

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The occurrence of oxidative stress is relevant to both free radicals and reactive oxygen species (ROS) [12]. Free radicals, such as superoxide (·O2-), hydroxyl radical (·OH), nitric oxide (NO), and the non-radical oxidizing agents like hydrogen peroxide (H2O2) can generate ROS. Actually, under homeostasis, ROS participates in a series of cellular functional reactions. However, in an excessively exposed situation, ROS becomes toxic and increases oxidative damage, especially being harmful for cells [345]. Cellular damage by oxidative stress disrupts the normal mechanisms of cellular signaling [67]. According to Gille and Joenje [8], disturbing the prooxidant-antioxidant balance is a way to induce oxidative stress. Generally, H2O2 itself is stable in culture media; however, it can generate toxic ·OH in the cell through the transition metal-catalyzed reaction. Although cellular catalase has been reported to decompose H2O2 to water and oxygen to prevent ·OH formation, the function of catalase might loss balance when treated with high doses of H2O2 and the catalase activity is inversely correlated with the cytotoxic effect of H2O2 [9]. The exposure of cells to H2O2 results in DNA damage and eventually causes cellular apoptosis or/and induces inflammatory responses [10]. These alterations of homeostasis cause cellular damage by H2O2-induced oxidative stress. In the central nervous system, glial cells play a crucial role in protecting neurons [1112]. Oxidative stress in glial cells is closely related to neurodegenerative dysfunctions such as Alzheimer's disease (AD) and Parkinson's disease [1314]. Therefore, research on the regulation of glial cell function and protection from oxidative stress has gained interest.
Oxidative stress occurs with inflammation and activates apoptosis pathways [1215]. Inflammatory cells release cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) mediators, which are associated with age-related degenerative diseases [1617]. Inflammatory cytokines activate mitogen-activated protein kinases (MAPKs) pathways, including c-Jun NH2-terminal kinase (JNK) and p38 MAPK, which are important for cellular proliferation and apoptosis. Moreover, the phosphorylated JNK and p38 are known to regulate apoptotic cell death [18].
The edible mushroom Cordyceps militaris (CM) is a kind of traditional medical herb widely used for hundred years in Asia. Several studies have shown that CM has exerted functions on immunomodulatory, neuroprotection, anti-bacterial and antitumor properties [192021]. A previous research indicated the protective effect of CM against oxidative damage under high glucose induction conditions [22]. According to Ng and Wang [23], the Cordyceps species were reported to contain several active compounds, including cordycepin, ergosterol, and polysaccharides. Of these, the antioxidant property of cordycepin isolated from CM (3′-deoxyadenosine, C10H13N5O3) has been studied recently [2425]. Ramesh et al. [24] have reported that cordycepin attenuates oxidative stress and exerts antioxidant capacity in vivo. Moreover, polysaccharides isolated from CM have also demonstrated to inhibit mitochondrial injury and improve immune responses by scavenging ROS [2627]. In addition, CM has been reported to inhibit ROS generation and protect against oxidative stress induced in human dermal fibroblast cells [28]. The present study aimed to investigate the protective effect of CM against H2O2-induced oxidative stress and explore the related molecular mechanisms against oxidative stress and apoptosis in C6 glial cells.
MATERIALS AND METHODS
Sample and experimental groups
The ethanol extract of CM was obtained from the National Institute of Horticultural and Herbal Science (RDA, Jeolabuk-do, Korea). CM was dissolved in dimethyl sulfoxide (DMSO) to prepare a stock solution and further diluted with Dulbecco's modified Eagle's medium (DMEM) before the experiment. The experimental groups of C6 glial cells were categorized as follows:
(1) Normal: no treatment with CM or H2O2;
(2) Control: treatment with 300 µM H2O2;
(3) 0.5 µg/mL: treatment with CM (0.5 µg/mL) + H2O2 (300 µM);
(4) 1 µg/mL: treatment with CM (1 µg/mL) + H2O2 (300 µM);
(5) 1.25 µg/mL: treatment with CM (1.25 µg/mL) + H2O2 (300 µM);
(6) 2.5 µg/mL: treatment with CM (2.5 µg/mL) + H2O2 (300 µM).
