

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Since the past few decades, obesity and its related complications have globally been a topic of great interest. Despite many measures being taken, the prevalence of obesity, mainly in developing countries, continues to increase; it is estimated that by 2025, 18% of men and 21% of women worldwide will be obese [1]. Fat overload in the adipose tissue leads to adipocyte dysfunction, which may stimulate the development of different metabolic complications by affecting glucose and lipid metabolism and increasing inflammatory responses [2]. Chronic conditions such as diabetes, hypertension, cardiovascular diseases, and cancer are associated with obesity, contributing to increases in morbidity and mortality rates and economic burden [3]. The concept of meta-inflammation was introduced to explain the chronic low-grade inflammatory response, which is considered crucial in stimulating obesity-related complications. A characteristic feature of obesity-associated meta-inflammation is that it causes chronic low-grade activation of the innate immune system, eventually affecting metabolic homeostasis [4]. Adipose tissue is a major site of inflammatory responses initiated by macrophage infiltration into the tissue and proinflammatory cytokine production. Activation of the inflammatory nuclear factor kappa B (NF-κB) pathway by endotoxins such as lipopolysaccharide (LPS) via membrane receptor Toll-like receptor 4 (TLR4) in fat cells is among the suggested mechanisms that trigger obesity-associated inflammation [5]. Long-term high-fat diet (HFD) consumption causes a microbial imbalance in the gut, creating suitable conditions for the growth of gram-negative bacteria that produce LPS. This LPS can translocate into the systemic circulation and initiate inflammatory responses. In addition, dietary fat affects intestinal permeability by affecting intestinal tight junction proteins, allowing more LPS to translocate into the blood [6]. The integrity of the gut barrier, which act as a shield protecting the body from various pathogens and toxins, should be maintained so that the intestine functions normally. Changes in dietary pattern and sedentary lifestyle are the major factors contributing to obesity. Hence, including foods with health benefits in our daily diet is the most effective way to maintain health.
Oils are an inevitable component of every cuisine, and oils used in different regions have different fatty acid compositions based on their source, availability, and the processing methods used [7]. Perilla oil (PO) is one of the major oils in Korean cuisine, and it is a rich source of n-3 polyunsaturated fatty acids (PUFAs), particularly alpha-linolenic acid (ALA). ALA acts as the precursor for the synthesis of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which are omega-3-fatty acids containing several health benefits [8]. Olive oil (OO), an inevitable component of the Mediterranean diet, is rich in oleic acid, which is a monounsaturated fatty acid (MUFA) and is known to reduce the risk of cardiovascular disease [9]. Safflower oil (SO) is another widely used cooking oil. Some reports have shown that SO diet reduce fat mass compared to high-butter diet and increase diet-induced thermogenesis compared to a lard diet [10].
Over the past decade, many studies have shown that the type of fatty acid consumed in high concentration, the imbalance between saturated fatty acid (SFA) and unsaturated fatty acids (USFA), and the ratio of omega-6/omega-3 fatty acids can affect susceptibility towards certain diseases [1112]. Though various types of oils are used in our daily life, studies comparing the health effects of oils with different fatty acid compositions are few [1314]. Therefore, based on some previous studies that have reported that PO, OO, and SO have lipid-lowering effects, this study aimed to investigate the effects of these three vegetable oils on HFD-induced obesity and colon inflammation.
MATERIALS AND METHODS
Diet and animals
Commercially available PO, OO, and SO were purchased from a local supermarket. Five-week-old C57BL/6J mice were obtained from Chung-ang Bio (Seoul, Korea). The mice were divided into five groups after one week of acclimatization: LFD (low-fat diet), HFD (high-fat diet), HPO (high-fat diet supplemented with PO), HOO (high-fat diet supplemented with OO), and HSO (high-fat diet supplemented with SO). The major fatty acids present in the oils used in the study were as follows; PO (ALA-59.5%, oleic acid-14.2%, linoleic acid-12.3%, palmitic acid-5.8%), OO (oleic acid-71.2%, palmitic acid-11.51%, linoleic acid-6.15%), and SO (oleic acid-72.4%, linoleic acid-13.6%, palmitic acid-4.7%). The fatty acid composition of the oils used in the study is given in Supplementary Table 1. The experimental diet was prepared weekly. The composition of the diet is presented in Table 1. As this study mainly aimed to investigate the effect of the three vegetable oils which contains unsaturated fatty acids as its major component, the experimental diet was made such that 25% (w/w) of lard in 60% kcal% fat diet (245 g lard in 270 g total fats; Table 1) was replaced with respective oils and the total amount of oil in the diet was 8 g/100 g diet (weight/weight).
