

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Aging is characterized by a progressive decline in physiological function and integrity [1]. Similar to most organs which deteriorate and accumulate deleterious changes with aging, the brain also becomes impaired in regard to its structure, function, and neurocognition with aging [23]. An aged brain is characterized by fewer neurons, hippocampus atrophy, and cognitive decline [3]. Correspondingly, cognitive and memory impairments are common clinical characteristics of brain aging, and are also key factors for maintaining independence in daily activities and overall quality of life in older adults [4]. In particular, the cerebral cortex and hippocampus are the most relevant regions of the brain for aging. These regions undergo changes in structure, weight, and thickness [5]. These regions are also responsible for long-term and short-term memory, physical performance, and cognition [56]. Both brain volume and weight decrease with aging. In addition, neuronal generation is rapidly diminished [78].
D-galactose (Dg) is a reducing sugar which naturally exists in the body. Foods such as dairy products and soy sauce also contain Dg [9]. However, the presence of excess Dg can induce inflammation, oxidative stress, and apoptosis in neuronal cells [1011]. Thus, chronic or high-dose exposure to Dg either in vitro or in vivo is commonly used to induce artificial senescence and brain damage. Markers of this damage include the senescence-related markers, p16, p21, and p53, as well as senescence-associated β-galactosidase (SA-β-gal) staining [31213]. Furthermore, exposure to Dg can enhance mitochondria dysfunction, inflammation, oxidative stress, and apoptosis to cause memory decline and aging [314].
Over the past few decades, it has been demonstrated that plant-based polyphenols can mediate delay in senescence by regulating oxidative stress. For example, anti-aging effects of polyphenols such as anthocyanin, isoflavone, and caffeine have been verified in Dg-induced aged models [151617]. In Asia, curcumin (Cur) is a polyphenolic non-flavone compound derived from turmeric (Curcuma longa), and it is broadly used as both a food spice and an herbal medicine [18]. Cur has been associated with several biological benefits, including anti-inflammation, anti-cancer, and anti-aging properties [19]. These anti-aging and longevity effects of Cur have been elucidated in various experimental models, including Caenorhabditis elegans, fruit fly, and mouse [202122]. However, despite these significant beneficial effects, Cur is characterized by low bioavailability and low solubility. Thus, it has been proposed that encapsulation or a combination administration can improve absorption [232425].
Hesperetin (Hes), a hesperidin aglycone, is a polyphenolic flavanone which can be extracted from citrus fruit [2627]. It has been reported that Hes mediates antioxidant and neuroprotective effects both in vitro and in vivo [2628]. However, the ability of Cur and Hes to improve Dg-induced brain aging has not been elucidated. Therefore, the objective of this study was to investigate whether Cur, Hes, and/or their combination exert anti-aging effects in Dg-induced aged neuronal cells and rats.
MATERIALS AND METHODS
Cell culture and reagents
SH-SY5Y (SY5Y) cells were purchased from American Type Culture Collection (Manassas, VA, USA) and cultured in a 1:1 mixture of Minimum Essential Medium and Ham's F-12 (Welgene, Daegu, Korea) supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA), 1% penicillin (100 U/mL), and streptomycin (100 μg/mL) (Invitrogen, Carlsbad, CA, USA). Cells were maintained at 37°C with 5% CO2 atmosphere.
All-trans-retinoic acid (RA), Cur, Hes, and Dg were purchased from Sigma-Aldrich (St. Louis, MO, USA). RA, Cur, and Hes were dissolved in dimethyl sulfoxide (DMSO). SY5Y cells were treated with RA (10 μM) for 6 days to induce differentiation. Media containing 5% FBS was subsequently used to minimize a serum effect. Due to the light sensitivity of the compounds used, all cell culture experiments were performed under dim light.