Reagents
Griess reagent, 1,1-Diphenyl-2-picrylhydrazyl (DPPH), 2-deoxyribose, 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), dichlorofluorescin diacetate (DCF-DA), and DMSO were purchased from Sigma Aldrich Co. (St. Louis, MO, USA). Thiobarbituric acid (TBA) and trichloroactetic acid (TCA) were obtained from Acros Organics (Kangnam-gu, Seoul, Korea) and Kanto Chemical Co. (Tokyo, Japan), respectively. H2O2 and sodium pentacyanonitrosylferrate (III) dihydrate (SNP) were purchased from Junsei Chemical Co. (Tokyo, Japan). DMEM, fetal bovine serum (FBS), and penicillin-streptomycin were acquired from Welgene (Daegu, Korea). Radioimmunoprecipitation assay (RIPA) buffer and the protein marker were obtained from Elpis Biotech (Daejeon, Korea). The polyvinylidene fluoride (PVDF) membrane was purchased from Millipore (Billerica, MA, USA). Primary and secondary antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA) and Santa Cruz Biotechnology (Santa Cruz, MA, USA), respectively.
DPPH assay
The DPPH radical scavenging assay was performed as described previously by Hatano et al. [29]. Sixty micromoles of the DPPH solution were added following treatment of the sample (100, 250, 500, 1,000 µg/mL). The absorbance was measured at 540 nm following incubation for 30 min at room temperature.
·OH assay
The scavenging activity of the ·OH radical was measured according to the method described by Chung et al. [30]. The CM extract was mixed with a solution of 10 mM FeSO4· H2O-EDTA, 10 mM 2-deoxyribose, and 10 mM H2O2. The sample mixture was incubated at 37℃ for 4 h and was added to a solution containing 1.0% TBA and 2.8% TCA. The solution was boiled for 20 min and cooled in ice bath. The absorbance was measured at 490 nm.
NO assay
The scavenging activity of the NO radical was measured according to the method described by Marcocci et al. [31]. The sample (100, 250, 500, 1,000 µg/mL) was mixed with 5 mM SNP solution and incubated for 150 min [31], after which it was mixed with Griess reagent in a ratio of 1:1 and incubated at room temperature for 30 min. The absorbance was measured at 540 nm.
Cell culture
C6 glial cells were obtained from the Korea Cell Line Bank (Seoul, Korea) and were cultured in T75-flasks using DMEM, 10% FBS, and 100 U/mL penicillin- streptomycin at 37℃ under an atmosphere of 5% CO2. For subculture, the cell culture medium was replaced daily by using 0.05% trypsin-EDTA.
Measurement of cell viability
Cell viability was determined using the MTT assay as described by Mosmann [32]. C6 glial cells were seeded in a 96-well plate at a density of 5 × 104 per well and incubated for 24 h. Different concentrations of CM (0.5, 1, 1.25, 2.5 µg/mL) was administered to the cells for 1 h after which H2O2 (300 µM) was added. Following overnight incubation, the cell culture media was replaced with MTT solution and the plate was incubated for 30 min. Finally, cell viability was determined by measuring absorbance at 540 nm.
Measurement of ROS production
DCF-DA solution was used as a fluorochrome in ROS generation according to the method from Ma et al. [33]. At a concentration of 5 × 104 cells/mL, C6 glial cells were seeded into a 96-well plate and incubated for 24 h. The cells were treated with CM (0.5, 1, 1.25, 2.5 µg/mL) for 1 h, following which 300 µM H2O2 was added. Finally, DCF-DA (80 µM) was added to the cells followed by incubation for 30 min. ROS production in cells was monitored by FLUO star OPTIMA (BMG labtech, Ortenberg, Germany) at excitation and emission wavelengths of 480 nm and 535 nm, respectively.
