

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
In recent decades, basic medicinal research has been actively conducted using natural products from food sources. Notably, extracts from plants have been demonstrated to help prevent diseases or promote health, as well as to provide nutritional supplementation [123]. These natural products are especially appealing from a medicinal standpoint as they are already known to be safe for human consumption.
Several studies have reported the immune-enhancing effects of natural product extracts. For example, Graviola leaf extract can activate the innate immune system through the production of nitric oxide (NO) and tumor necrosis factor (TNF)-α in RAW 264.7 cells [4]. The water-soluble polysaccharide fraction (LLP-1A) from Lilium lancifolium has also been shown to induce macrophage activation through the nuclear factor kappa B (NF-κB) signaling pathway [5]. NF-κB is a transcription factor that regulates various physiological processes, including immune responses and cell survival [6]. Activated NF-κB by natural product extracts enters the nucleus to induce the expression of NO and pro-inflammatory cytokines, such as TNF-α, interleukin (IL)-1β, and IL-6 [78].
Recent studies showed that AMP-activated kinase (AMPK) may act as an inhibitor of acute pro-inflammatory responses [910]. Although the inhibitory mechanism of these responses by AMPK in macrophages is unclear, it is recently reported that AMPK regulates multiple downstream targets such as acetyl-CoA carboxylase (ACC) activity [10]. Suppression of AMPK, using a dominant negative construct or RNA interference, increases NF-κB signaling [911], whereas activation of AMPK using a constitutively active construct or an AMPK activator, 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), decreases NF-κB signaling in macrophage cells [911].
Platycodon grandiflorum (PG) has been used as an oriental herbal medicine native to East Asia (Korea, China, and Japan). It has various beneficial effects such as anti-inflammatory [12] and antibacterial [13] activities, improvement of liver function [14], and enhancement of cognitive ability [15]. In particular, platycodin D and platycodin D3 as main active ingredients of PG have anti-inflammatory activities, as it was shown to mitigate the inflammatory response induced by lipopolysaccharides (LPS) in RAW 264.7 (a macrophage cell line) cells [16]. Moreover, PGPStc, a polysaccharide derived from PG, stimulates the proliferation of spleen lymphocytes and promotes cell cycle progression of T cells [17], suggesting that PG may have immunomodulatory effects.
Fermented PG extract (FPGE), which was made by incubation with Saccharomyces cerevisiae, increased the platycodin D content when compared to raw PG [18]. However, the underlying immune-enhancing mechanisms of FPGE in macrophage cells are largely unknown.
In the present study, we assessed whether FPGE has immune-enhancing function by measuring the effect of FPGE on the proliferation of RAW 264.7 cells and production of pro-inflammatory cytokines. We also investigated the NF-κB and AMPK signaling pathways to determine the underlying mechanisms of the immunomodulatory effects of FPGE in RAW 264.7 cells.
MATERIALS AND METHODS
Cell culture and reagents
RAW 264.7 mouse macrophage cells were purchased from the Korean Cell Line Bank (Seoul, Korea). Cells were cultured in Dulbecco's Modified Eagle Medium containing 10% fetal bovine serum and 1% antibiotic-antimycotic in a 5% CO2 humidified incubator at 37°C. All cell culture reagents were purchased from GIBCO (Gaithersburg, MD, USA) and other reagents not specified were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Preparation of the extract from FPGE
Powdered FPGE was produced by SK Bioland (Ansan, Korea) using the standard production process. Briefly, roots of Korean PG were washed and ground. Two kilograms of the powder was placed into the fermenter and 8 kg of tap water was added. The mixture was stirred at 100 rpm during sterilization at 121°C for 1 h. The extract was inoculated with yeast (S. cerevisiae) and cultured for 14 to 16 h at 30°C. The temperature of the cultured solution was adjusted to 50°C, then 8 g of cellulase (0.4%) and 4 g of amylase (0.2%) were added for enzymatic reaction for 16 h. The solution was perlite filtered, and subsequently concentrated through a concentrator (DooYoung Hi-Tech, Seoul, Korea). Gamma cyclodextrin was added to the total solid content to a final concentration of 10%, and the product was autoclaved at 95°C for 1 h. After sterilization, the product was dried using a spray dryer (Ein system, Seoul, Korea). FPGE was previously analyzed by high-performance liquid chromatography-evaporative light scattering detector (Waters, Milford, MA, USA) [18]. The main active ingredient of FPGE was platycodin D, which was 4 times higher content than in PG extract.