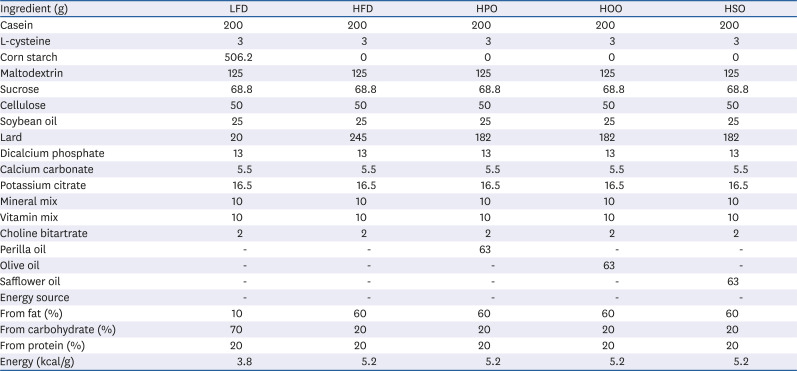
Table 1
Components of the experimental diet fed to the mice
Mice were divided into 5 groups (n = 8); LFD, low-fat diet; HFD, high-fat diet; HPO, high-diet supplemented with perilla oil; HOO, high-fat diet supplemented with olive oil; HSO, high-fat diet supplemented with safflower oil.
![]()
Body weight was measured weekly and diet intake every other day during the 16-week experimental period. The mice were maintained in a housing of 23 ± 1°C, with alternate 12 h light and dark cycles. All the experiments were carried out following the guidelines of the Animal Care and Use Committee of Chungnam National University (CNU-00918).
Micro-computed tomography (micro-CT) analysis for abdominal fat volume
The abdominal fat volume of mice (n=3) was analyzed using micro-CT analysis, one week before sacrifice. Briefly, the mice were anesthetized with Rompun: Zoletil 50 in the ratio 1:1. The abdominal fat region from lumbar vertebrae 1 to 5 was analyzed, micro-CT images were captured, and total fat volume was analyzed by CT analyzer, Skyscan software (Konitch, Belgium).
Biochemical parameters
The mice were sacrificed after 12 h of fasting at the end of the experimental period. The length of the colon was measured (from cecum to rectum) using a fixed ruler. The macroscopic score was assessed according to the scale; 0 - no ulcer or inflammation, 1 - ulceration with local hyperemia, 2 - ulceration without hyperemia, 3 - ulceration and inflammation at one site, 4 - two or more sites with ulceration and inflammation, and 5 - ulceration extending more than 2 cm.
Serum was separated by centrifugation at 1,500 ×g for 15 min at 4°C and was stored at −72°C until analysis. Tissues were immediately frozen in liquid nitrogen and stored at −72°C. Commercially available kits were used to analyze the levels of serum triglyceride (TG), total cholesterol (TC), and high-density lipoprotein-cholesterol (HDL-C; Asan Pharmaceutical Co, Korea). Leptin and adiponectin levels in serum and proinflammatory cytokine such as tumor necrosis factor alpha (TNF-α), interleukin (IL)-1β, and IL-6 levels in the colon were measured using ELISA kits (R&D Systems, Cambridge, MA, USA).