Cell proliferation assays
Cell proliferation was evaluated in 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) assays. Briefly, SY5Y cells were seeded in 96-well plates (6 × 104 cells/well) for 24 h and then were differentiated with RA. The cells were then pretreated with various doses of Cur and Hes for 24 h before treatment with 300 mM Dg for 3 days. Treated cells were subsequently incubated in medium containing MTT solution for 3 h at 37°C. After the medium was removed, 150 μL DMSO was added. Optical density was measured for each well at 560 nm in a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Morphological changes in differentiated SY5Y cells
To evaluate neurite outgrowth after neuronal cell differentiation, a low density of SY5Y cells were seeded on 6-well plates and differentiated with RA for 6 days. The cells were subsequently treated with Cur, Hes, or their combination for 24 h, followed by Dg to induce cellular senescence. After 3 days, cell images were obtained at 100× magnification (Olympus, Tokyo, Japan). Neurite length was measured with ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Assessment of cellular senescence
SA-β-gal assays were performed as previously described [12]. Briefly, differentiated SH-SY5Y cells were pre-treated with 1 μM Cur, 1 μM Hes, or their combination for 24 h, Dg (300 mM) was subsequently added. After 3 days, cells were washed with phosphate buffered saline and fixed with formaldehyde for 15 min. After incubation with SA-β-gal staining solution (Cell Signaling Technology, Danvers, MA, USA) for 24 h at 37°C, stained cells were imaged at 100× magnification (Olympus). The numbers of positively stained cells were counted and the percentage of positive cells was calculated.
Protein extraction and Western blot assays
Western blot assays were performed as previously described [12]. Briefly, cell cytosol or frozen brain tissue proteins were extracted with protein extraction solution (PRO-PREP; iNtRON Biotechnology, Seongnam, Korea). These extracts were then separated in sodium dodecyl sulfate polyacrylamide gels and transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). After the membranes were blocked with 5% bovine serum albumin or non-fat dry milk in Tris-buffered saline (pH 7.4) containing 0.1% Tween-20 (TBS-T) at room temperature (RT) for 1 h, they were incubated with primary antibodies diluted in blocking solution at 4°C overnight. The membranes were subsequently washed with TBS-T several times and then incubated with appropriate horseradish peroxidase-conjugated secondary antibodies at RT. After 1 h, the membranes were incubated with enhanced chemiluminescence solution (Visual Protein, Taipei, Taiwan) to detect and visualize bound antibodies with X-ray film (Agfa, Mortsel, Belgium). The primary antibodies were used for Western blot as follows: β-tubulin III, neurofilament heavy polypeptide (NEFH), and α-tubulin from Sigma Aldrich; superoxide dismutase (SOD) 1, catalase, Bax, and Bcl-2 from Santa Cruz Biotechnology (Dallas, TX, USA); glutathione peroxidase (Gpx) 1 from Abfrontier (Seoul, Korea); phosphorylated extracellular signal-regulated kinases (ERK) and poly (ADP-ribose) polymerase (PARP) from Cell Signaling Technology; p16, p21, and β-actin from Abcam (Cambridge, MA, USA).
Animal treatment
Male Wistar rats (10 weeks old, 290–310 g) were purchased from Central Lab Animal Inc. (Seoul, Korea). All of the animals were maintained under controlled temperature (22 ± 2°C) and humidity (50 ± 5%) conditions with a 12 h/12 h light/dark cycle. All of the animals received an AIN-93G diet (Raon Bio, Seoul, Korea) and sterile water ad libitum during the experimental period. After a 7 days adaptation period, the animals were randomly divided into 5 groups as follows: control (Ctrl) group (n = 10), 250 mg/kg body weight (b.w.) Dg-induced aging (n = 10), 250 mg/kg b.w. Dg with 50 mg/kg Cur (n = 10), 250 mg/kg b.w. Dg with 50 mg/kg b.w Hes (n = 9), and 250 mg/kg b.w. Dg in combination with 50 mg/kg b.w. Cur and 50 mg/kg b.w. Hes (Combi, n = 10). The Ctrl and Dg groups received 1.5% carboxyl methylcellulose (CMC, Sigma Aldrich) orally as a vehicle. Animals in the Cur, Hes, and Combi groups were administered each compound mixed with 1.5% CMC orally. Aging was induced by subcutaneous injection of 250 mg/kg b.w. Dg daily for 3 months. Control animals were injected with 0.9% saline. Body weight and food intake were recorded twice a week and supplements were freshly prepared based on body weight prior to each administration.
