

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Cardiovascular disease (CVD) is the leading cause of mortality worldwide, accounting for ~31% of global deaths in 2012 according to World Health Organization [1]. Atherosclerosis, a condition characterized by the accumulation of lipid-laden plaques in the blood vessel, is one of the major contributors to the pathogenesis of CVD [2]. High concentrations of serum total cholesterol (TC) and low-density lipoprotein cholesterol (LDL-C) facilitate atherosclerosis development [3]. The consumption of foods containing high amount of cholesterol, such as eggs and animal products, was limited to maintain normal serum TC and LDL-C concentrations until recently. National Dietary Guidelines of 2015–2020 eliminated the recommendation that limits daily dietary cholesterol consumption to 300 mg due to a lack of evidence supporting a positive relationship between dietary cholesterol intake and hypercholesterolemia in humans [4567].
While studies have shown a positive association of egg intake with CVD risk in humans with type 2 diabetes mellitus or coronary heart disease [8910], egg consumption does not raise plasma TC concentrations in healthy humans [1112] although the underlying mechanisms remain elusive. Studies have suggested that constituents of eggs, such as phospholipids, ovomucin, and caseins, may prevent an increase in plasma TC concentrations after consuming eggs [131415]. One compelling mechanism is related to high phospholipid (PL) contents in eggs. Eggs are rich in PL, accounting for approximately 10–40% of daily PL consumption in the U.S. [2]. Interestingly, it has been shown that PL can reduce intestinal cholesterol absorption in animals and humans [16171819]. Therefore, it is presumable that egg PL (EPL) may inhibit intestinal absorption of the cholesterol present in eggs. In support of this possibility, duodenal infusion of egg phosphatidylcholine, a major PL species in eggs, inhibited cholesterol absorption in rats as demonstrated by lower level of absorbed cholesterol in the lymph compared to control rats [20]. However, there is a lack of in vivo evidence that demonstrates a possible inhibitory effect of EPL on intestinal cholesterol absorption. Therefore, in this study, we sought to determine whether acute EPL administration or chronic consumption of EPL could alter intestinal cholesterol absorption as well as cholesterol metabolism in mice.
MATERIALS AND METHODS
Animal feeding and care
Acute administration of EPL
Male C57BL/6J mice (The Jackson Laboratory, Bar Harber, ME, USA) on a regular chow diet were gavaged with increasing daily dosages, i.e., 6, 11, and 19 mg/mouse (n = 5, 5, and 8, respectively), of EPL for 3 consecutive days. EPL (powdered L-α-phosphatidylcholine) was purchased from Avanti Polar Lipids (Alabaster, AL, USA). Fecal samples were collected 24 h after the last EPL gavage to determine intestinal cholesterol absorption as described below.
Chronic consumption of EPL
To investigate the effect of chronic consumption of EPL on cholesterol metabolism, male C57BL/6J mice at age of 11 weeks were fed a modified AIN-93M high fat/high cholesterol diet (HF/HC; 35% fat, 0.25% cholesterol, w/w) for 4 weeks to induce hypercholesterolemia. Subsequently, mice were fed a HF/HC control diet or a HF/HC supplemented with 0.4% or 0.8% EPL (n = 8 and 10, respectively) by weight (HF/HC-EPL) for 6 weeks. Detailed diet composition is described in Table 1. Mouse body weights and food consumption were recorded weekly. Intestinal cholesterol absorption was determined 5 weeks after mice were fed the experimental diets as described below. After 6 weeks on the experimental diets, mice were anesthetized with ketamine-HCl (110 mg/kg)/xylazine (10 mg/kg) (Henry Schein Animal Health, Dublin, OH, USA) after 8 h fasting, and then euthanized by cardiac puncture and subsequent cervical dislocation. Mouse blood samples were incubated at room temperature for 30 min, and then centrifuged at 1,500 × g at 4℃ for 15 min to remove red blood cells. Liver and small intestine samples were collected, weighed, snap-frozen, and stored at −80℃ until use.