Fecal microbial culture
Approximately 0.1 g of mice feces was collected in sterilized tubes and diluted 10-fold with autoclaved phosphate-buffered saline (PBS). The samples were further serially diluted 10-fold. For analyzing Enterobacteriaceae, the samples were inoculated in desoxycholate agar plates and cultured aerobically for 24 h at 37°C. For Bifidobacteria, the samples were plated onto BL agar plates and cultured anaerobically for 48 h at 37°C. The agar powders were purchased from MB cell, Seoul, Korea.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR) and western blot
To check the expression of genes associated with obesity and colon inflammation, qRT-PCR was performed using liver, epididymal fat, and colon tissue. The mRNA was isolated from liver tissues using the TRI method and from colon using QIAGEN kit. After checking the purity, cDNA was synthesized using the PrimeScript RT master mix and qRT-PCR was performed. Sequences of the primers used in the study are given in Table 2. Inflammatory marker proteins, such as phospho-p65 (p-p65), inducible nitric oxide synthase (iNOS), and cyclooxygenase-2 (COX-2) in the colon, were measured using western blot. Tissues were lysed in Radioimmunoprecipitation assay buffer, centrifuged (4°C, 12,000×g, 15 min), and the supernatant was collected. After checking the protein concentrations, the samples were matched to the same concentration and heated at 95°C after mixing with 5X protein buffer. The proteins were electrophoresed on a 10–12% sodium dodecyl sulfate-polyacrylamide gel and then transferred onto a polyvinylidene difluoride membrane for blotting.
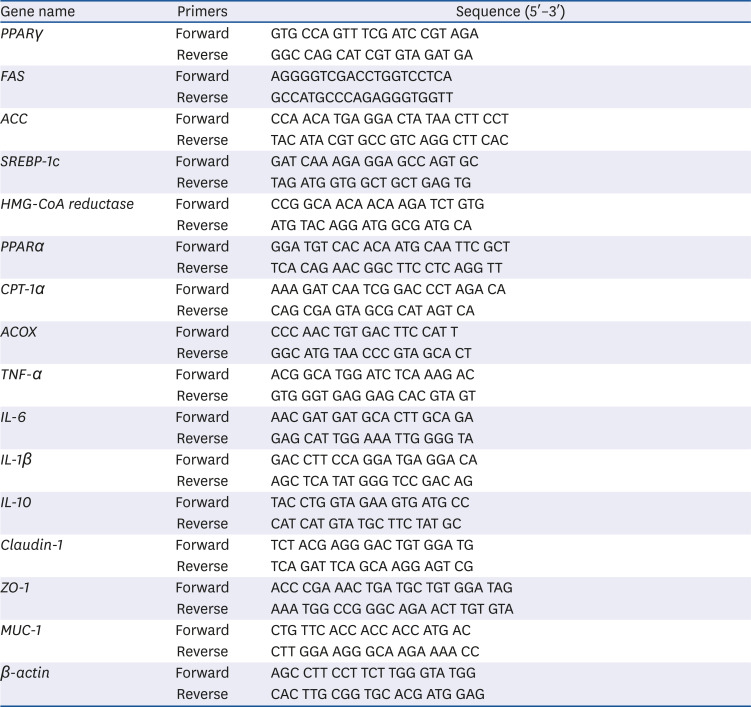
Table 2
Primer sequence of the genes used in the present study
PPAR, peroxisome proliferator-activated receptor; FAS, fatty acid synthase; ACC, acetyl CoA carboxylase; SREBP, sterol regulatory element-binding protein; CPT, carnitine palmytoyl transferase; ACOX, acyl CoA oxidase; TNF, tumor necrosis factor; IL, interleukin; ZO, zonula occludens; MUC, mucin.
![]()
Statistical analysis
Statistical differences were analyzed using one-way Analysis of Variance in SPSS version 17.0 (SPSS Inc., Chicago, IL, USA). Significance was considered at P < 0.05, using Duncan's multiple range test. Values are expressed as mean ± standard deviation and values with different superscripts indicate significant difference between groups.