Cell viability assay
RAW 264.7 cells were grown in 96-well culture plates (2 × 105 cells/mL). After 24 h, cells were treated with FPGE (0, 0.8, 4, 20, 100, or 500 μg/mL) for 48 h, and cell viability was determined using the Cell Counting Kit-8 (CCK-8) assay according to the manufacturer's instructions (Dojindo Molecular Technologies, Inc., Kumamoto, Japan). Viability was quantified by measuring the absorbance at 450 nm on an Epoch Microplate Spectrophotometer (Biotek Inc., Winooski, VT, USA).
NO measurement
NO levels were measured using Griess reagent (Promega, Madison, WI, USA) according to the manufacturer's instructions. RAW 264.7 cells were grown in 6-well culture plates (2 × 105 cells/mL). After 24 h, cells were treated with LPS (1 μg/mL) or FPGE (0, 20, 100, or 500 μg/mL) for 24 h. The supernatant was harvested and mixed with Griess reagent for 10 min with the substrate solution at room temperature, followed by further incubation with the coloring solution for 10 min in the dark. Absorbance of the mixture was measured at 540 nm using an Epoch Microplate Spectrophotometer (Biotek Inc.).
RNA isolation and real-time reverse transcription polymerase chain reaction (RT-PCR)
RAW 264.7 cells were grown in 6-well culture plates (2 × 105 cells/mL). After 24 h, cells were treated with LPS (1 μg/mL) or FPGE (0, 20, 100, or 500 μg/mL) for 24 h. Total RNA was isolated using the RNA Extraction Kit (iNtRON Biotechnology, Seongnam, Korea), and complementary DNA (cDNA) was synthesized using iScript cDNA synthesis Kit (BioRad, Hercules, CA, USA). Real-time RT-PCR was performed on the synthesized cDNA and analyzed with SYBR Green Master Mix (TaKaRa Bio, Otsu, Japan) using ABI QuantStudio 3 (Applied Biosystems, Foster City, CA, USA). The primer sequences (5′-3′) used in the experiments were as follows: Tnf-α forward, TGTCCCTTTCACTCACTGGC and reverse, CATCTTTTGGGGGAGTGCCT; IL-1b forward, AACTGTTCCTGAACTCAACTGT and reverse, GAGATTTGAAGCTGGATGCTCT; IL-6 forward, GGGACTGATGCTGGTGACA A and reverse, TCCACGATTTCCCAGAGAACA; Actb forward, GACGTTGACATCCGTAAAG and reverse, CAGTAACAGTCCGCCT. All gene expression values were normalized to Actb (β-actin).
Luciferase assay
Cells were co-transfected with 3×κB-Luc reporter gene plasmids [19] and pNL1.1.TK vector using FuGENE® HD Transfection Reagent (Promega). Twenty-four h after transfection, the cells were incubated with LPS (1 μg/mL) or FPGE (0, 20, 100, or 500 μg/mL). After incubation for 24 h, the media was replaced with luciferase assay solution (Nano-Glo® Dual-Luciferase® Reporter Assay System) following the manufacturer's instructions. The luciferase activity was measured using a GloMax® Discover Multimode Microplate Reader (Promega).
Western blot analysis
Crude extracts were prepared by incubation with PRO-PREP™ Protein Extraction Solution (iNtRON Biotechnology) containing Halt™ phosphatase inhibitor cocktails (Thermo Scientific, Waltham, MA, USA). The protein concentration was determined by BCA assay (Thermo Scientific). The proteins were resolved using 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto a polyvinylidene fluoride membrane. After blocking, the membrane was probed with the indicated antibodies. Antibodies against phospho-NF-κB (pNF-κB) p65 (Ser536), NF-κB p65, phospho-IκBα (Ser32), inhibitor of kappa B (IκBα), phospho-AMPKα (Thr172) and AMPKα were purchased from Cell Signaling Technology (Danvers, MA, USA) Antibodies against phospho-ACC (Ser79), ACC, and β-actin were obtained from Abcam (Cambridge, MA, USA) and horseradish peroxidase-conjugated secondary antibodies were purchased from Promega. The signals were visualized using a WEST-Queen™ RTS Western Blot Detection Kit (iNtRON Biotechnology) and detected by ImageQuant LAS 500 (GE Healthcare Life Sciences, Little Chalfont, UK).