Western blotting
The cells were washed with phosphate-buffered saline (PBS) after aspiration of the culture medium. Thereafter, the cells were lysed in lysis buffer containing a mixture of RIPA buffer and 1× protease inhibitor cocktail. For western blotting, an equal amount of proteins was loaded in each lane. Fifteen micrograms of total protein were separated using a 10% or 13% SDS-polyacrylamide gel and were transferred to PVDF membranes. The membranes were blocked with 5% non-fat milk and incubated overnight at 4℃ with primary antibodies [COX-2 and iNOS (1:200, Calbiochem); p-p38 (1:1,000, Cell Signaling); p-JNK (1:200, Santa Cruz); p-ERK (1:1,000, Cell Signaling); and β-actin (1:1,000, Cell Signaling)]. Finally, the membranes were probed with HRP-linked secondary antibodies. Protein expression was measured using the enhanced chemiluminescence detection system (Davinci Chemi, Seoul, Korea).
RESULTS
DPPH radical scavenging activity
DPPH is widely used to measure the ability of various biological substances to scavenge free radicals [34]. As shown in Table 1, the treatment with CM exact of the concentrations from 100 to 1,000 µg/mL showed a significant scavenging effect of DPPH radical. Moreover, the capacity on DPPH radical scavenging increased concentration-dependently.
NO radical scavenging activity
The NO radical is supposed to be related to the formation of peroxynitrite anion with high toxicity and oxidative damage [36]. Table 1 illustrated the NO radical scavenging activity of CM extract. It indicated that CM extract significantly increased NO scavenging ability in a concentration-dependent manner.
Effect of CM on cell viability in H2O2-induced C6 glial cells
We performed the MTT cytotoxicity assay to determine the effect of CM on cell viability (Fig. 1). We assessed the toxicity of various concentrations (0.1, 0.25, 0.5, 1, 1.5, 2.5, and 5 µg/mL) of the CM extract in C6 glial cells. At 5 µg/mL, the cell viability decreased to below 80%, indicating that CM might produce a toxic effect on glial cells at high doses (Fig. 2). When C6 glial cells were treated with 300 µM of H2O2, the cell viability was decreased to 74.13%, whereas the normal group was observed to be 100%. However, treatment of H2O2-induced cells with CM showed a significant increase in cell viability to 83.84% and 79.09% at the concentration of 1.25 and 2.5 µg/mL, respectively.
Effect of CM on ROS production in H2O2-induced C6 glial cells
The ability of CM to inhibit ROS production over a time course of 1 h was determined (Fig. 3). ROS levels in the H2O2-induced control group increased significantly than that in the normal group. Conversely, CM-treated groups significantly inhibited ROS production.
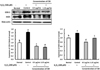
Effect of CM on COX-2 and iNOS protein expression in H2O2-induced C6 glial cells
The protein expression of COX-2 and iNOS was up-regulated in the H2O2-induced control group relative to that in the normal group (Fig. 4). However, the CM-treated groups significantly inhibited COX-2 and iNOS protein expressions in the C6 glial cells. These results indicate that CM regulates the inflammatory pathway in H2O2-induced C6 glial cells by down-regulating of COX-2 and iNOS protein expressions.
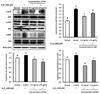
Effect of CM on p-p38, p-JNK, and p-ERK protein expression in H2O2-induced C6 glial cells
The inhibitory effect of CM on p-p38, p-JNK, and p-ERK protein expression in H2O2-induced C6 glial cells are shown in Fig. 5. The protein levels of p-p38 and p-JNK were increased when C6 cells were treated with H2O2 compared with the normal group. Moreover, compared with the normal group, p-ERK protein expression was up-regulated in the H2O2-treated control group, while the groups treated with CM significantly decreased the level of p-ERK, indicating that under H2O2-induced oxidative stress, CM might inhibit the p-ERK expression. Furthermore, p-p38, p-JNK, and p-ERK protein expression was significantly low in the 1.25 µg/mL CM-treated group. These results suggest that CM might inhibit p-p38, p-JNK, and p-ERK protein expressions under H2O2-induced oxidative stress.