RESULTS
PO, OO, and SO significantly lowered body weight and organ weight
As shown in Fig. 1, all the HFD-fed groups showed significantly increased body weight compared to the LFD group. However, the three oil-supplemented groups showed significantly lower body weight than the HFD group from the seventh week of study (Fig. 1A) and a similar pattern was seen in the final body weight (Fig. 1B). The liver and epididymal fat weights were also significantly lowered in the oil-supplemented groups (Fig. 1C &and D). Micro-CT results (Fig. 1E) revealed that PO and SO significantly reduced the levels of fat, whereas OO showed a slightly lower abdominal fat mass compared to HFD (Fig. 1F).
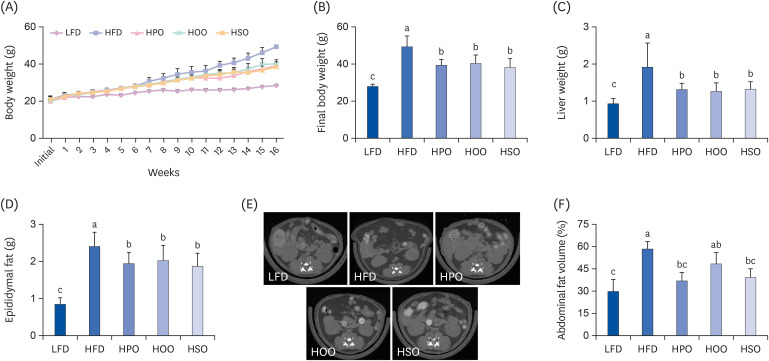
Fig. 1
Anthropometric parameters and micro-CT analysis. Body weight for the whole experimental period (A), final body weight (B), liver weight (C), epididymal fat (D), and (F) percentage of abdominal fat volume. Values are shown as mean ± SD. Values with different superscripts (a, b, c) letters are significantly different among groups. The micro-CT image (E) of abdominal region (dark grey portions indicate fat tissue). Mice were divided into 5 groups (n = 8); low-fat diet (LFD); high-fat diet (HFD); high-diet supplemented with perilla oil (HPO); high-fat diet supplemented with olive oil (HOO); high-fat diet supplemented with safflower oil (HSO).
CT, computed tomography.
![]()
PO, OO, and SO improved obesity-related serum biochemical parameters
As shown in Fig. 2A, serum TG level was significantly reduced in HPO and HSO, whereas the HOO showed a reducing tendency compared to the HFD group. There were no significant differences in the TC and HDL-C levels between the HFD and treatment groups (Fig. 2B and C). As shown in Fig. 2D and E, leptin and adiponectin levels were increased and decreased, respectively, in the HFD group. Interestingly, all the three oil-supplemented groups showed significantly reduced leptin levels. Furthermore, PO and SO significantly increased adiponectin levels whereas OO showed an increasing tendency compared with HFD.
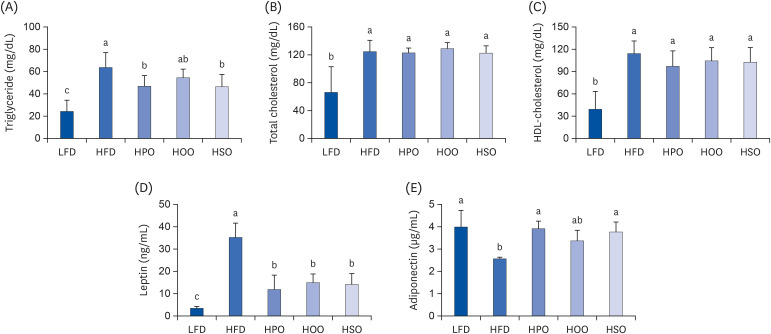
Fig. 2
Serum lipids and adipokine levels. (A)Triglyceride, (B) total cholesterol, (C) high-density lipoprotein-cholesterol, (D) leptin, and (E) adiponectin. Values are shown as mean ± SD. Values with different superscripts (a, b, c) letters are significantly different among groups. Mice were divided into 5 groups (n = 8); low-fat diet (LFD); high-fat diet (HFD); high-diet supplemented with perilla oil (HPO); high-fat diet supplemented with olive oil (HOO); high-fat diet supplemented with safflower oil (HSO).