Passive-avoidance test
Passive avoidance tests were performed by using a shuttle box apparatus (Jeong do BNP, Seoul, Korea). This apparatus is divided into 2 parts: an illuminated room and a dark room which are separated by a guillotine door. The behavior test was performed as previously described with modification [293031]. Before the rats were trained, each of the animals was put into the apparatus and allowed to move freely between the 2 rooms for 5–10 min. This was conducted for the last 2 days of the treatment in order to adapt the animals to the apparatus. On the following day, an acquisition trial was performed. Briefly, each rat was gently held with their body placed on the illuminated room, not faced the guillotine door. When the rats turned their body to the door, the door was opened to let the animals enter the dark room. The initial latency time of entrance of the animals to the dark room from the time of the door opened was recorded. Once the entire body and tail of the animal entered the dark room, the door was closed and an electronic shock (0.6 mA, 5 sec) was delivered when all paws were on the grid. The animals were subsequently returned to the illuminated room. Twenty-four h later, the animals were placed in the illuminated room and latency was recorded to check retention time. Each rat was given a latency time up to 3 min.
Statistical analysis
GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA, USA) was used to perform statistical analyses. One-way analysis of variance with Newman-Keuls multiple comparison test was used to identify significant differences between groups. Data are presented as the mean ± standard error of the mean. P-values < 0.05 were considered statistically significant.
RESULTS
Effects of Cur, Hes, and their combination on cell proliferation
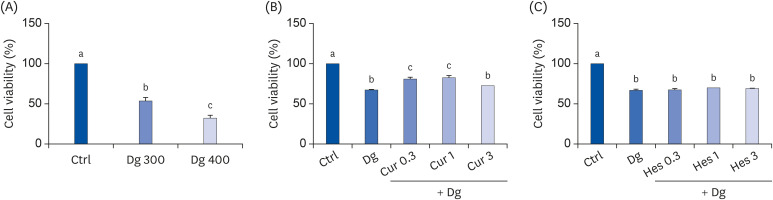
To identify an optimal dose of Dg which affects proliferation of SY5Y cells, MTT assays were performed. Following treatment with 300 mM D-gal and 400 mM D-gal, cell proliferation decreased by 53% and 33%, respectively, compared to the Ctrl group (Fig. 1A). Thus, in subsequent experiments, 300 mM D-gal was used to induce aging. MTT assays with SY5Y cells were also performed to determine appropriate doses of Cur and Hes. Treatment with 0.3 μM and 1 μM Cur increased cell viability by 19% and 22%, respectively, compared with the D-gal group (Fig. 1B). In contrast, cell proliferation was largely unchanged following treatment with Hes (Fig. 1C). Thus, a dose of 1 μM was used for both Cur and Hes in subsequent experiments.
Fig. 1
Effects of Cur, Hes, and their combination on cell viability and proliferation. MTT assays were performed to evaluate cell viability and proliferation. (A) SY5Y cells were treated with Dg (300 mM or 400 mM) for 48 h. (B, C) SY5Y cells were pretreated with Cur or Hes (0.3, 1, or 3 μM) for 24 h, and then were treated with 300 mM Dg for 72 h. Data are represented as the mean ± SEM. Letters label the bars which represent significant differences (P < 0.05).
Ctrl, control; Dg, D-galactose; Cur, curcumin; Hes, hesperetin; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide; SEM, standard error of the mean.
Effects of Cur, Hes, and their combination on D-gal-induced neuronal degeneration
To investigate changes in neurite growth induced by D-gal in SY5Y cells, average neurite length was analyzed (Fig. 2A). Treatment with D-gal significantly reduced the average neurite length compared to the Ctrl group (P < 0.001). Meanwhile, treatment with Cur, Hes, or their combination significantly rescued the average neurite length by 56%, 46%, and 69%, respectively, compared to the D-gal group (P < 0.001 for all).
