

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Osteoporosis is recognized by decreased bone density and a deterioration in bone structure, causing a decrease in bone strength and an increase in bone fragility and fracture problems [1]. Approximately 200 million persons worldwide have osteoporosis-related health complications, with more than 9 million fractures being reported annually. About 20% of men and 33% of women over 50 years of age are at high risk of fractures due to osteoporosis and with the rapid increase in this population worldwide, the incidence of hip fracture will continue to increase significantly in the near future [2]. The reported prevalence of osteoporosis in women was 16% in the United States of America, 15% in France and Germany, 9% in the United Kingdom and 38% in Japan, while in men, the prevalence was 1% in the United Kingdom, 4% in Japan, 3% in Canada, and 8% in France in 2014 [3].
Glucocorticoids (GCs) have been used in the treatment of many diseases for decades to suppress tissue inflammation and immunological reactions. Many patients with diseases of the lungs, joints, skin, muscle, eyes, nerves, etc., take prednisolone to control their disease activity and symptoms. A significant part of the population in the following countries (Belgium, Germany, Netherlands, UK, France, Italy, Spain, USA, Canada and Australia) has been reported to be receiving GC therapy [4]. GCs suppress bone formation, increase bone resorption cause negative calcium balance, induce a high risk of fractures and lead to GC-induced osteoporosis [5]. GCs are considered one of the main culprits of secondary osteoporosis which leads to dangerous fractures and causes significant morbidity. After starting of oral GCs, bone loss occurs rapidly, and fracture risk increases [6].
The omega-3 polyunsaturated fatty acids (ω-3 PUFAs) have been reported to have valuable effects in the regulation of bone metabolism and to be positively correlated with bone mineral density (BMD) [7]. The major ω-3 PUFAs include the essential fatty acid (FA), alpha-linolenic acid (ALA), and its longer chain derivatives, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which are important in the control of osteoblastogenesis and inhibition of bone resorption [8]. A recent review of several micronutrients for improving health status among older people identified ω-3 PUFAs as selected micronutrients that might effectively increase the bone formation rate and skeleton functions [9].
Marine fish is a major source of omega-3 PUFA, Hutchins-Wiese et al. [10] found that fish oil can decrease bone resorption and inhibit osteoclastogenesis, thereby decreasing bone resorption. Some plant oils like flaxseed and soybean are rich sources of PUFAs. The essential FA, alpha-linolenic acid (ALA), is usually consumed from different food sources including nuts and seeds (walnuts and chia seeds) and plant-based oils (flaxseed oil, canola oil, and soybean oil) [7]. El-Saeed et al. [11] investigated the role of flaxseed oil in the homeostasis of bones and its effect on changes in the levels of the bone formation markers. They found that flaxseed oil has beneficial impacts on the prevention of osteoporosis. Hassan et al. [12] concluded that consumption of soybean oil-supplemented diet is considered a functional food for preventing and decreasing the problems of osteoporosis in rats.
The lack of effective treatments for the risk of osteoporosis in GC-treated individuals results in a special focus on prevention strategies, such as using diet to protect bone loss as a side effect for patients being treated with prednisolone. Therefore, the main purpose of the present study was to evaluate the role of soybean, flaxseed, and fish oils in preventing bone loss and osteoporosis in GC-treated rats.
Methods
Fatty acid (FA) analyses of oils
Fish oil (from cod liver) was obtained from Tula pharmaceutical factory, Tula, Russian federation. Soybean and flaxseed oils were purchased from the Food Technology Research Institute, Agriculture Research Center, Giza, Egypt. Samples were stored at −20℃ in dark glass bottles until used. The oil samples were esterified and methylated according to the AOCS Official Method Ce 2-66 (AOCS 2009) [13], Methyl esters of FA were determined by gas chromatography using a Perkin Elmer Auto System XL model supplied with flame ionization detector. The compounds were separated on a fused silica capillary column DB-Wax (60 m × 0.32 mm intern diameter, 0.20 µm film thickness). The temperature of the oven was initially kept at 50℃ for 5 min, then programmed to increase from 50 to 220℃ at a rate of 4℃/min. The carrier gas was helium at a flow rate of 1.1 ml/min. The temperatures of the injector and detector were 220 and 250℃, respectively. FAs were identified by comparing the retention times of pure standard methyl esters of FAs with the separate components of the samples, which were quantified by area normalization (%) [14]. A mixture composed of 37 FA methyl esters was used as a standard (Supelco, Sigma-Aldrich Co. LLC, Germany) with a purity between 99.1 and 99.9%.
