma-Aldrich.
Preparation of RVSE
RVSEs were prepared by Medience Co., Ltd. (Chuncheon, Korea). Dried RVS cultivated in Chuncheon, Korea were purchased from an herbal medicine store (Chuncheon, Korea). Dried RVS were sliced to 2 cm, after which, 100 g of sliced RVS was refluxed in 1 L of water at 100°C for 10 h. This extraction procedure was repeated twice. The extracts were filtered through Whatman filter paper #2, after which the filtrate was freeze-dried. The resulting powder was used as pre-Rhus verniciflua Stokes extract (pRVSE). To prepare concentrated and refined RVSEs, 100 g of pRVSE was refluxed in 1 L of 95% ethanol at room temperature for 1 h. The extract was centrifuged at 3,000 rpm for 10 min, after which the supernatant was collected and dried below 60°C in a vacuum. The resulting powder was used as RVSE. The pRVSE and RVSE extracts were stored −20°C until further use.
High-performance liquid chromatography (HPLC) analysis
Both pRVSE and RVSE were analyzed using HPLC (SPD-20A; Shimadzu, Tokyo, Japan) with a C18 column packed with 5 µm diameter particles (250 × 4.6 mm, YMC-Pack ODS-A, YMC Co., Ltd., Kyoto, Japan) and detection at 520 nm. Fustin and fisetin used as reference standards were purchased from Biopurity Phytochemicals Ltd. (Chengdu, China). The operating conditions were as follows. The mobile phase solvents used were 5% acetic acid, methanol, and acetonitrile (70:20:10). The flow rate and injection volume were 1.0 mL/min and 10 µL, respectively. Before performing HPLC, the samples and reference standards were dissolved in methanol and filtered through 0.45 µm filters. The chromatographic peaks of the analytes were confirmed by comparing their ultraviolet spectra and retention time with those of the reference standards.
Cell culture and cell viability assay
AGS human gastric adenocarcinoma cells were acquired from the American Type Culture Collection (Manassas, VA, USA). AGS cells were cultured in RPMI1640 containing 100 mL/L FBS, 100,000 U/L penicillin and 100 mg/L streptomycin at 37°C in a humidified atmosphere of 5% CO2 in air. To determine cell viability, the cells were plated in 24-well plates at 5 × 104 cells/well and incubated for 24 h. After 24 h, the cells were treated with pRVSE or RVSE at concentrations ranging from 0 to 150 µg/mL and incubated for 24 h. pRVSE and RVSE were dissolved in dimethyl sulfoxide (DMSO). DMSO was applied to all cells until they reached a final concentration of 0.01%. Cell viability was determined by MTT assay as described previously [24].
In vitro migration assay
The ability of AGS cells to migrate was determined using the CytoSelect™ 24-well cell Migration Assay kit (Cell Biolabs Inc., San Diego, CA, USA). The AGS cells were serum-deprived in RPMI1640 supplemented with 10 mL/L FBS (serum-deprivation medium) for 24 h. The serum-deprived AGS cells were plated in 8 µm pore size polycarbonate membrane inserts in 24-well plates at 1.5 × 105 cells/insert and treated with the different concentrations of pRVSE and RVSE in serum-deprivation medium. The lower chamber of the well was filled with RPMI1640 containing 100 mL/L hormone-free, gelatinase-free FBS as a chemoattractant. After 4 h of incubation, the migrated cells were stained with hematoxylin and eosin and observed with a light microscope. To quantify the migration ability, the migrated cells were stained with the CytoSelect™ Cell Stain Solution and Extraction Solution to extract the dye stain. Then, the absorbance was measured at 560 nm with a microplate reader, according to the manufacturer's instructions.
In vitro invasion assay
The invasion ability of AGS cells was measured using 8 µm pore size polycarbonate membrane inserts coated with dried basement membrane matrix solution contained in the CytoSelect™ 24-well Cell Invasion Assay kit (Cell Biolabs Inc.). The invasion assay was performed in the same way as the migration assay detailed above.
Preparation of conditioned media and cell lysates
AGS cells were plated in 100 mm dishes at 1 × 106 cells/dish and incubated for 24 h. Next, the cells were serum-deprived in serum-deprivation medium for 24 h and treated with the indicated concentration of RVSE for 24 h. Media conditioned for 24 h were collected. The cells were lysed as described previously [25]. The protein contents of the conditioned media and cell lysates were determined using a BCA protein assay kit (Thermo Fisher Scientific, Rockford, IL, USA) according to the manufacturer's instruction.