RESULTS
FPGE induces cell proliferation in RAW 264.7 cells
First, we tried to determine the non-cytotoxic concentration of the FPGE. To ascertain whether FPGE affects cell viability, we performed the CCK-8 assay using RAW 264.7 cells. We found that FPGE did not induce cytotoxicity at any concentration (0.8–500 μg/mL); conversely, we found that FPGE treatment led to increased cellular proliferation at high concentrations (100–500 μg/mL; Fig. 1).
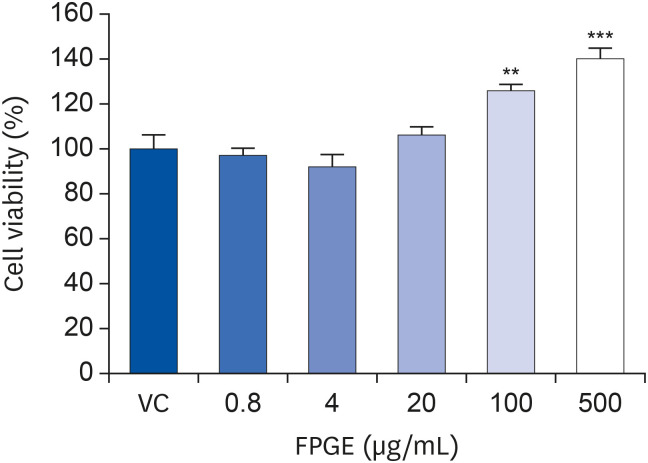
Fig. 1
FPGE induces cell proliferation in RAW 264.7 cells. The RAW 264.7 cells were treated with various concentrations of FPGE for 48 h prior to measure cell viability. Cell viability is expressed as a relative value to that of the vehicle-treated cells, which is set to 100%. The data represents the mean ± SEM (n = 3).
FPGE, fermented Platycodon grandiflorum extract; VC, vehicle control.
**P < 0.01, ***P < 0.001.
![]()
FPGE enhances NO production in RAW 264.7 cells
NO is a crucial physiological messenger in immune responses [20]. To examine the effect of FPGE on NO production in macrophages, we measured nitrite (NO2) level using the Griess Reagent System. Previously, we have shown that LPS induces NO production 18 to 24 h after treatment in RAW 264.7 cells [2122]. In this study, we also confirmed the induction of NO production by LPS in the macrophage. After treating RAW 264.7 cells with FPGE for 24 h, we observed a slight but statistically significant enhancement in the production of NO at the highest concentration (500 μg/mL) of FPGE (Fig. 2).
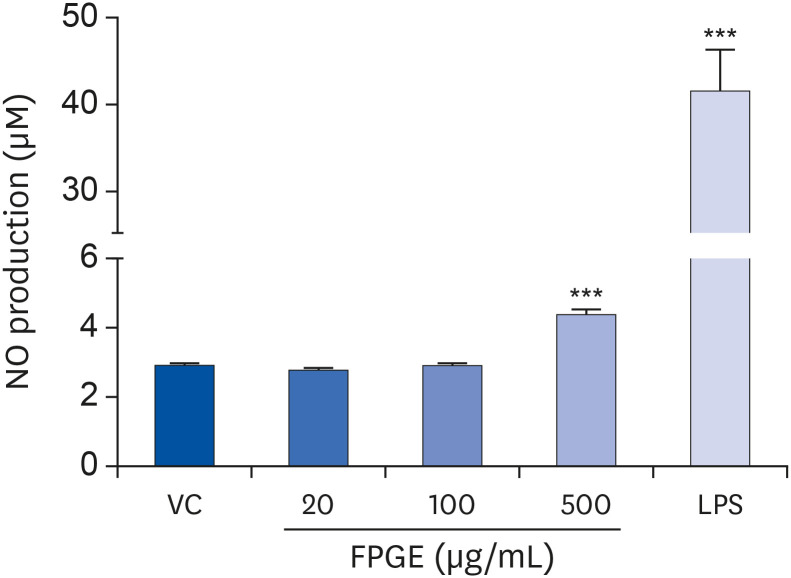
Fig. 2
FPGE enhances NO production in RAW 264.7 cells. Cells were treated with various concentrations of FPGE or LPS (1 μg/mL) for 24 h prior to measure nitrite level. The data were expressed as the mean ± SEM (n = 3).