Fig. 2
Effect of Cur (1 μM) and Hes (1 μM) on synaptic loss and neuronal degeneration in Dg-induced aged SH-SY5Y cells. (A) Representative images (100× magnification) are shown. Neurite length was measured by Image J software. (a) Ctrl; (b) D-gal; (c) Cur; (d) Hes; (e) Combi; (f) Quantified average neurite length from each group. (B, C) The protein expressions of neuronal markers, β-tubulin III, p-ERK, and NEFH were analyzed by Western blot assay. α-tubulin was used as a loading control. (B) Representative blots, (C) quantified proteins levels. Data are represented as the mean ± SEM. Letters label the bars which represent significant differences (P < 0.05).
Ctrl, control; Dg, D-galactose; Cur, curcumin; Hes, hesperetin; Combi, curcumin + hesperetin; ERK, extracellular signal-regulated kinases; NEFH, neurofilament heavy polypeptide; SEM, standard error of the mean.
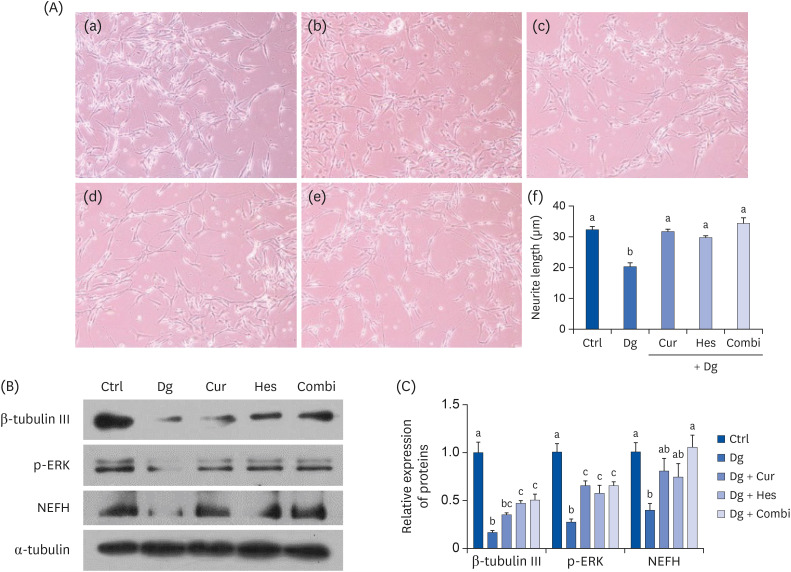
Next, we examined neuronal cell differentiation and neuro-axonal damage-associated markers, β-tubulin III, p-ERK, and NEFH (Fig. 2B and C). D-gal treatment significantly down-regulated expression of all 3 markers compared to the Ctrl group (P < 0.001 for β-tubulin III and p-ERK, P < 0.01 for NEFH). In contrast, Cur treatment restored expression of p-ERK and NEFH, while Hes increased the expression of β-tubulin III and p-ERK. A combination treatment with both Cur and Hes rescued the expression of all 3 markers.
Effects of Cur, Hes, and their combination on D-gal-induced cellular senescence
To analyze cellular senescence induced by D-gal in SY5Y cells, SA-β-gal staining assays were performed (Fig. 3A). In these assays, D-gal treatment significantly increased the number of SA-β-gal positive cells compared with the Ctrl group (P < 0.001). In contrast, treatment with Cur, Hes, or their combination markedly reduced the number of SA-β-gal positive cells by 79%, 82%, and 88%, respectively, compared with the D-gal treatment group (P < 0.001 for all).
Fig. 3
Effect of Cur (1 μM) and Hes (1 μM) on cellular senescence in Dg-induced aged SH-SY5Y cells. (A) Representative images (100× magnification) are shown. Cellular senescence was examined by SA-β-gal staining. (a) Ctrl; (b) D-gal; (c) Cur; (d) Hes; (e) Combi; (f) Quantification of β-galactosidase stained cells. (B, C) The protein expressions of cellular senescence markers, p16 and p21 were analyzed by Western blot assay. α-tubulin was used as a loading control. (B) Representative blots, (C) quantified proteins level data are represented as the mean ± SEM. Letters label the bars which represent significant differences (P < 0.05).
Ctrl, control; Dg, D-galactose; Cur, curcumin; Hes, hesperetin; Combi, curcumin + hesperetin; SEM, standard error of the mean.