Protein antibody array
Conditioned media and cell lysates were analyzed by a Proteome Profiler™ Antibody Array kit (R&D Systems, Minneapolis, MN, USA) to estimate the expression levels of various angiogenesis (metastasis)-related proteins, according to the manufacturer's instructions. The relative abundance of each protein spot was quantified using an Image Quant™ LAS 500 imaging system (GE Healthcare Bio-Sciences AB, Uppsala, Sweden). The expression levels were normalized to the control protein.
Western blot analysis
Western blot analyses were performed as described previously [25]. Blots were detected with Luminata™ Forte Western HRP Substrate (Millipore Corporation, Billerica, MA, USA). The relative intensity of each protein band was quantified using an Image Quant™ LAS 500 imaging system (GE Healthcare Bio-Sciences AB). The expression levels were normalized to β-actin expression.
Statistical analyses
All results were presented as the mean ± SEM. The statistical significance of results among the treatment group differences was analyzed via one-way analysis of variance, followed by Duncan's multiple range test. Differences were considered significant at P < 0.05.
Go to :
RESULTS
Quantification of flavonoid contents of RVSE
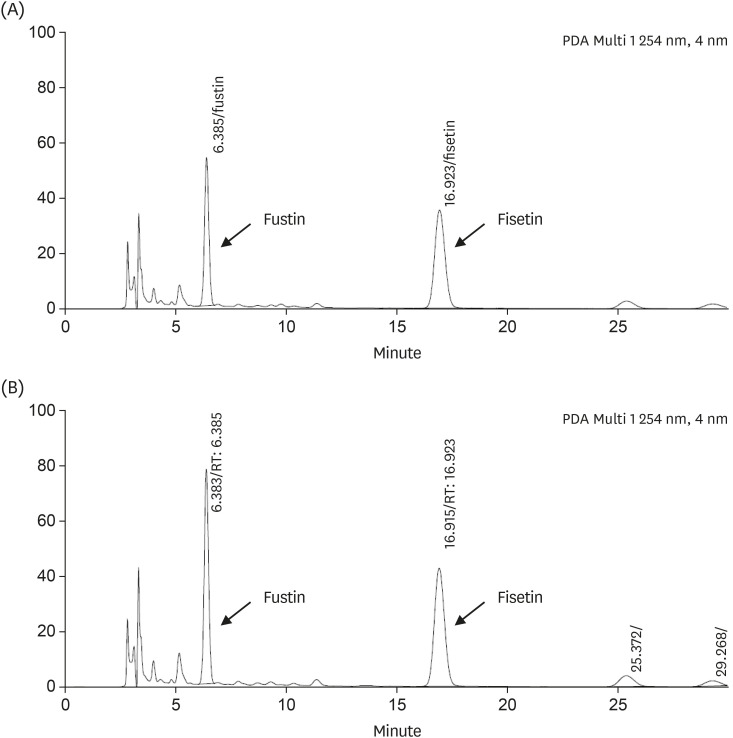
As a result of analyzing the flavonoid components of pRVSE and RVSE with HPLC, urushiol was not detected in both pRVSE and RVSE, and fustin and fisetin were detected in pRVSE and RVSE. RVSE has an increased fusitin and fisetin content than pRVSE. pRVSE contained 129 mg/g and 57 mg/g of fustin and fisetin, respectively, while RVSE contained 219 mg/g and 82 mg/g of fustin and fisetin, respectively. These HPLC results indicate that RVSE, a product of the concentration and refining process, contained more flavonoids including fustin and fisetin (Fig. 1).
Effect of RVSE on cell viability of AGS cells
Prior to investigating the effect of pRVSE and RVSE on the migration of AGS cells, an MTT assay was done in order to determine the pRVSE and RVSE concentrations shown to be non-cytotoxic to AGS cells. pRVSE significantly increased AGS cell viability at 50 and 100 µg/mL concentrations but did not affect viability at 150 µg/mL. RVSE did not affect cell viability at concentrations ranging from 25 to 100 µg/mL. However, the viability of cells treated with 150 µg/mL of RVSE was significantly decreased compared to the non-treated cells (Fig. 2). Based on these results, the subsequent experiments were conducted at concentrations of 0–100 µg/mL to exclude the possibility that the inhibition of migration and invasion of AGS cells was due to cytotoxic effects of pRVSE and RVSE.