NO, nitric oxide; VC, vehicle control; FPGE, fermented Platycodon grandiflorum extract; LPS, lipopolysaccharides.
***P < 0.001 as compared with VC.
![]()
FPGE increases the expression of pro-inflammatory cytokines in RAW 264.7 cells
It is well known that activated macrophages stimulate the production of innate immune-related factors such as TNF-α, IL-1β, and IL-6 [8]. For example, after treatment with LPS as a positive control, we observed increased messenger RNA expression of Tnf-α, IL-1b, and IL-6 (Fig. 3). To assess whether macrophages were similarly activated by FPGE, we treated RAW 264.7 cells with various concentrations of FPGE and measured messenger RNA levels of pro-inflammatory cytokines using real-time RT-PCR. Compared to LPS treatment, the effect of FPGE on cytokine levels was less pronounced, but statistically significant compared with the vehicle control at high concentrations (Fig. 3).
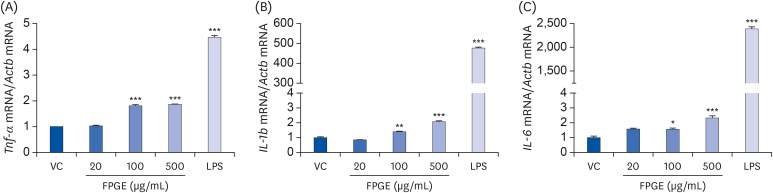
Fig. 3
FPGE increases the mRNA level of pro-inflammatory cytokines in RAW 264.7 cells. RAW 264.7 cells were treated with FPGE (20, 100, 500 μg/mL) or LPS (1 μg/mL) for 24 h and assessed using real-time RT-PCR for (A) Tnf-α, (B) IL-1b, (C) IL-6. All gene expression values were normalized to those of Actb (β-actin) and were expressed as a relative value to that of the VC, which is set to 1. The figures show the mean ± SEM (n = 3).
Tnf-α, tumor necrosis factor-α; mRNA, messenger RNA; VC, vehicle control; LPS, lipopolysaccharides; FPGE, fermented Platycodon grandiflorum extract; IL, interleukin; RT-PCR, reverse transcription polymerase chain reaction.
*P < 0.05, **P < 0.01, ***P < 0.001.
![]()
FPGE induces NF-κB activation in RAW 264.7 cells
To explore the molecular mechanism underlying the immune-enhancing properties of FPGE, we examined the effect of FPGE on NF-κB, a key transcription factor that modulates activation of the immune system. Aligning with prior studies [11], we observed increased NF-κB promoter activity by LPS in RAW 264.7 cells, transfected with appropriate reporter constructs. The treatment with FPGE at 500 µg/mL also showed the induction of luciferase activity (Fig. 4A). To further confirm the effect of FPGE on NF-κB activation, we assessed the phosphorylation status of multiple components in the NF-κB pathway by western blot. As a positive control, LPS increased the levels of pNF-κB and IκBα (Fig. 4B); under the same conditions, phosphorylation levels of NF-κB were increased by FPGE in a dose dependent manner. Phosphorylation levels of IκBα were slightly increased at the highest concentration (500 μg/mL) of FPGE. Therefore, these results demonstrate that FPGE can activate macrophages by NF-κB signaling.