![]()
PO, OO, and SO improved the gene expression of obesity-related markers in liver and adipose tissue
As shown in Fig. 3, the expression levels of genes associated with lipogenesis such as peroxisome proliferator-activated receptor gamma (PPARγ), fatty acid synthase (FAS), acetyl CoA carboxylase (ACC), sterol regulatory binding protein 1c (SREBP1c), and 3-hydroxy-3-methyl-glutaryl-CoA reductase (HMG-CoA reductase) were significantly increased in the liver and adipose tissue of HFD compared to LFD. Treatment with all three oils reduced the expression levels of these genes. On the other hand, the expression levels of hepatic lipid catabolic genes such as PPARα, carnitine palmitoyl transferase 1 alpha (CPT-1α), and acyl CoA oxidase (ACOX) were significantly upregulated in HPO compared to HFD, HOO, and HSO. Altogether, these results suggest that PO, OO, and SO exert anti-obesity effects by downregulating lipid synthesis and upregulating β-oxidation.
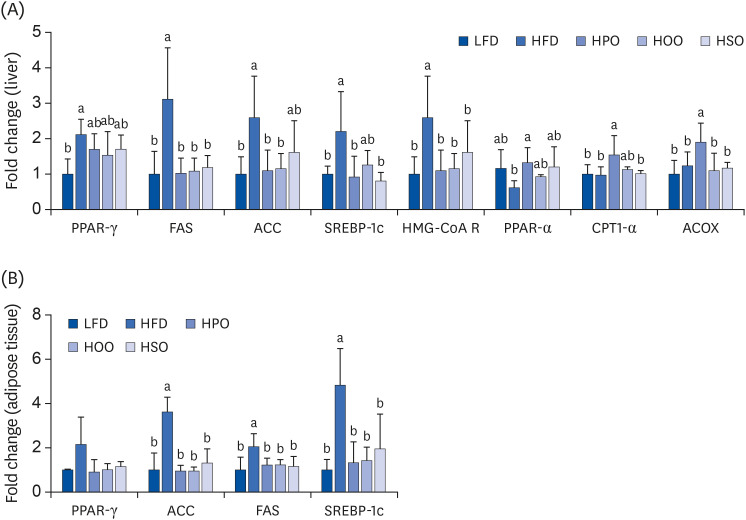
Fig. 3
The mRNA expression levels in (A) liver and (B) adipose tissue. Values are shown as mean ± SD. Values with different superscripts (a, b) letters are significantly different among groups. Mice were divided into 5 groups (n = 8); low-fat diet (LFD); high-fat diet (HFD); high-diet supplemented with perilla oil (HPO); high-fat diet supplemented with olive oil (HOO); high-fat diet supplemented with safflower oil (HSO).
PPAR, peroxisome proliferator-activated receptor; FAS, fatty acid synthase; ACC, acetyl CoA carboxylase; SREBP, sterol regulatory binding protein; HMG-CoA R, 3-hydroxy-3-methyl-glutaryl-CoA reductase; CPT, carnitine palmitoyl transferase; ACOX, acyl CoA oxidase.
![]()
PO, OO, and SO improved colon inflammation
The colon length of the HFD group was significantly reduced compared to that of LFD. However, the colon length of PO, OO, and SO-supplemented groups was higher than that of the HFD (Fig. 4A). Similarly, the macroscopic score was also lower in the treatment groups than in the HFD group (Fig. 4B). Treatment with PO significantly reduced the levels of TNF-α (Fig. 4C), IL-6 (Fig. 4D), and IL-1β in the colon (Fig. 4E). However, OO and SO significantly reduced IL-6 while showing a reducing tendency in the TNF-α and IL-1β levels.