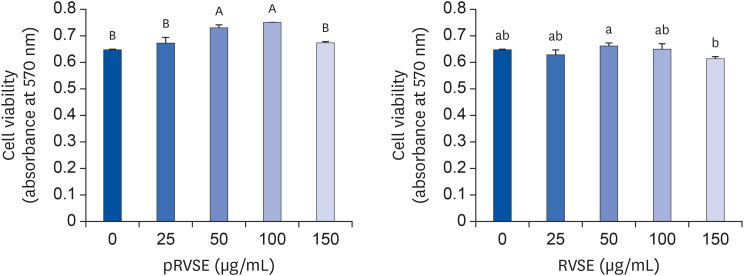 | Fig. 2Effect of pRVSE and RVSE on the viability of AGS cells.Cells were plated in 24-well plates at 5 × 104 cells/well in DMEM supplemented with 100 ml/L FBS. After 24 h, the cells were serum-deprived with DMEM supplemented with 10 mL/L charcoal stripped FBS (SDM) for 24 h. After serum deprivation, the cells were incubated in serum-deprivation medium at varying concentration (0, 25, 50, 100, 150 µg/mL) of pRVSE or RVSE. Viable cell numbers were estimated by MTT assays. Each bar represents the mean ± SEM (n = 4). Means without a common letter differ, P < 0.05.
pRVSE, pre-Rhus verniciflua Stokes extract; RVSE, Rhus verniciflua Stokes extract; DMEM, Dulbecco's Modified Eagle's Medium; FBS, fetal bovine serum; SDM, serum-deprivation medium; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide.
|
Effect of RVSE on the migration and invasion of AGS cells
To compare the effect of pRVSE and RVSE on the migratory ability of AGS cells, we conducted an in vitro migration assay. As shown in Fig. 3, pRVSE and RVSE dose-dependently inhibited the migration of AGS cells. However, when cells were treated with the same concentration of pRVSE and RVSE, the number of migrated cells was markedly reduced by RVSE treatment compared to pRVSE treatment. These results indicate that RVSE may inhibit AGS cells migration more than pRVSE.
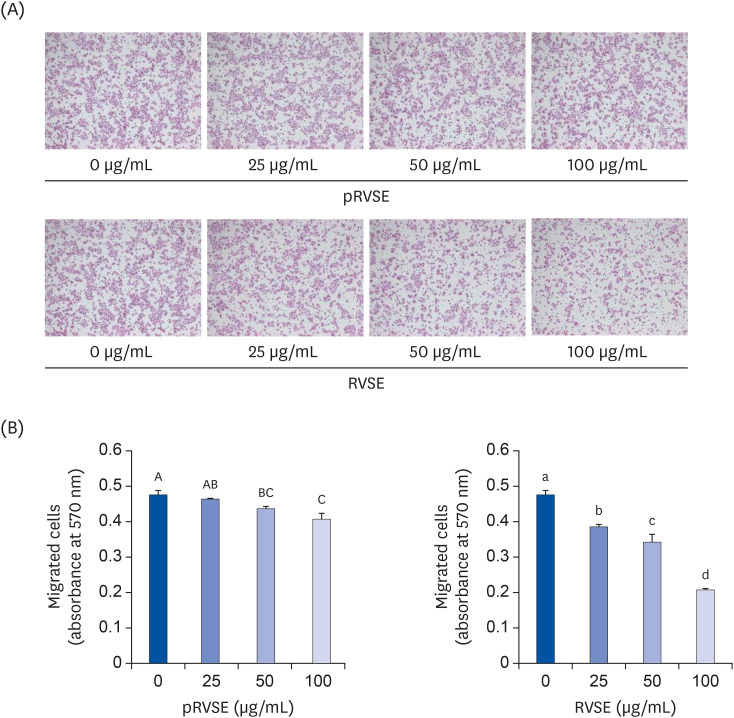 | Fig. 3Effect of pRVSE and RVSE on the migration of AGS cells.The migration ability of AGS cells was measured using the CytoSelect™ 24-well Cell Migration Assay kit. The cells were serum-deprived in DMEM supplemented with 10 mL/L charcoal-stripped FBS for 24 h. Serum-deprived cells were plated in trans-well inserts in 24-well plates at 1.5 × 105 cells/filter and treated with 0–100 µg/mL pRVSE or RVSE. The lower chamber of the well was filled with SDM supplemented with 1g/L BSA as a chemoattractant. The cells were then incubated for 4 h. (A) The migrated cells were stained with hematoxylin and eosin. (B) The migrated cells were stained with cell stain solution and quantified by measuring the absorbance at 560 nm after dye extraction. Each bar represents the mean ± SEM (n = 4). Means without a common letter differ, P < 0.05.