EPL dosages were determined based on body surface normalization to a 70 kg human individual [21]. EPL dosages of 6, 11, and 19 mg/mouse for oral gavage are equivalent to 0.8, 1.5, and 2.5 eggs in humans. Dietary supplementations of EPL at 0.4% and 0.8% (w/w) are equivalent to daily consumption of ~1.5 eggs and ~3 eggs, respectively, in humans. All mice were maintained under the 12 h light/12 h dark cycle with free access to food and water. All procedures were approved by the Institutional Animal Care and Use Committee of the University of Connecticut (protocol # A13-026).
Intestinal cholesterol absorption rate analysis
Acute administration of EPL
For the evaluation of the acute effect of EPL on intestinal cholesterol absorption, mice were gavaged daily with 30 µL of vegetable oil only or of one of 3 EPL doses (6, 11, or 19 mg per mouse) for 3 days as previously described [22]. To determine intestinal cholesterol absorption rate, we used 14C-labeled cholesterol with 3H-labeled sitosterol which serves as a nonabsorbable control. On the first day, mice were administered with 0.1 µCi of [14C]cholesterol and 0.14 µCi of [3H]sitosterol (American Radiolabeled Chemicals, St. Louis, MO, USA) 30 min after EPL gavage. Fecal samples were collected 24 h after the last day of EPL gavage. Fecal lipids were extracted using Folch method as previously described [2324], and the ratio of [14C]cholesterol to [3H]sitosterol in fecal lipids were calculated to determine intestinal cholesterol absorption rates.
Chronic consumption of EPL
For the chronic study, mice fed with either a HF/HC control or HF/HC-EPL were gavaged with 0.09 µCi of [14C]cholesterol and 0.13 µCi of [3H]sitosterol for 2 consecutive days after 5 weeks of experimental diets and fecal samples were collected 5 days after the first gavage of [14C]cholesterol and [3H]sitosterol. Fractional intestinal cholesterol absorption was determined by the dual fecal isotope method used in acute study.
Blood chemistry
Circulating concentrations of TC and triglyceride (TG) were enzymatically measured using L-Type TG M kit (Wako, Richmond, VA, USA) and Cholesterol reagent set (Pointe Scientific, Canton, MI, USA), respectively, as we previously described [25]. Plasma concentrations of high-density lipoprotein cholesterol (HDL-C) were measured using the Cholesterol reagent set after precipitating apoB containing lipoproteins using HDL cholesterol precipitating reagent set (Pointe Scientific, Canton, MI, USA).
Gene expression analysis using reverse transcription quantitative realtime PCR (RT-qPCR)
Total RNA from liver and intestine was extracted as previously described [25]. The expression of genes involved in cholesterol transport, synthesis, and uptake was determined by RT-qPCR as we previously described using the SYBR Green procedure [25]. Glyceraldehyde 3-phosphate dehydrogenase (Gapdh) was used as a reference gene for data normalization.
RESULTS
Body and liver weights, and blood lipids in HF/HC diet-fed mice
Supplementation of 0.4% or 0.8% EPL diet for 6 weeks did not significantly alter body and liver weights of mice compared with HF/HC control mice (data not shown). EPL consumption did not alter circulating TC, HDL-C and TG concentrations compared to those of the control (Table 2).
Effect of acute and chronic consumption of EPL on intestinal cholesterol absorption
There were no significant differences in intestinal cholesterol absorption when mice acutely consumed EPL at dosages of 6 or 11 mg compared to control (Table 3). However, 19 mg of EPL administration significantly decreased intestinal cholesterol absorption compared to control (P = 0.01). Consumption of 0.4% EPL for 6 weeks, however, did not significantly alter intestinal cholesterol absorption in hypercholesterolemic mice.