Experimental animals
Twelve-week-old male Sprague Dawley rats (135 ± 15 g) were bought from the Laboratory Animal House, National Research Center, Cairo, Egypt. Only male rats were used because of the hormonal fluctuation of female rats during the menstruation cycle, which may have influenced our experiment and results. The rats were housed individually in ventilated cages and maintained in a 12 h light/12 h darkness cycle at 23–25℃, with free access to food and deionized water. This study was approved by the ethical committee of the National Research Center, Egypt (approval number: 4/3/15).
Experimental design and diet
The diet was based on the purified balanced AIN-93GM diet, which was formulated for the maintenance of adult rats according to Reeves et al. [15]. After two weeks of acclimatization, the rats were divided into the following 5 groups (8 rats/group): I - (normal control), which was fed a balanced diet with sunflower oil; II - (prednisolone control) which was fed the balanced diet and administrated with prednisolone (10 mg/kg per daily) [16]; III - soybean oil (S), prednisolone (10 mg/kg per daily) + soybean oil (7% w/w in substitution with sunflower oil in the balanced diet); IV - flaxseed oil (X), prednisolone (10 mg/kg per daily) + flaxseed oil (7% w/w) [11]; and V - fish oil (F), prednisolone (10 mg/kg per daily) + fish oil (7% w/w). The experimental period was 3 weeks. Food intake was monitored daily and body weight was recorded weekly during the experimental period.
Blood assays
After the experimental period, the rats were fasted for 12 h and sacrificed by cervical dislocation. Blood samples were collected in heparinized tubes and centrifuged at 2,500 rpm under cooling. The separated plasma was frozen at −80℃ for future biochemical analyses. The concentration of malondialdehyde (MDA), an index of lipid peroxidation, was measured according to the method of Draper and Hadley [17]. Tumor necrosis factor-alpha (TNF-α) was assayed using an enzyme-linked immunosorbent assay (ELISA) kit purchased from RayBio® Rat TNF-α (RayBiotech, Inc., Norcross, Georgia, USA). Calcium (Ca), phosphorus (P) and magnesium (Mg) values were assessed using colorimetric assay kits (BioSystems S.A., Costa Brava, Barcelona, Spain). 1,25-(OH)2-Vitamin D3 was quantitatively measured in plasma using an ELISA kit (IBL International GmbH, Hamburg, Germany). The C-terminal telopeptide (CTX) was determined using Plasma Cross Laps One-Step ELISA (Osteometer BioTech, Herlev, Denmark). Parathyroid Hormone (PTH) was determined using an ELISA kit (MyBiosource, Inc., San Diego, USA).
Bone examination
The right femur of each rat was anatomized, all tissues covering the bone were removed, and washed in saline. The femur mass was recorded by electronic balance, and the length was measured using an electronic caliper. The BMD of the femurs was determined using dual-energy X-ray absorptiometry (DEXA) (Norland XR-600, Swissray International, Inc., Edison, NJ, USA). For histopathological examination, bones were put in 10% formalin for 48 hours. Decalcification was started by ethylene diamine tetra-acetic acid (EDTA) 10% in 7–7.4 pH for 4 weeks. The tissues were cleared with xylol. Femurs were cut transversely. The cleared fixed tissues were transferred into melted paraffin to form blocks of hard paraffin with tissues in the center. Each block of paraffin was cut into 6 µm-thick sections using a rotatory microtome. Paraffin sections were then transferred to clean slides of glass smeared with glycerin-albumin. The slides were dried and stained by hematoxylin and eosin stain (H & E) for histopathological examination by light microscope. Images were analyzed by Optimas (Media Cybernetics, Inc., USA) 1998 version 6.21.19 [18].
Statistical analysis
The data were statistically analyzed using CoStat Version 6.451 statistical program (CoHort Software, USA). The differences in mean values among all groups were evaluated by one-way completely randomized analysis of variance (ANOVA), followed by Tukey's honestly significant difference (HSD) post hoc test to compare differences in values and determine the significance level (p < 0.05). The results are presented as mean ± standard error (SE).