pRVSE, pre-Rhus verniciflua Stokes extract; RVSE, Rhus verniciflua Stokes extract; DMEM, Dulbecco's Modified Eagle's Medium; FBS, fetal bovine serum; SDM, serum-deprivation medium; BSA, bovine serum albumin.
|
We subsequently attempted to determine whether RVSE inhibits the invasive ability of AGS cells. RVSE inhibited the invasion of AGS cells in a dose-dependent manner. Treating AGS cells with 100 µg/mL of RVSE resulted in a 52.7 ± 3.2% reduction in the number of cells invaded (Fig. 4).
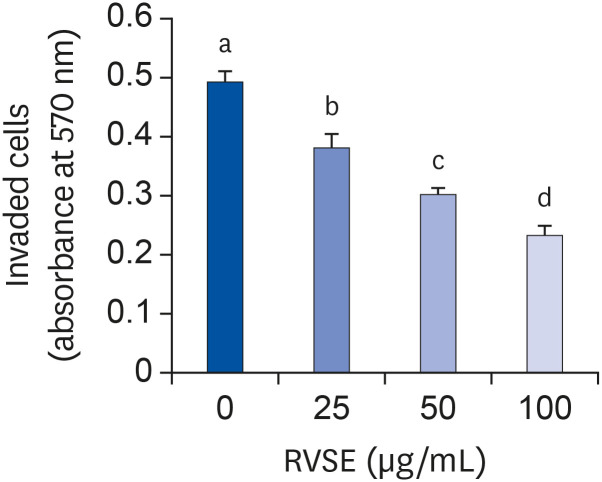 | Fig. 4Effect of RVSE on the invasion of AGS cells.The invasion ability of AGS cells was measured using the CytoSelect™ 24-well Cell Invasion Assay kit. Serum-deprived AGS cells were plated in trans-well inserts in 24-well plates at 1.5 × 105 cells/filter and treated with 0–100 µg/mL RVSE. The lower chamber of the well was filled with SDM supplemented with 1g/L BSA as a chemoattractant. The cells were then incubated for 6 h. The invaded cells were stained with cell stain solution and quantified by measuring the absorbance at 560 nm after dye extraction. Each bar represents the mean ± SEM (n = 4). Means without a common letter differ, P < 0.05.
RVSE, Rhus verniciflua Stokes extract; SDM, serum-deprivation medium; BSA, bovine serum albumin.
|
Effect of RVSE on metastasis-related proteins expression in AGS cells
To determine the mechanisms by which RVSE inhibits the migration and invasion of AGS cells, we measured the relative expression levels of metastasis (angiogenesis)-related proteins in total cell lysates and conditioned media using a Proteome Profiler™ Antibody Array kit. Among the 55 kinds of proteins measureable with the kit, MMP-9 protein expression was markedly decreased and PAI-1 protein expression was dramatically increased in the total cell lysates and conditioned media of cells treated with 100 µg/mL RVSE (data not shown). To validate these results, we subsequently performed Western blot analyses of total cell lysates for MMP-9 and PAI-1. As shown in Fig. 5, RVSE significantly inhibited MMP-9 protein expression in AGS cells. PAI-1 protein expression was markedly increased by RVSE treatment. Treatment of AGS cells with 100 µg/mL RVSE resulted in a 443 ± 60% increase in PAI-1 protein expression. In addition to PAI-1, the effect of RVSE on uPA protein expression was investigated. RVSE dose-dependently inhibited the uPA protein expression in AGS cells (Fig. 6).