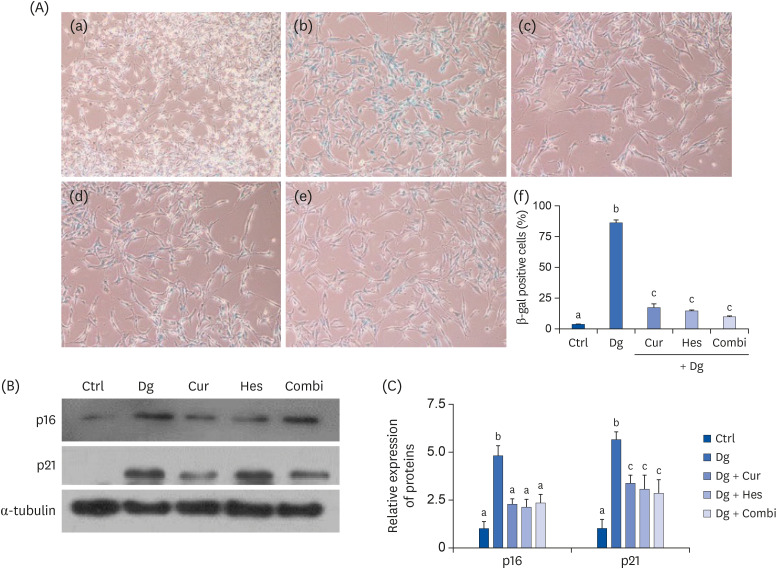
Expression of the cellular senescence-associated markers, p16 and p21, were also investigated by Western blot (Fig. 3B and C). Consistent with the SA-β-gal staining results, D-gal treatment appeared to significantly up-regulate expression of p16 compared with the Ctrl group (P < 0.001). Meanwhile, treatment with Cur, Hes, or their combination significantly attenuated the levels of p16 by 52%, 55%, and 51%, respectively, compared with the D-gal treatment group (P < 0.01 for all). Similarly, expression of p21 significantly increased in the D-gal treatment group compared to the Ctrl group, while expression of p21 was reduced in the Cur, Hes, and Combi groups (P < 0.01 for all).
Effects of Cur, Hes, and their combination on D-gal-induced oxidative enzyme expression
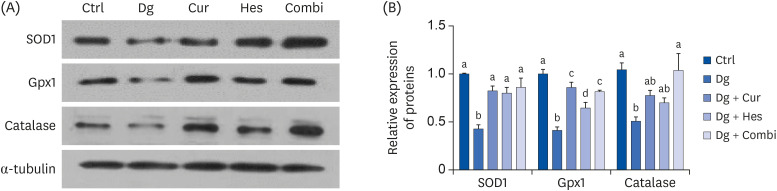
Since oxidative stress is a cause of aging and the mechanism of Dg, expressions levels of the antioxidant enzymes, SOD1, Gpx1, and catalase, were examined (Fig. 4A and B). The D-gal treatment group exhibited significantly lower expression of SOD1, Gpx1, and catalase compared to the Ctrl group (P < 0.001 for SOD1 and Gpx1, P < 0.05 for catalase). Meanwhile, Cur treatment recovered the expression of SOD1 and Gpx1 to control levels, while Hes attenuated the suppression of SOD1 and Gpx1. Treatment with both Cur and Hes recovered expression of all 3 antioxidant enzymes to control levels.
Fig. 4
Effect of Cur (1 μM) and Hes (1 μM) on the expression of anti-oxidant enzymes in Dg-induced aged SH-SY5Y cells. The protein expressions of anti-oxidant enzymes, SOD1, Gpx1, and catalase were analyzed by Western blot assay. α-tubulin was used as a loading control. (A) Representative blots, (B) quantified proteins level. Data are represented as the mean ± SEM. Letters label the bars which represent significant differences (P < 0.05).
Ctrl, control; Dg, D-galactose; Cur, curcumin; Hes, hesperetin; Combi, curcumin + hesperetin; SOD1, superoxide dismutase 1; Gpx1, glutathione peroxidase 1; SEM, standard error of the mean.