Results
Fatty acid (FA) profile of oils
As is well known, fish, flaxseed and soybean oils have different compositions of the various FAs. The specific FA profiles in the three study oils were determined and the results are shown in Table 1. Therefore, it was necessary to evaluate which oil is more effective in bone metabolism and in preventing osteoporosis.
Weight gain, food intake, BMD, mass and length of femur
As presented in Table 2, the body weight gain and the daily food intake values showed insignificant changes among the five groups. Femur mineral density, mass and length are also shown in Table 2. Administration of prednisolone in the positive control group significantly reduced femur mass and BMD compared to the negative normal group, and the reduction in femur mass was improved insignificantly by the treatment with oils in all treated groups. Furthermore, the treatment with fish oil significantly increased BMD compared to the prednisolone control group. However, the femur length did not change significantly among the five groups.
Calcium, phosphorus and magnesium in blood
Plasma minerals were estimated (Table 3). The prednisolone control group demonstrated a significant decrease in the plasma levels of calcium and phosphorus compared to the normal group, and this decrease was significantly improved by the supplementation of the three oils in the treated groups. The fish oil group had the highest calcium level among all treatments and showed no significant change compared to the normal control, while the plasma magnesium levels in the five groups didn't show any significant differences.
Biomarkers related to bone metabolism, inflammation and oxidative stress
Table 4 shows the inflammatory marker (TNF-α), oxidative stress marker (MDA), and changes in 1,25-(OH)2-vitamin D3, PTH and CTX. The prednisolone control group recorded the highest values of inflammatory and oxidative stress markers TNF-α and MDA, respectively, while the normal control recorded the lowest values. Treatment with oils significantly improved these values in the treated groups, especially fish oil, which showed a normal value of MDA compared to the normal control group. Supplementation with fish oil also demonstrated the highest value of vitamin D3 with no significant differences from the normal control, while the supplementation with flaxseed oil showed the lowest value. Administration with prednisolone in the control group significantly increased PTH and CTX compared to the normal control, and the PHT level was decreased after treatment with flaxseed oil and fish oil. Furthermore, the CTX levels in all treated groups were significantly improved compared to the prednisolone control group.
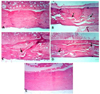
Histological findings of bones
The histological results of femoral examination showed that the normal control (Fig. 1A) and fish oil (Fig. 1E) groups had normal cortical bone thickness and normal osteocyte with no histopathological changes. In contrast, the prednisolone control group (Fig. 1B) clearly demonstrated a significantly thin cortical bone, accompanied by cracks and fissures, abnormal osteocytes, and several resorption cavities within the matrix. On the other hand, only few cracks and fissures were seen in the cortical bone of the soybean oil group (Fig. 1C). There was some focal necrosis with few cracks and fissures in the cortical bone of rats from the flaxseed oil group (Fig. 1D).
Discussion
GCs are absorbed through the cell membrane by special receptors and affect lipid, protein and carbohydrate metabolisms. The impact of GCs on bone could be linked to the imbalance between bone resorption and bone remodeling, vascular occlusion, and activation of apoptotic signaling pathway by some GCs [19]. Ton et al. [20] suggested that taking even low doses of prednisolone significantly decreases bone formation, may inhibit bone remodeling and could have different effects on bone strength. There are few published recommendations for preventing GC-induced osteoporosis. The prednisolone dosage in the present study was based on data from a previous study on rats [16]. Given that systemic metabolism and the biotransformation of drugs are stronger in rats than in humans, a ten-time transformation rate was adopted in biological experiments conducted on rats. Osseous tissues of rats are distinguished by a fast rate of metabolic turnover. Therefore, the three-week study duration was considered sufficient to evaluate the impact of prednisolone on bone loss in rats [21].
Several studies reported that PUFAs and their metabolites play significant roles in controlling bone metabolism. The link between PUFAs and bone metabolism could be related to some suggested mechanisms such as regulation of the activity and differentiation of the osteoclast and osteoblast, as well as changing the FA composition of bone cell membranes [22]. The omega-3 PUFAs have been reported to suppress osteoclast activity and improve osteoblast activity [23]. Dietary supplementation of long-chain PUFAs demonstrated a protective effect against aging-related bone loss [24].