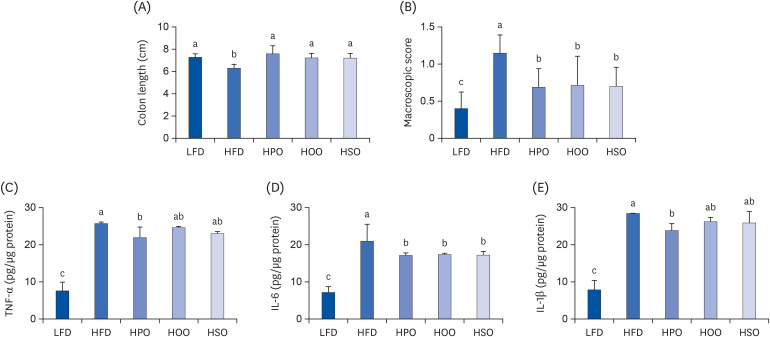
Fig. 4
Inflammation markers in the colon. (A) colon length, (B) macroscopic score, (C) TNF-α, (D) IL-6, and (E) IL-1β. Values are shown as mean ± SD. Values with different superscripts (a, b, c) letters are significantly different among groups. Mice were divided into 5 groups (n = 8); low-fat diet (LFD); high-fat diet (HFD); high-diet supplemented with perilla oil (HPO); high-fat diet supplemented with olive oil (HOO); high-fat diet supplemented with safflower oil (HSO).
TNF-α, tumor necrosis factor alpha; IL, interleukin.
![]()
As shown in Fig. 5A and B, PO significantly increased the number of Bifidobacteria and reduced the number of Enterobacteriaceae. Treatment with OO and SO did not affect the number of Bifidobacteria; instead, the number of Enterobacteriaceae was reduced compared to the HFD group. Endotoxin levels were significantly reduced in all the three oil- supplemented groups, as shown in Fig. 5C.
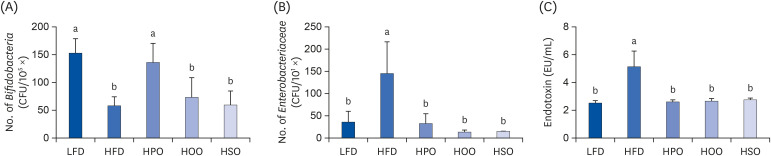
Fig. 5
Number of Bifidobacteria (A) and Enterobacteriaceae (B) in the feces and serum endotoxin levels (C). Values are shown as mean ± SD. Values with different superscripts (a, b) letters are significantly different among groups. Mice were divided into 5 groups (n = 8); low-fat diet (LFD); high-fat diet (HFD); high-diet supplemented with perilla oil (HPO); high-fat diet supplemented with olive oil (HOO); high-fat diet supplemented with safflower oil (HSO).
![]()
As shown in Fig. 6A, the expression levels of proinflammatory markers such as TNF-α, IL-6, and IL-1β in the colon tissue were significantly increased in the HFD compared to that of LFD. Treatment with PO significantly attenuated these increases in proinflammatory marker levels while increasing the expression of the anti-inflammatory marker IL-10. Furthermore, the mRNA expression levels of the tight junction proteins claudin-1 and zonula occludens-1 (ZO-1) were significantly upregulated in the PO-supplemented group. Although not significant, the gene expression of mucin-1 (MUC-1) was higher than the HFD group. The gene expression levels of TNF-α, IL-6, and IL-1β were significantly reduced in the OO and SO-supplemented groups, while the levels of IL-10 were increased. Additionally, the levels of tight junction proteins and MUC-1 showed an increasing tendency in the SO-supplemented group whereas OO did not show any effect on these markers. The protein expressions analyzed using western blot also showed that all the oil-treated groups significantly reduced the levels of p-p65, iNOS and COX-2. Compared to the HFD, HPO showed lower levels of proinflammatory markers, followed by HSO and HOO.