Effects of Cur, Hes, and their combination on passive avoidance tests in D-gal-induced aged rats
During training, there were no significant differences in passive avoidance latency among the 5 experimental groups. However, the D-gal group tended to have a shorter latency time compared with the Ctrl group during the 24-h retention trial test, although the results were not significant (Fig. 5A). Hes supplementation blocked the decrease in latency time compared with the D-gal group (P < 0.05), whereas Cur and the combination treatment tended to recover the latency time, albeit not significantly.
Fig. 5
Effect of Cur and Hes on cognitive impairment and neuronal apoptosis in Dg-induced aged rats. (A) Memory retention was investigated by passive avoidance test in Dg-induced aged rats. Training latency was measured on acquisition day. After 24 h of electric shock, memory retention was recorded to a maximum of 180 sec. (B, C) The apoptosis-associated proteins expressions were analyzed in cerebral cortex of Dg-induced aged rats. β-actin was used as a loading control. (B) Representative blots, (C) quantified protein levels. Data are represented as the mean ± SEM. Letters label the bars which represent significant differences (P < 0.05).
Ctrl, control (n = 10); Dg, 250 mg/kg b.w D-galactose (n=10); Cur, 250 mg D-galactose + 50 mg/kg b.w. curcumin (n=10); Hes, 250 mg/kg b.w. D-galactose + 50 mg hesperetin (n = 9); Combi, 250 mg/kg b.w. D-galactose + 50 mg/kg b.w. curcumin + 50 mg/kg. b.w. hesperetin (n=10); SEM, standard error of the mean.
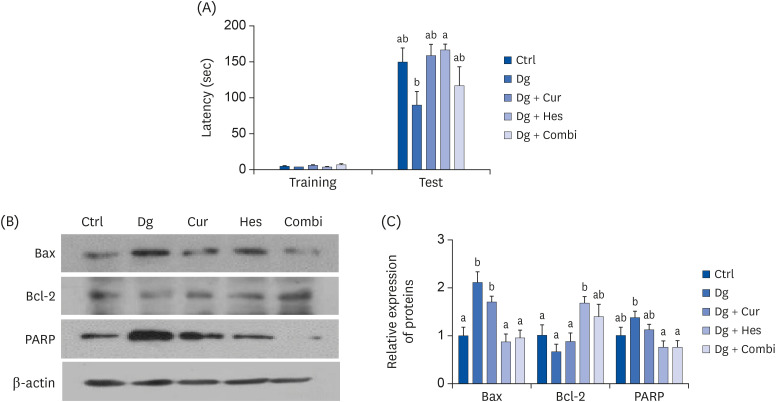
Effects of Cur, Hes, and their combination on apoptosis-related markers in D-gal-induced aged rats
To investigate whether Cur, Hes, or their combination can ameliorate apoptosis in the brain of D-gal treated rats, expression levels of Bax, Bcl-2, and PARP were analyzed in brain cortex tissues (Fig. 5B and C). The expression level of Bax induced by D-gal treatment was recovered to control levels by Hes and the combination treatment. Expression of Bcl-2 was up-regulated by Hes treatment and expression of PARP was inhibited by both Hes and the combination treatment.
DISCUSSION
In the present study, treatment with Cur, Hes, or their combination exerted an anti-brain aging effect by regulating neuron loss, antioxidant enzyme expression, and apoptosis in a Dg-induced brain aging model both in vitro and in vivo. In particular, all 3 treatments improved neuron loss and degeneration, while also inhibited cellular senescence and oxidative stress by upregulating antioxidant enzyme expression in RA-induced SY5Y cells. Correspondingly, our animal study indicated that administration of Cur, Hes, or their combination also protected cognitive impairment and suppressed neuronal apoptosis in the cerebral cortex of Dg-induced brain aged rats.
Dg is widely used to mimic natural aging. Dg is metabolized to glucose and hydrogen peroxide and it turns on the apoptosis pathway when it is chronically or excessively administered. The latter is activated by an accumulation of superoxide anions and free radicals which lead to oxidative stress and aging [332]. We observed in the present study that treatment with 300 mM Dg for 48 h induced cellular senescence in SY5Y cells. Previously, it was reported that Dg treatment efficiently induced senescence not only in animal models, but also in in vitro cell models [123334]. For example, Wang et al. [35] and Rahimi et al. [36] reported that 200–400 mM Dg could cause senescence in RA-induced differentiated SY5Y cells as a result of increased oxidative stress and apoptosis. These results are consistent with those of the present study.