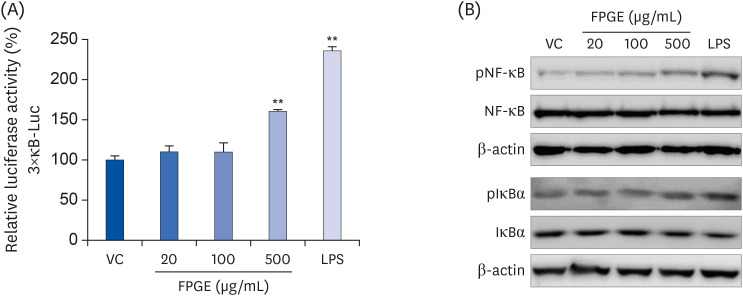
Fig. 4
FPGE induces NF-κB activation in RAW 264.7 cells. (A) The cells were co-transfected with 3×κB-Luc and pNL1.1.TK constructs and treated with FPGE (20, 100, 500 μg/mL) or LPS (1 μg/mL) for 24 h prior to measure luciferase activities. Luciferase activity is expressed as a relative value to that of the VC, which is set to 100%. The figures show the mean ± SEM (n = 3). (B) The cells were treated with FPGE (20, 100, 500 μg/mL) or LPS (1 μg/mL) for 1 h prior to western blot analysis. Western blots are representative of 3 independent experiments.
VC, vehicle control; FPGE, fermented Platycodon grandiflorum extract; LPS, lipopolysaccharides; pNF-κB, phospho-nuclear factor kappa B; NF-κB, nuclear factor kappa B; pIκBα, phospho-inhibitor of kappa B; IκBα, inhibitor of kappa B.
**P < 0.01.
![]()
FPGE-induced NF-κB activation is associated with AMPK in RAW 264.7 cells
Because AMPK may act as an inhibitor of NF-κB in the immune cells, we have questioned whether AMPK signaling acts on LPS- or FPGE-induced NF-κB activation in macrophage cells. When we compared phosphorylation levels to total protein levels of AMPK and ACC, LPS was shown to inhibit AMPK signaling as expected (Fig. 5A-C). There was no significant difference in phosphorylation levels of AMPK or ACC after FPGE treatment in RAW 264.7 cells, although the ratios of phosphorylated to total AMPK and ACC tended to decrease (Fig. 5A-C). To further examine the effect of AMPK on NF-κB signaling, which remains poorly understood, we performed NF-κB reporter gene analysis after treatment with a known AMPK inhibitor (compound C) or an activator (AICAR) in addition to our standard LPS or FPGE treatment. We found that the NF-κB reporter activity increased after treatment with compound C, and decreased after treatment with AICAR. Furthermore, AICAR significantly suppressed the NF-κB activity elevated by LPS and FPGE (Fig. 5D). These results suggest that AMPK inhibition is associated with the NF-κB transcriptional activation induced by FPGE treatment.
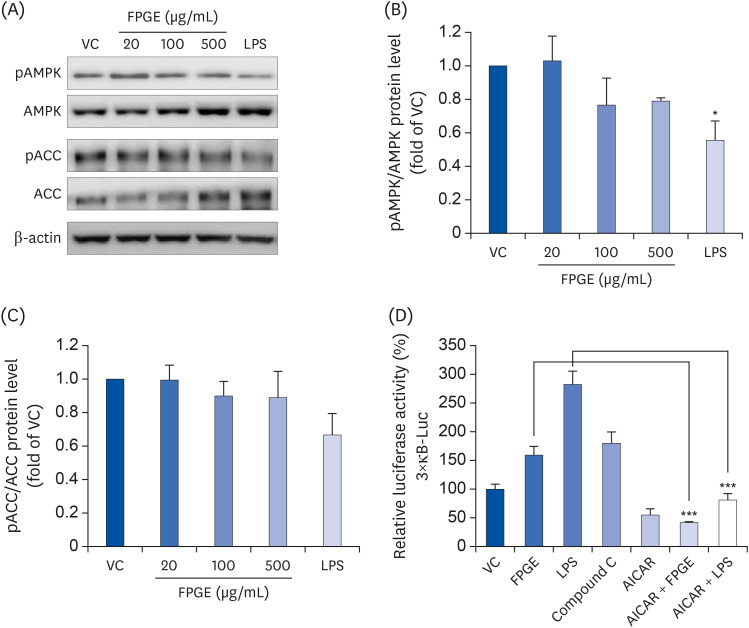
Fig. 5
FPGE-induced NF-κB activation is associated with AMPK in RAW 264.7 cells. (A) The cells were treated with FPGE (20, 100, 500 μg/mL) or LPS (1 μg/mL) or 1 h prior to western blot analysis. The phosphorylated/total ratios of (B) AMPK and (C) ACC were plotted based on the quantification of the signal intensities. (D) The cells were co-transfected with 3×κB-Luc and pNL1.1.TK constructs and treated with FPGE (500 μg/mL), LPS (1 μg/mL), compound C (10 μM), and/or AICAR (1 mM) for 24 h prior to measure luciferase activities. Luciferase activity is expressed as a relative value to that of the VC, which is set to 100%. The figures show the results from 3 independent experiments which were expressed as the mean ± SEM.