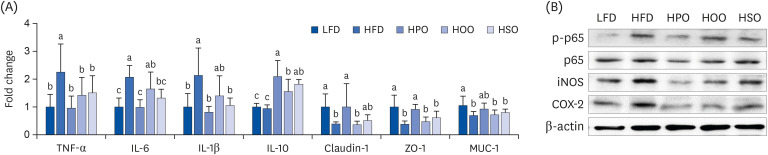
Fig. 6
The expression levels of inflammatory markers in colon. (A) mRNA expression using PCR, (B) protein expression using western blot. Values are shown as mean ± SD. Values with different superscripts (a, b, c) letters are significantly different among groups. Mice were divided into five groups (n=8); low-fat diet (LFD); high-fat diet (HFD); high-diet supplemented with perilla oil (HPO); high-fat diet supplemented with olive oil (HOO); high-fat diet supplemented with safflower oil (HSO).
p-p65, phospho-p65; iNOS, inducible nitric oxide synthase; COX-2, cyclooxygenase-2; TNF-α, tumor necrosis factor alpha; IL, interleukin; ZO, zonula occludens; MUC, mucin.
![]()
DISCUSSION
Obesity-associated complications contribute to increased economic burden in both developed and developing countries. Dietary and behavioral interventions are the preliminary steps toward managing obesity and related diseases [15]. Compared with unsaturated fat diet, saturated fat diet contributes to obesity and hepatic steatosis, and it can increase dietary fat flow to the intestine resulting in changes in the gut microbiota [16]. Reports have shown that replacing saturated fat with PUFA can reduce the risk of cardiovascular disease in humans [17]. Furthermore, long-term intake of omega-3 fatty acids reduces the risk of ulcerative colitis [18]. Oils are an unavoidable part of every cuisine, and each oil differs in its fatty acid composition. In this study, we aimed to investigate the effect of three mainly used dietary oils; PO, OO, and SO in HFD-induced obesity and colon inflammation.
In the present study, compared with HFD, supplementation with PO, OO, and SO improved body weight, organ weights, serum TG, leptin, and adiponectin levels. Interestingly, in the analysis of abdominal volume using micro-CT, HPO and HSO showed more pronounced effects than HOO. In some previous reports, PO has been shown to lower serum TG, TC, and low-density lipoprotein-cholesterol levels [1920]. In the present study, PO lowered serum TG levels without affecting serum TC levels. According to a previous report, OO has hypolipidemic activity [21]. In the present study, compared with HFD, HOO group showed reduced TG levels, however TC level was not significantly different.
High leptin and low adiponectin levels are characteristic of obesity. Adiponectin contributes to β-oxidation by mediating the phosphorylation of AMP-activated protein kinase (AMPK) and activating PPARα [22]. PO suppressed leptin levels and increased adiponectin levels, which might have been reflected in the increased gene expression levels of β-oxidation in the HPO group. The results of our study showed that each of the oils exerts lipid-lowering effects and can improve obesity.
Supplementation with PO downregulated the expression of lipogenic genes, such as PPARγ, FAS, ACC, SREBP1c, and HMG-CoA reductase and upregulated the expression of PPARα, the transcription factor that increases the expression of genes related to β-oxidation. The observed pattern was similar to that of mRNA expression of lipogenic genes in adipose tissue. The results of a previous study also showed that PO improves lipid metabolism in the liver by phosphorylating ACC via activation of the AMPK pathway in HFD-fed mice [23]. Furthermore, Zhang et al. [24] reported that PO induces lipid oxidation in rats by increasing PPARα and CPT-1α. Treatment with OO and SO also showed downregulation of lipogenic genes and showed a tendency to increase β-oxidation-related genes. A previous study showed that high oleic acid SO increase the expression of hepatic PPARα target genes and decrease the expression SREBP-1c, ACC and FAS in rats [10]. Altogether, our results suggest that PO, SO, and OO ameliorate HFD-induced obesity possibly by reducing lipogenesis and increasing β-oxidation.