Both dementia and mild Alzheimer disease can be induced by Dg administration. This is a widely employed brain aging model since it exhibits characteristics which are similar to those of aging in the human brain, such as memory decline, motor dysfunction, mitochondria dysfunction, and accumulated oxidative stress [3]. Numerous studies have demonstrated that animal models treated with Dg exhibited declines in spatial learning and memory, decreased antioxidant enzyme activity, and increased apoptosis. As a result, neurodegeneration and brain aging were observed [233738].
Polyphenols possess various biological properties, including anti-cancer, anti-inflammation, antioxidation, and anti-aging properties. For example, resveratrol, which is most abundant in red grapes, has exhibited anti-aging effects by improving age-related memory and mood dysfunction with neurogenesis in older F344 rats [39]. Flavonoid rich orange juice has also been shown to benefit cognitive function when it was consistently consumed by healthy middle-aged men [40]. Cur is a bioactive polyphenol with a long history of use as a therapeutic reagent against inflammation and aging-related disorders [4142]. Anti-aging effects of Cur have been reported in various cell lines, in an animal model, and in a human intervention trial [434445]. In addition, Cur exerted a synergistic effect when it was administered with piperine or hesperidin in Dg-induced aged rats [234647].
Emerging evidence suggests that the role of flavonoids in cognition and memory enhancement is mediated by scavenging reactive oxygen species, activating signaling proteins of the survival pathway, and inhibiting of oxidant-induced neuronal apoptosis [27]. It has recently been demonstrated that 2 flavonoids present in citrus fruit, hesperidin (a flavanone glycoside) and Hes (an aglycone form of hesperidin), exhibit a variety of biological activities, including anti-inflammatory, anti-depressant, and neuroprotective effects [26484950]. For example, hesperidin and Hes both induce neuroprotective effects against H2O2-induced cytotoxicity in PC12 cells and amyloid-induced autophagy in Neuro-2A cells [5152]. However, while both hesperidin and Hes exert similar 2,2-diphenyl-1-picrlhydrazyl radical scavenging activity, Hes has been found to provide better protection from lipid peroxidation in rat brain homogenates and to increase cell viability in primary rat cortical cells [26].
In this study, both Cur and Hes significantly increased neurite length and neuronal marker expression. Neurite outgrowth occurs in the nervous system and also during nerve regeneration [53]. During the latter, neurites grow and connect to other neurites, thereby establish trans neuronal signaling [53]. However, when neurite length is decreased and fewer neurite are present, synaptic degeneration can occur. This is a common characteristic in dementia and Alzheimer disease [54].
In the present study, Cur, Hes, and their combination significantly reduced the number of SA-β-gal-positive Dg-aged cells. When cells are aged by various factors, the cells accumulate lysosomes, and begin to lose their function. SA-β-gal is a biomarker which is widely used to detect aged cells based on its staining of accumulated lysosome [12555657]. A molecular analysis of aged SY5Y cells in the present study showed that Cur, Hes, and their combination were able to down-regulate the senescence-associated proteins, p16 and p21. Moreover, Cur, Hes, and their combination were found to recover antioxidant enzyme expression levels suppressed by Dg. These findings are consistent with those of previous reports which have been showed that Cur or other antioxidant polyphenols, such as caffeine and hesperidin, are able to inhibit oxidative stress and apoptosis related proteins, such as Bax, Bcl-2, and caspase 3 [4658]. Moreover, although hesperidin and Hes are detected at very low concentrations in the brain, they can across the blood brain barrier [59]. An additive effect of Cur and hesperidin in Dg-treated rats has also previously been reported. This combination of supplements improved both cognition and mitochondrial enzymes [46]. Mitochondria play a vital role in the aging progress since oxidative stress mainly occurs in these organelle [4660].