FPGE, fermented Platycodon grandiflorum extract; VC, vehicle control; LPS, lipopolysaccharides; pAMPK, phospho-5′ adenosine monophosphate-activated protein kinase; AMPK, 5′ adenosine monophosphate-activated protein kinase; pACC, phospho-acetyl-CoA carboxylase; ACC, acetyl-CoA carboxylase; AICAR, 5-aminoimidazole-4-carboxamide ribonucleotide.
*P <0.05, ***P < 0.001 as compared with VC.
![]()
DISCUSSION
Herbal immunostimulants have been used as immune-enhancing agents, but there are very few products whose effects have been validated scientifically. Despite its prolonged medicinal use and suspected health benefits, PG is produced in limited areas such as Korea, Japan, and China, making it difficult to find overseas research status.
Fermentation process increased the platycodin D content in PG extract [18]. The immune-related effects of platycodin D have been reported to have anti-inflammatory and immunoadjuvant activity [1623]. To the best of our knowledge, this is the first study to investigate immune-enhancing effects of FPGE in a macrophage cell. We demonstrated that FPGE induced macrophage cell proliferation in a dose-dependent manner. Unlike LPS-induced macrophage proliferation, which accompanies morphological change (i.e. stretching-out shape like dendritic cells) [4], FPGE appears to increase the number of cells without morphological change.
Many medicinal plant extracts with immune-enhancing effects modulate at least one pro-inflammatory cytokine, including TNF-α, IL-1, and IL-6 [24]. Our real-time RT-PCR results revealed that FPGE also modulates the expression of these pro-inflammatory cytokines. Thus, we confirmed that FPGE could act as an immunostimulant.
Interestingly, certain natural product such as β-glucan and ginseng displayed both immune-enhancing and anti-inflammatory properties in RAW 264.7 cells [252628]. These indicate that natural product could be effective immune modulators, although the mechanisms are not clearly understood. In this study, we showed that FPGE activates NF-κB signaling, which may be an important underlying mechanism of FPGE-induced production of pro-inflammatory cytokines. Although NF-κB enhances immunity, it also causes inflammation [6]. To investigate the effect of FPGE in severe conditions such as LPS-induced inflammation, we co-treated macrophage cells with FPGE and LPS. We observed that FPGE markedly suppressed NF-κB luciferase activity (Supplementary Fig. 1) induced by LPS, supporting previous research that FPGE has anti-inflammatory effects [18].
PG or ingredients derived from PG are known to increase the AMPK signaling in several cell types, such as hepatocytes [14] and adipocytes [29], as well as in mouse models [30]. However, in our experimental system, FPGE showed a tendency to reduce the signaling of AMPK in RAW 264.7 cells. These observations suggest that cellular response to FPGE treatment may be cell type-dependent.
As the role of AMPK in NF-κB signaling remains controversial in immune cells, we utilized various chemical modulators of AMPK and assessed their effects on NK-κB activity using the luciferase reporter system. Inhibition of AMPK by compound C, a well-known AMPK inhibitor, significantly promoted NF-κB reporter gene activity. We also showed that the efficacy of FPGE is comparable with compound C (Fig. 5D). Therefore, these data suggest that FPGE can be a promising immune-enhancing agent stimulating NF-κB signaling.
In conclusion, our data demonstrates that FPGE induces the production of both NO and pro-inflammatory cytokines in RAW 264.7 murine macrophages. In addition, we show that FPGE has immune-enhancing effects via AMPK-mediated NF-κB signaling. Further studies in animal model and humans are needed to confirm the immune-enhancing effect of the fermented PG root extract as demonstrated in our study.
 XML Download
XML Download