In our study, we found that the main FA detected in flaxseed oil was ω-3 PUFAs α-linolenic acid (ALA), followed by the monounsaturated oleic acid, and these results are in line with some recent research [25]. Furthermore, the major dietary FA found in soybean oil was ω-6 PUFA (linoleic acid) at more than 50%, and this result was similar to Dorni et al. [26] who found that ω-6 polyunsaturated linoleic acid was the highest (more than 50%) in soybean oil, followed by the monounsaturated oleic acid, and then the saturated palmitic acid, which together accounted for more than 90% of the total FAs. On the other hand, sunflower oil is also known to be rich in linoleic acid and oleic acid. Fish oil has been reported to be a good source of long-chain ω-3 PUFAs with significant quantities of EPA and DHA that are considered to be metabolically active compounds [27].
In the present work, the femur mass and BMD were reduced in rats with prednisolone-induced osteoporosis. Kaczmarczyk-Sedlak et al. [16] found that administration of prednisolone 10 mg/kg daily for three weeks significantly reduced caused a significant reduction in femur mass by (11.2%). The ratio of femur mass to body mass was lower in rats with prednisolone-induced osteoporosis than in those from the normal control group. This observed decrease in femur mass could be related to bone loss and bone resorption. The negative effect of GCs on the osseous tissue includes both a reduction in BMD and an increase in the number of bone fractures [28]. Pennisi et al. [29] administered prednisolone (5 mg/kg) for 4 weeks to rats and found a reduction in growth and low mineral density of the vertebrae, hip and femur. In our study, feeding rats with PUFAs with slightly different compositions from three different oils increased bone mass in all treated groups, which is in line with Lukas et al. [30] In the current study, only rats fed with fish oil had a significant improvement in BMD compared to the other treated groups. Sun et al. [31] reported that rats fed fish oil, which has high levels of EPA and DHA, had higher BMD and stronger bone than rats fed soybean oil, which is rich in ALA.
Our evaluation of plasma minerals demonstrated that administration with prednisolone significantly decreased plasma calcium and phosphorus compared to the normal group. Some mechanisms proposed in the literature lead to GC-induced bone loss by increasing renal calcium excretion and reducing the absorption of calcium through the intestinal tract [32]. GCs have many indirect negative effects on bones, including disturbance of calcium-phosphorus homeostasis (through blocking intestinal calcium and phosphorus absorption and increasing urinary calcium elimination), reducing growth hormone, and affecting PTH [33]. Therefore, non-pharmacological treatments such as suitable nutrition for preventing bone resorption are necessary for all adults who are taking prednisolone. Our data showed significantly improved levels of plasma calcium and phosphorus after supplementation with the three oils. Beneficial interactions between calcium and fish oil were observed in the rat model of osteoporosis. These data support the proposed mechanism in which calcium absorption and excretion are regulated after supplementation of fish oil [31]. Omega-3 PUFAs affect bone health by improving calcium balance and bone turnover. The ω-3 PUFAs also influence bone structure by affecting osteoclastic and osteoblastic differentiation and activity [22]. Omega-3 improved plasma calcium levels, and thereby increased bone mass; hence, our data confirm the valuable effect of PUFAs on bone formation. Heaney et al. [34] found that supplementation of ω-3 PUFAs increased calcium absorption in humans. Long-chain PUFAs preserved Ca and P homeostasis and caused no disturbance in their nutritional balance. Soybean oil has been reported to promote bone health through increasing calcium and phosphorus retention [35].
In the current study, the group of rats fed with fish oil had the highest vitamin D3 levels among the five groups. Fish oil is considered a good source of vitamin D3 and is recognized as one of the major sources of vitamin D3 in the human diet [36]. Vitamin D is hydroxylated to 25(OH)2D3 in the liver, followed by conversion to its most potent biological metabolite (1,25-(OH)2D3) by the enzyme 1α-hydroxylase in renal proximal and distal convoluted tubules. An et al. [37] demonstrated that ω-3 PUFAs enhance 1,25-dihydroxyvitamin D level in dialysis patients. One hypothesis is that increased 1,25-(OH)2D3 concentrations were caused by the enzyme 1α-hydroxylase activation by omega-3 PUFAs [38].