Many recent researches have shown the association between HFD and low-grade inflammation [252627]. Chronic ingestion of HFD causes changes in the microbial population, which results in increased production of LPS. This in turn increases the production of proinflammatory cytokines such as TNF-α, IL-6, and IL-1β, and if this condition persists, it can lead to damage of the layers of intestinal barrier. Excess LPS is also known to affect the permeability of the intestinal barrier by disrupting tight junction proteins. Translocation of LPS into the systemic circulation can stimulate inflammatory responses in different tissues, and this is the starting point for several complications associated with obesity such as insulin resistance [628]. In the present study, PO, OO, and SO improved the inflammatory conditions by lowering proinflammatory cytokine levels and prevented colon shortening. Interestingly, PO exerted better improvement effects than OO and SO, as evident from the significantly reduced TNF-α and IL-1β levels. PO also increased Bifidobacteria, while OO and SO did not show any effect. However, all three oils lowered the numbered of Enterobacteriaceae, which is responsible for the reduced serum endotoxin level.
The expression levels of genes associated with colon inflammation and tight junction proteins showed that PO exerts a better protective effect than OO and SO. The mRNA expression levels of proinflammatory cytokines were downregulated by PO, OO, and SO supplementation. However, the expression of tight junction proteins, such as claudin-1 and ZO-1, was increased only in HPO compared to HFD. Although HSO showed an increasing tendency in terms of claudin-1 expression, HPO showed more pronounced expression levels. The expression levels of proteins such as p-p65, iNOS, and COX revealed that PO, OO and SO significantly reduced inflammatory markers in the colon compared to HFD. Interestingly, compared with OO and SO, PO showed a markedly higher inhibiting effect on NF-κB activation, as evident by the reduced p-p65 levels. A previous report on a TNBS-induced colitis model showed that ALA suppressed NF-κB activation as well as reduced proinflammatory cytokines and markers of oxidative stress in the colon [29]. ALA being the major compound in PO, although we used HFD-induced colon inflammation mice model, our study results were similar. A recent study on OO showed that it reduced the expression of TNF-α gene expression in the colon; however, although it showed a hypocholesterolemic effect, it did not show protective effect against colon inflammation in HLA-B27 transgenic rats, which spontaneously develop immune-mediated chronic intestinal inflammation [30]. The results of this study partially agree with those of the present study because OO reduced the expression of inflammatory markers in the colon and improved the colon length and macroscopic score. The difference in the mice models used might be a reason for variations in the results.
The fatty acid composition analysis of the three edible oils used in this study showed that PO contained a high amount of ALA (59.7 g/100 g), the precursor of EPA and DHA. Both OO and SO consisted of a high amount of oleic acid (71.2 g and 72.4 g per 100 g oil, respectively). The ratio of SFA: USFA was the same (1:10) in PO and SO. However, the ratio of MUFA: PUFA was higher in SO (5:1) compared to PO (1:5). Though OO and SO have a similar amount of MUFA, the ratio of SFA: USFA and MUFA: PUFA in OO were 1:5 and 10:1, respectively. There are two kinds of SO available in the market, one is high-oleic acid SO and the other is high-linoleic acid SO. Some previous researches have shown that high MUFA containing diets with a low SFA: USFA may ameliorate obesity [3132]. In the present study, we also found that high-oleic acid SO that has a low SFA: USFA ratio showed less fat accumulation compared to OO which has a high SFA: USFA ratio.
To summarize, we investigated the effect of PO, OO and SO, oils with different fatty acid compositions, on HFD-induced obesity and colon inflammation. HFD contains high amounts of saturated fat whereas PO, SO, and OO contain higher amounts of unsaturated fats. Previous reports have shown that a low saturated fat: unsaturated fat ratio is beneficial for health [33]. In our study, we observed that HPO, HOO, and HSO showed lower levels of inflammatory markers, indicating that a diet containing a low ratio of saturated to unsaturated fatty acids is effective in reducing colon inflammation. Furthermore, PO showed a more pronounced effect on the improvement of colon inflammation when compared to OO and SO. These results emphasize the importance of replacing saturated fats with unsaturated fats, especially PUFA-rich perilla oil in consideration of a healthy dietary pattern.
 XML Download
XML Download