DISCUSSION
Free radicals are involved in neural disorders that closely related to oxidative damage [3738]. In addition, researchers have demonstrated the correlation between oxidative stress and ROS generation, which synergistically cause degenerative diseases by inflammation and cell death [39]. Synthetic drugs are widely used as a treatment regimen for oxidative stress-related degenerative diseases, but it can cause irreversible side effects. Therefore, the use of natural medicines becomes hot topic of discussion. Dong et al. [40] has demonstrated antioxidant and cytotoxic properties of the methanol extract of CM against tumor cells. In addition, CM exhibits a protective effect in human dermal fibroblasts, suggesting the promising potential of CM treatment for degenerative diseases [28]. However, the neuroprotective effect of CM against oxidative stress has not been studied yet. The objective of this study was to investigate the neuroprotective effect of CM on oxidative stress-induced C6 glial cells.
Glial cells are closely related to the progression of degenerative diseases in the nervous system, including AD [4142]. Choi et al. [43] has reported on the C6 glial cell as a well-established cellular model for studying ROS generation and apoptotic pathways in neurodegenerative diseases. Impairment of neurons and glial cells by free radicals causes inflammation, which in turn promotes damage of the central nervous system [4445]. Furthermore, inflammatory cytokines released from glial cells and cellular apoptosis are important components of neurode-generative pathology [46]. A recent study by Yu et al. [47] showed that COX-2 and iNOS protein expression is up-regulated under oxidative stress by H2O2 in PC12 cells - a model for neurotoxicological study - indicating that H2O2 treatment induces neuroinflammation. Furthermore, rapid expression of COX-2 in glial cells may lead to neuroinflammation and affect brain function; there, COX-2 is gaining interest as a target in neurological diseases [48]. Additionally, iNOS has been shown to be involved in COX-2 regulation [49]. The present study demonstrates the role of CM in H2O2-induced inflammation in C6 glial cells by determining COX-2 and iNOS protein expression levels. The results indicate that treatment with the CM extract down-regulates COX-2 and iNOS protein expression in H2O2-induced C6 glial cells, whereas H2O2 up-regulates COX-2 and iNOS expression.
To investigate the protective effect of CM against oxidative stress and inflammatory in C6 glial cells, the expressions of JNK, p-38 and ERK were observed. Treatment of glial cells with H2O2 led to neuron death by up-regulating p38 and JNK protein expression. Xie et al. [50] have demonstrated evidence that the signaling pathways of the phosphorylated forms of JNK and p38 contribute to neuronal dysfunction and death. Kwon et al. [51] have also indicated the role of JNK and p38 signaling pathways in neurodegenerative diseases. Furthermore, ERK has been shown to participate in the regulation of cell growth and differentiation, response to cellular stress, and cytokines. Choi et al. [52] have demonstrated the inhibitory role of cordycepin on ERK phosphorylation in LPS-stimulated RAW 264.7 cells. In this study, we observed a significant up-regulation in the protein expression levels of p-JNK, p-p38, and p-ERK in H2O2-treated glial cells; however, treatment of H2O2-induced cells with CM showed a suppressive effect on the p-JNK and p-p38 signaling pathways, suggestive of the neuroprotective ability of CM.
The present study revealed that among the various concentrations of CM (0.5, 1, 1.25, 2.5 µg/mL), 1.25 µg/mL showed protective effect against oxidative stress in C6 glial cells. In our previous study, we investigated that oral administration of the CM extract (100 and 200 mg/kg/day) attenuates amyloid beta-induced cognitive impairment in mice models. We have shown that a dose of 200 mg/kg of the CM extract significantly improves learning and memory ability without demonstrating any side effects or hepatotoxicity (data not shown). According to Shin et al. [53], a dose of 200 mg/kg for animals could be translated into approximately 16 mg/kg for humans (60 kg) by using km factor (human: 37, mouse: 3). Therefore, the daily intake dose of CM extract for humans is approximately 960 mg [53].
In conclusion, this study demonstrates that H2O2-induced cell injury is related to ROS overproduction. However, treatment with CM significantly attenuates the damage in C6 glial cells by up-regulation of COX-2 and iNOS protein expression, and down-regulation of p-JNK and p-p38 protein expression. These results show the neuroprotective effects of CM against oxidative stress. It suggested that Cordyceps militaris could play the protective role against oxidative stress in glial cells.




 XML Download
XML Download