In addition to the enhanced bone formation, the treated groups showed a reduction in bone loss markers, as reflected by lowered CTX levels in plasma compared to the prednisolone control. Dong et al. [39] surveyed CTX levels in 164 postmenopausal women and found a significant decrease in CTX level in the intervention group after supplementation with ω-3 PUFA compared to the control group. High levels of PTH play a role in the development of bone loss by transferring bone calcium into the blood to restore the normal plasma calcium levels. Consequently, elevated levels of PTH are associated with increasing bone resorption rate, as seen in our results after prednisolone administration. Varela-López et al. [40] also suggested that PTH is released when plasma calcium levels are low to restore calcium from bone into blood, which induces bone resorption process. On the other hand, fish oil significantly reduced the level of PTH. It has been reported that the protective effect of ω-3 PUFAs against bone loss was due to the modulation of systemic calcitrophic hormones, including PTH [41].
MDA is a secondary substance of lipid peroxidation that is strongly associated with oxidative stress [17]. According to our data, treatment with oils significantly decreased oxidative stress in all treated groups. Han et al. [42] found that flaxseed oil supplementation significantly reduced MDA levels in serum and liver, and suppressed hepatic oxidative stress. The immunological effects of soybean oil and its effect in improving pro-inflammatory status and oxidative stress have been reported [43]. Muga and Chao [44] found that treatment with fish oil reduced lipid peroxidation level to equal or below control levels and suggested that it may prevent or treat lipid peroxidation. Pipingas et al. [45] studied the effect of fish oil on health in 160 healthy individuals and suggested that consumption of daily fish oil prevents oxidative stress. Osteocytes can modulate the formation and function of both osteoblasts and osteoclasts. Apoptotic osteocytes release some proinflammatory cytokines such as TNF-α to stimulate osteoclastogenesis. Exposure to GCs can lead to osteocyte apoptosis and then bone loss. Our data showed that the prednisolone group recorded high values of inflammatory marker TNF-α. The GC-induced apoptotic osteocytes could increase the formation of TNF-α [5]. According to our research findings, treatment with oils rich in PUFAs decreased the inflammation. Recent investigation has reported that intake of PUFAs effectively decreased the inflammatory markers [27]. Muga and Chao [44] reported that treatment of fish oil regulated inflammation by decreasing the TNF-α concentration.
Our histological investigation of bones revealed that prednisolone affected bones at the cellular level and induced many abnormalities in the cortex and osteocytes, while treatment with oils, especially fish oil, reduced most of these side effects of prednisolone. According to Kasem et al. [46] histological examination of rat femur after injection of prednisolone to induce osteoporosis showed significantly decreased cortical thickness and osteocyte number of the osteoporosis group when compared with the normal control group. Derakhshanian et al. [47] recorded a reduction in cortical and trabecular thickness, as well as a significant decrease in the number osteoblasts in GC-treated rats. At the cellular level, GCs can increase apoptosis, inhibit mitosis, and suppress type II collagen synthetic ability in osteoblasts. Moreover, GCs inhibit cartilage growth, suppress synthesis of proteoglycans and decrease osteocyte feasibility, thereby decreasing bone strength [5]. PUFAs suppress the action of osteoclasts and enhance the activity of osteoblasts, thereby inhibiting bone resorption and promoting bone formation. Supplementation of fish oil improved BMD and the mechanical and histological characteristics of bones [48].
Summary
The treatment of osteoporosis remains a considerable problem. Therefore, preventing bone loss is an important demand nowadays. The present data indicated that feeding dietary oils containing omega-3 PUFAs such as fish oil, soybean oil or flaxseed oil could reduce the potential effect of prednisolone in inducing the development of osteoporosis in rats. This may occur through some intracellular pathways, including the improvement of calcium absorption, regulation of bone metabolism, differentiation of the osteoblast and osteoclast, suppression of oxidative stress and modulation of inflammatory response. Thus, these oils and fish oil, in particular, could be used as a natural approach to help prevent the bone loss associated with GC therapy, and this potential offers an interesting prospect for future research.




 XML Download
XML Download