However, the additive effect was not observed in the present study. Although we found that Cur or/and Hes individually rescued Dg-induced brain aging in this study in vitro, combination treatment of Cur and Hes alleviate the brain aging only a few markers. Both polyphenols are rapidly metabolized and excreted in urine or feces [6162]. The absorption of Cur is regulated by the activity of transporter proteins in the luminal membrane of enterocytes. Absorbed Cur is metabolized to a less active form, Cur glucuronides, in the liver. Approximately, 1–2 h after oral ingestion, the plasma level of Cur reaches its highest peak [63]. However, Hes, a glycoside form of hesperidin, is produced by hydrolyzation of hesperidin by colon microflora and a more active form than hesperidin. Hes is detectable 20 min after oral administration of hesperidin, and it reaches its highest peak after 4 h [62]. Since both compounds have different peak times in plasma, they may mediate anti-oxidation effects at different time points. Therefore, if Cur and Hes are administered at regular intervals, additive effects of these 2 polyphenols may be observed. It remains for this hypothesis to be investigated in future studies.
We selected oral supplementation doses based on previous studies [234664] and our preliminary study (unpublished data). Banji et al. [46] reported improved cognitive ability following treatment with Cur (50, 100 mg/kg) and hesperidin (10, 25 mg/kg) in Dg injected rats. Furthermore, in a Morris water maze experiment conducted with D-gal-induced aged rats treated with Cur and hesperidin individually and in combination, a significantly higher number of platform crossings were observed.
Administration of Cur (50, 100 mg/kg b.w.) and Hes (25, 50, 100 mg/kg b.w.) individually, or in combination, were performed in our preliminary study (unpublished data). All of the dosages of Cur and Hes exhibited anti-aging effects. In particular, 50 mg/kg b.w. Cur and both 25 and 50 mg/kg b.w. Hes were associated with improved memorial retention in a passive avoidance test. Based on these results, 50 mg/kg b.w. Cur and 50 mg/kg b.w. Hes were selected as the doses used for oral supplementation in the present study.
Human doses can be extrapolated from animal dosages from calculations of body surface area [65]. By applying the formula, the 50 mg/kg (rat dose) is multiplied by the Km factor (6) for rat and then divided by the Km factor (37) for humans. A 486 mg dose of Cur and/or Hes would be administered to a human weighing 60 kg. It has been demonstrated that up to 8 g/day is a pharmacologically safe dosage range for Cur [66]. The highest intake of Cur to date has been reported in India (100 mg/day) [67]. While the dosage used in the present study is somewhat higher than the dose normally consumed by humans, it still remains less than toxic dose. Moreover, since the bioavailability of Cur between humans and rats differs, the dose administered represents a physiological dose for humans. Glycoside forms are abundant in citrus fruit and juices [68]. Daily intake of a diet rich in vegetables and citrus provides 132 mg/day Hes [69]. Based on these studies, the dose of Hes used in this study represents a physiological dose for humans.
It is well-known that Cur exhibits low bioavailability. Correspondingly, ingestion of single doses of Cur (4, 6, and 8 g) resulted in peak bioavailability 1 to 2 h after ingestion. In addition, average serum concentrations were 0.51 ± 0.11 μM, 0.63 ± 0.06 μM, and 1.77 ± 1.87 μM, respectively. Plasma levels of Cur were also found to increase with long-term administration of Cur over 3 months in humans [70]. Furthermore, no toxicity was associated with the doses of Cur ingested (up to 8 g/day) [66]. Based on these results, in vitro experimental dose of 1 μM is predicted to represent an oral dose of approximately 7 g in humans. While this small amount of Cur remains in the body, Cur can cross the blood brain barrier and may directly affect brain function and aging [71]. There was a lack of evidence regarding the plasma concentration of Hes. Therefore, we are not able to calculate the oral supplementation dose of Hes from the in vitro results.
Based on the present results, we hypothesize that Cur and Hes are able to minimize Dg-induced neuronal aging by regulating antioxidation enzymes and apoptosis-related proteins. However, additional factors contribute to brain aging. These factors may include inflammatory disorders, misfolded proteins, and accumulation of amyloid β plaque. Therefore, it will be important to investigate inflammatory alterations in this model in future studies. In addition, since memory decline which accompanies dementia and Alzheimer disease is governed by both the cerebral cortex and the hippocampus, alterations in the hippocampus should also be investigated.
 XML Download
XML Download