Effect of chronic EPL consumption on intestinal and hepatic gene expression
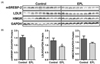
We determined whether chronic EPL consumption in a hypercholesterolemic condition could alter the expression of genes involved in cholesterol transport, synthesis, and uptake in the liver. mRNA levels of nuclear receptor subfamily 1 group H member 3 (Nr1h3), which functions as an oxysterol sensor, as well as its target gene, ATP-binding cassette transporter A1 (Abca1), were significantly decreased in 0.4% EPL-fed mice compared to the control (Table 4). Also, hepatic mRNA abundance of sterol regulatory element-binding factor 2 (Srebf2) was decreased by 0.4% EPL supplementation, while its downstream genes, such as HMG-CoA reductase (Hmgcr) and LDL receptor (Ldlr), were not significantly different between control and 0.4% EPL-fed mice. EPL supplementation at a dose of 0.4% did not significantly change mRNA of ATP-binding cassette transporter G5 (Abcg5), sterol regulatory element-binding factor 1c, and scavenger receptor class B type 1 (Scarb1) (data not shown). Mice fed 0.8% EPL supplementation did not show any significant changes in their hepatic mRNA levels of Nr1h3, Abca1, Srebf-2, Hmgcr, and Ldlr compared with control. However, hepatic SREBP-2 and LDLR proteins were decreased in 0.8% EPL group compared with control although their respective mRNA levels were not different (Fig. 1). Hepatic HMGCR protein levels were not altered by 0.8% EPL consumption.
In the small intestine, mRNA abundance of Niemann-Pick C1-Like 1 (Npc1l1), a major cholesterol transporter, was significantly decreased by 0.8%, but not 0.4% EPL consumption. However, any of EPL doses did not significantly alter mRNA levels of Hmgcr, Ldlr, Abca1, and Abcg5.
DISCUSSION
High plasma TC and LDL-C concentrations are the primary risk factors of CVD [26]. Therefore, dietary cholesterol was historically viewed as a dietary component that increased CVD risk [27], and the American Heart Association set a dietary recommendation to limit dietary cholesterol consumption to less than 300 mg/day in 1968 [28]. In the 2015 dietary guideline, the recommendation was eliminated based on mounting evidence that dietary cholesterol does not increase incidents of heart diseases [5272930]. However, the underlying mechanisms for the observation have not been clearly elucidated. Inhibition of intestinal cholesterol absorption largely affects plasma TC concentrations [31], and dietary PL from various sources, including egg yolk, soybeans, and milk, have been suggested to inhibit intestinal cholesterol absorption [1620]. Therefore, we sought to determine whether acute or chronic consumption of EPL, accounting for ~28% of total lipids in eggs by weight [2], may inhibit intestinal cholesterol absorption. In the present study, we found that acute EPL administration at a dosage equivalent to 2.5 eggs in humans significantly reduced intestinal cholesterol absorption in mice while chronic consumption of EPL equivalent to 1.5 eggs in humans was not able to alter intestinal cholesterol absorption in hypercholesterolemic mice. This inhibitory action of EPL on intestinal cholesterol absorption explains, at least in part, the observations in several studies which show that egg consumption did not increase plasma TC concentrations in healthy humans despite high daily intake of dietary cholesterol from eggs [1112].
Our study found that acute oral administration of 19 mg EPL significantly reduced intestinal cholesterol absorption in mice. This finding provides in vivo evidence, which supports the previous report that duodenal infusion of lipid emulsion containing egg phosphatidylcholine, a major PL species in eggs, reduced cholesterol levels in the collected lymph compared to control rats [20]. Although how EPL inhibits intestinal cholesterol absorption has not been clearly demonstrated, the inhibitory action of EPL may be related to its role in the modulation of interaction between micelles and enzymes/receptors involved in cholesterol absorption. We assume that it is possible that increased micellar EPL may inhibit binding of micelles to SR-B1, a cholesterol membrane transporter in the small intestine [32], thereby decreasing intestinal cholesterol absorption. In fact, Goncalves et al. [33] demonstrated that the composition of micelles affects an interaction between micelles and SR-B1. Using mixed micelles prepared with different concentrations of phosphatidylcholine, they reported a linear decrease in the binding capacity of micelles to SR-B1 with increasing concentrations of phosphatidylcholine. However, considering NPC1L1, but not SR-B1, is the major cholesterol transporter in the intestine [34], and cell-based as well as in vivo evidence is lacking to support the action of PL, it is not clear how much this action of PL in lowering the binding of micelles to SR-B1 can contribute to overall intestinal cholesterol absorption. Further studies are necessary to determine the role of EPL on the composition of micelles and their interaction with intestinal cholesterol transporters, i.e., NPC1L1 and SR-B1.