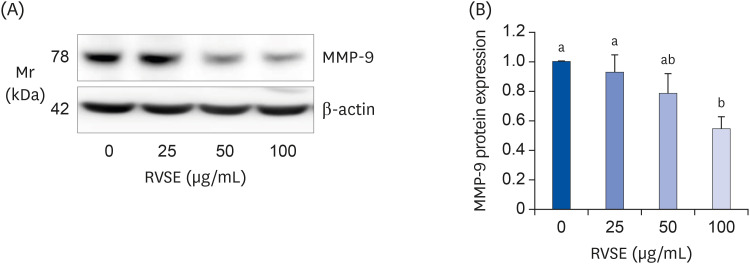 | Fig. 5Effect of RVSE on MMP-9 protein expression in AGS cells.Cells were plated in 100 mm dishes at 1 × 106 cells/dish in DMEM containing 100 mL/L FBS. After 24 h, the cells were serum-deprived with SDM for 24 h. After serum-deprivation, the cells were incubated with 0–100 µg/mL RVSE for 24 h. The cell lysates were subjected to Western blotting with MMP-9 antibody. (A) Photographs of chemiluminescent detection of the blots, which are representative of 3 independent experiments, are shown. (B) Quantitative analysis of the blots. The relative abundance of each band to its own β-actin was quantified, and the control levels were set at 1. Each bar represents the adjusted mean ± SEM (n = 4). Means without a common letter differ, P < 0.05.
RVSE, Rhus verniciflua Stokes extract; MMP, matrix metalloproteinase; DMEM, Dulbecco's Modified Eagle's Medium; FBS, fetal bovine serum; SDM, serum-deprivation medium.
|
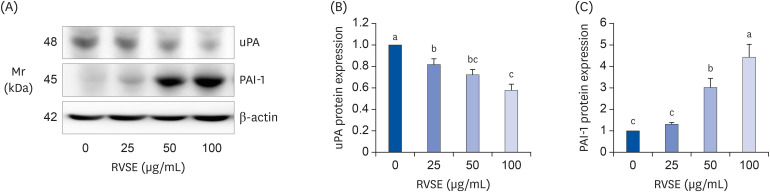 | Fig. 6Effect of RVSE on uPA and PAI-1 protein levels in AGS cells.The cells were plated and treated as described in Figure 5. The cell lysates were subjected to Western blotting with uPA and PAI-1 antibodies. (A) Photographs of chemiluminescent detection of the blots, which are representative of 3 independent experiments, are shown. (B) Quantitative analysis of the blots. The relative abundance of each band to its own β-actin was quantified, and the control levels were set at 1. Each bar represents the adjusted mean ± SEM (n = 4). Means without a common letter differ, P < 0.05.
RVSE, Rhus verniciflua Stokes extract; uPA, urokinase-type plasminogen activator; PAI-1 plasminogen activator inhibitor-1.
|
Effect of RVSE on the phosphorylation of STAT3 in AGS cells
We next determined the effect of RVSE on the phosphorylation of STAT3 by Western blot analysis. RVSE significantly suppressed the phosphorylation of STAT3 in AGS cells dose-dependently. The protein expression of STAT3 in AGS cells was not altered by RVSE treatment (Fig. 7).
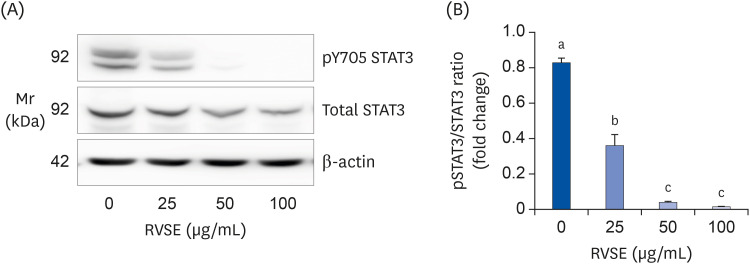 | Fig. 7Effect of RVSE on the phosphorylation of STAT3 in AGS cells.The cells were plated and treated as described in Fig. 5. The cell lysates were subjected to Western blotting with pY705 STAT3 and STAT3 antibodies. (A) Photographs of chemiluminescent detection of the blots, which are representative of 3 independent experiments, are shown. (B) Quantitative analysis of the blots. The relative fold-change in pSTAT3 to its own STAT3 was quantified, and the control levels were set at 1. Each bar represents the adjusted mean ± SEM (n = 4). Means without a common letter differ, P < 0.05.
RVSE, Rhus verniciflua Stokes extract; STAT, signal transducer and activator of transcription.
|
Go to :
DISCUSSION
In the traditional East Asian medical system, traditional medicines have been used to treat various diseases, including cancer. As the scientific basis for their efficacy continues to be revealed, the potential use of traditional medicines as therapeutic agents has gained popularity, thanks to their low price, superb efficacy, and few side effects. RVS is also a traditional medicinal used in the Orient for centuries to treat digestive system diseases, liver detoxification, hemostasis, and cough. Previous studies have shown that RVS inhibited cell proliferation and induced apoptosis in tumor cells. In human gastric cancer cell line studies, RVS induced G1 phase cell cycle arrest and promoted the mitochondrial death pathway by inhibiting the PI3K-Akt/PKB pathway [2022]. In an ovarian cancer cell line study, allergen-free RVSE promoted apoptosis through JNK activation [26]. In addition, the anticancer effects of RVS were reported in an in vitro study of various cancer cell lines, including Lewis lung carcinoma, non-small cell lung cancer, human lymphoma, osteosarcoma, and breast cancer cells [2027282930]. Several case studies and clinical studies have reported positive effects of RVS, including tumor size reduction and prolonged survival time, in advanced and metastatic cancer patients [163132333435].
In the past, despite these functional activities, the use of RVS has been very limited because it contains an allergen, named urushiol. So far, various methods of detoxification of RVS have been developed to remove urushiol, such as methods of utilizing organic solvents and oxidizers, enzymatic methods, and mushroom bacteria fermentation methods [363738]. By the way, the main component and bioactivity of the RVS are very different according to the detoxification processing method [17]. Therefore, it is important to identify healthier and safer extraction methods that can selectively remove urushiol alone while preserving beneficial ingredients such as flavonoid compounds and phenolic compounds. Among them, considering the cost and safety aspects of the human body, the physical method using the loss of urushiol activity at high temperatures is known to be the most appropriate. pRVSE is also the removal of urushiol by high temperature extraction. We made RVSE by adding concentration and refining methods here. RVSE, like pRVSE, did not detect urushiol, and the content of fisetin and fustin increased (Fig. 1).
RVS contains various phytochemical components. About 40 types of compounds have been isolated. Of these compounds, three are phenolic acids, four are flavonols, four are flavanols, three are flavones, one is chalconoid, and 2 are tannins [3940]. According to a study that analyzed the content of total phenolic and flavonoids compound by liquid chromatography with tandem mass spectrometry analysis on the skin, stem, and leaves of RVS, gallic acid, dihydroxybenzoic acid, methyl gallate, pentagalloyl glucose, fustin, quercitrin, taxifolin, garbanzol, kaempferol-3-O-rhamnoside, fisetin, sulfuretin, butein, and urushiol were detected. Among them, the content of three substances, methyl gallate, fustin, and quercitrin, was the highest [41].
Among the substances contained in RVS, buetin [424344], kaempferol and quercetin [40] have been studied for a lot of anticancer effects and mechanisms. Fisetin and fustin are expected to have similar efficacy as polyphenol compounds like these substances, but research on the anti-cancer effect of fisetin and fustin is still lacking. Fustin is a kind of flavonol that acts as an antioxidant, such as having a protective effect on 6-hydroxydopamine-induced neuronal cell death [45]. According to a recent comparative analysis of antioxidant activity of substances in Rhus typhina L. stem, fustin's antioxidant activity was reported to be less than methyl gallate, gallic acid and quercetin and greater than vitamin C and rutin [46]. Fisetin is a type of polyphenol belonging to the flavonoid group, and it contains a lot of fruits and vegetables such as strawberries and apples as well as RVS, and has excellent antioxidant properties [47].
Fisetin from RVS significantly reduced inflammatory cytokines such as tumor necrosis factor-α, interleukin (IL)-6, IL-8, and monocyte chemoattractant protein-1 [48]. According to the in vitro studies, the anticancer activity of fisetin appears to be related to the down-regulation of AMPK, cyclooygensae, epidermal growth factor receptor, extracellular signal regulated kinase, MMP, nuclear factor-kappa B, prostate-specific antigen, transcription factor T-cell factor, TNF-related apoptosis inducing ligand, Wnt inhibitory factor, X linked inhibitor of apoptosis, and PI3K/AKT/mTOR pathway [495051525353]. Fisetin also showed anti-cancer effects in cells and animal models by influencing cell cycles [54] and acting as a topoisomerase inhibitor [55].