Another possible mechanism for the inhibitory action of EPL on cholesterol absorption is that EPL may impede the hydrolysis of micellar PLs by phospholipase A2 (PLA2), thus inhibiting the uptake of micellar cholesterol by the enterocyte. Studies have demonstrated that PLA2 is crucial for cholesterol uptake into the enterocyte by decreasing the micellar PL contents. Studies have shown that FPL 67047XX, a PLA2 inhibitor, decreased cholesterol absorption in vitro and in vivo. In Caco-2 cells, the PLA2 inhibitor abolished a PLA2-dependent increase in cellular cholesterol uptake from micelles [35]. Similarly, duodenum infusion of lipid emulsion containing the PLA2 inhibitor significantly reduced cholesterol absorption in rats [36]. Therefore, it is presumable that increased micellar PL contents due to EPL consumption may inhibit intestinal cholesterol absorption via impedance of PLA2 action. Further study is needed to determine whether the enrichment of micelles with EPL elicits inhibitory action against the binding capacity of micelles to SR-B1, and PLA2-dependent cellular uptake of cholesterol to inhibit intestinal cholesterol absorption.
In our study, although acute administration of high EPL dose significantly inhibited intestinal cholesterol absorption in mice, EPL consumption for 6 weeks did not alter intestinal cholesterol absorption in hypercholesterolemic mice. In addition, EPL supplementation was not able to counteract HF/HC diet-induced increases in plasma TC, HDL-C, and TG concentrations. In the small intestine, 0.8% EPL supplementation reduced Npc1l1 mRNA expression in hypercholesterolemic mice, however, the expression of other genes sensitive to cellular cholesterol levels, such as Hmgcr, Ldlr, Abca1, and Abcg5, in the small intestine were not altered by EPL. In the liver, mRNA abundance of Nr1h3, Abca1, and Srebf2 decreased by 0.4% EPL supplementation, and SREBP-2 and LDLR protein levels were decreased in 0.8% EPL-fed mice. However, the changes in cholesterol metabolism-related mRNA and protein levels did not alter plasma cholesterol levels in hypercholesterolemia-induced mice, suggesting that the inhibitory role of EPL against intestinal cholesterol absorption in mice fed a normal chow was not achievable in hypercholesterolemia-induced mice. The discrepancy of the role of EPL in the regulation of intestinal cholesterol absorption between acute and chronic studies might be due to cholesterol and fat contents used in the experimental diet. High contents of cholesterol and fat in the experimental diet used for the chronic study might not be counteracted by the levels of EPL supplementation. This result suggests that EPL does not counteract hypercholesterolemia in diet-induced obesity, which is consistent with the evidence that reports a positive correlation with the egg consumption and CVD risk in type 2 diabetic patients [89]. However, it would be worthwhile to conduct a study using different levels of cholesterol and fats in experimental diets to investigate the effect of chronic consumption of EPL on intestinal cholesterol absorption.
In conclusion, this study demonstrates that acute oral administration of EPL inhibits intestinal cholesterol absorption in mice fed a normal chow, while chronic EPL consumption with a HF/HC diet did not alter plasma TC levels and intestinal cholesterol absorption in hypercholesterolemic mice. Several mechanisms are suggested to demonstrate the inhibitory action of EPL on cholesterol absorption in the intestine, however, more research is needed to elucidate the possible mechanisms by which EPL inhibits intestinal cholesterol absorption at the whole body and cell levels. Despite the beneficial effect of EPL in cholesterol absorption, concerns of EPL consumption remain, as EPL provides a substrate for trimethylamine N-Oxide (TMAO), a CVD marker [2]. Miller et al. [37] has demonstrated that egg consumption increases the postprandial level of plasma TMAO in human. Therefore, further studies are warranted to investigate the effect of chronic EPL consumption on TMAO formation and its contribution to CVD.
Nonetheless, our study provides in vivo evidence that EPL administration inhibits intestinal cholesterol absorption. Importantly, this observation demonstrates for the first time to our knowledge that consumption of PL in eggs prevents intestinal cholesterol absorption, which provides an explanation why the consumption of eggs rich in cholesterol does not exert hypercholesterolemic effects in most of the human population.




 XML Download
XML Download