In this study, we investigated the effect of RVSE on cancer cell metastasis in gastric cancer cells, the most frequent cancer in Koreans. pRVSE and RVSE were not cytotoxic in AGS gastric cancer cells up to a concentration of 100 μg/mL (Fig. 2). Therefore, we observed migration by treating AGS cells with 0, 25, 50, and 100 μg/mL pRVSE and RVSE. The results showed that the number of migrated cells decreased with increasing concentrations of both pRVSE and RVSE. In particular, RVSE was found to reduce migration much more dose-dependently than pRVSE (Fig. 3A and B). In subsequent invasive studies, RVSE reduced the number of invaded cells in a dose-dependent manner (Fig. 4). Migration and invasion are crucial steps in metastasis. Our results indicate that RVSE may inhibit metastasis.
MMPs are a class of endopeptidases that degrade and remodel the extracellular matrix. The migration and invasion of cancer cells to surrounding tissues are mediated by MMPs, particularly MMP-2 and MMP-9 [56]. Studies to determine the molecular mechanism of anti-metastasis in AGS cells, showed that treatment with RVSE dose-dependently decreased the level of MMP-9 protein. This result confirmed that the RVSE-induced anti-metastasis pathway was associated with MMP-9 inhibition in AGS cells. The uPA expression was decreased and the PAI-1 expression was increased by RVSE treatment. uPA, also known as urokinase, is a serine protease. The primary physiological substrate of this enzyme is plasminogen, is an inactive form of the serine protease plasmin. When plasmin is activated by uPA, it degrades the extracellular matrix and thus, uPA is considered to a cancer metastasis-promoting factor [57]. In contrast, PAI-1, also known as endothelial plasminogen activator inhibitor or serpin E1, is an inhibitor of tissue plasminogen activator (tPA) and uPA [58]. Our results showed that RVSE inhibited degradation of the extracellular matrix by inhibiting the activation of plasminogen, thereby inhibiting cancer metastasis.
STAT3 belongs to the STAT protein family. STAT3 regulates the expression of various genes in response to cell stimulation and plays an essential role in many cellular processes, such as cell growth and apoptosis [59]. In response to cytokines and growth factors, STAT3 is phosphorylated by receptor-associated Janus kinases and translocates to the cell nucleus where they act as transcription activators. For example, STAT3 induces the phosphorylation of amino acids at specific positions in response to ligands, such as interferons, epidermal growth factor, IL-5, IL-6, and mitogen-activated protein kinases [60616263]. Constitutive STAT3 activation has been correlated with various human cancers and usually suggests substandard prognosis [64656667]. Increased STAT3 activation in cancer cells has been shown to induce metastasis by causing changes in protein complexes that regulate the expression of inflammatory genes [68]. In our study, RVSE treatment showed a dose-dependent reduction of phosphorylated STAT3 expression. Based on these results, RVSE with high fisetin and fustin content suppresses phosphorylation of STAT-3, thereby inhibiting MMP-9 and uPA expression, and this prevents metastasis by inhibiting the decomposing and reconstruction of the extracellular membrane of target tissue where metastasis will occur. In the future, the cell- and its surrounding microenvironment-based mechanisms in relation to the metastasis inhibitory effect of RVSE need to be studied in more diverse ways.
In conclusion, this in vitro study demonstrated that RVSE effectively inhibited migration and invasion of AGS cells. MMP-9, uPA, PAI-1, and STAT3 were identified as key mediators modulating the metastasis of AGS cells. MMP-9 and uPA expression and STAT3 phosphorylation were significantly decreased and PAI-1 expression was significantly increased by RVSE treatment. Our results showed that RVSE inhibited the migration and invasion of AGS cells, which was mediated through the down-regulation of MMP-9 and uPA and the up-regulation of PAI-1. We suggest that RVSE could be a natural compound candidate for a preventive and/or therapeutic agent against gastric cancer metastasis.
Go to :
 XML Download
XML Download