

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
The endoplasmic reticulum (ER) has multiple functions in the cell, with protein folding being the most important. An oxidative environment is maintained in the lumen of the ER, which is essential for disulfide bond formation and proper folding of proteins. Any disturbance that causes intracellular stress in the ER is termed ER stress. Multiple alterations in the cellular environment, including viral infections, elevated protein synthesis, failure of protein folding and degradation, and calcium influx across the ER membrane can result in ER stress. The response which allows the ER adapt to the changing environment and reinstate normal function is the unfolded protein response (UPR) [1]. The three different mechanisms of the UPR are a) attenuating translation to prevent synthesis of additional proteins, b) activating genes involved in ER protein folding and degradation, and c) the clearance of accumulated proteins from ER into the cytosol using the ubiquitin-proteasome system.
Three ER membrane-bound sensors, IRE1α (inositol-requiring protein-α), PERK (protein kinase RNA (PKR)-like ER kinase), and ATF6 (activating transcription factor 6) are responsible for the initiation of UPR signaling. The signaling mechanisms promote the reduction in protein synthesis, increase protein folding capacity, and activate ER-associated protein degradation [2]. When ER stress persists for a long period of time, the C/EBP homologous protein (CHOP) protein promotes the apoptotic pathway [3]. Studies have shown that ER stress is associated with various pathological conditions, including diabetes, cancer, neurodegeneration, and immune responses [4]. The ER proteins IRE1 and X box-binding protein 1 (XBP1) play important roles in the differentiation of B lymphocytes [5]. Overexpression of PERK has been reported to damage pancreatic β-cells, which can lead to the development of type 1 diabetes [6]. Also, protein overload in the ER results in the generation of reactive oxygen species and the resulting UPR impairs the function of not only the ER but also associated organelles, such as mitochondria. In hepatocytes, free fatty acids (FFAs) cause an imbalance in endoplasmic reticulum homeostasis, resulting in the ER stress associated with the development of hepatic steatosis [7]. In addition, chronic ER stress can activate inflammatory responses [8]. The three pathways of UPR; PERK, IRE1 and ATF6, have been shown to promote proinflammatory responses by activating transcription factors like nuclear factor kappa B (NF-κB) [9]. Therefore, maintaining of ER homeostasis is important for maintaining a healthy body.
Different kinds of fatty acids have different effects on ER stress. High concentrations of saturated fatty acids are known to be toxic to cells, while unsaturated fatty acids provide several positive effects. Palmitic acid is one of the most abundant FFAs present in the human diet and serum. Several in vitro experiments have shown that palmitic acid induces ER stress by upregulating genes related to UPR [10]. Palmitic acid is also known to stimulate the synthesis of ceramides and generation of reactive oxygen species, both of which can induce ER stress [11].
Emodin (1,3,8-trihydroxy-6-methylanthraquinone) is an anthraquinone derivative present in a variety of herbs. It is used as a traditional medicine in East Asian countries and is reported to have anti-inflammatory, antioxidative, and antitumor properties, as well as antidiabetic, neuroprotective, and hepatoprotective activities. Emodin is reported to have a protective effect against cell injury in AR42J cells by reducing calcium overload [12]. It also has a protective effect against severe acute pancreatitis [13]. In this study, the effect of emodin on palmitic acid-induced ER stress in HepG2 cells was evaluated.
Methods
Preparation of palmitic acid-BSA conjugate
Palmitic acid-BSA conjugate (PA) was prepared using sodium palmitate (Sigma Aldrich, St. Louis, MO, USA) and BSA (Fraction V-fatty acid free; MERCK, Billerica, MA, USA). Sodium palmitate was conjugated with BSA at a ratio of 6:1, following the protocol by Seahorse Bioscience. The final stock was filtered and stored at −20℃ until treatment in the cells.
Cell culture and MTT assay
HepG2 cells (ATCC-HB-8065; ATCC, Manassas, VA, USA) were cultured in DMEM containing penicillin-streptomycin (Hyclone, Logan, Utah, USA) with 10% FBS (Gibco, Invitrogen, New York, USA) at 37℃ in a 5% CO2 incubator. The cytotoxicity of PA and emodin (Sigma Aldrich) were analyzed individually using the MTT assay. The cells were seeded in 96 well plates and allowed to adhere. Cells were treated with different concentrations of PA (0 ~ 1,000 µM) and emodin (0 ~ 50 µM) separately. After 24 h of incubation cytotoxicity was analyzed using EZ-Cytox (DoGen Bio, Seoul, South Korea) solution. The concentrations of PA and emodin used for treatment were decided based on the MTT assay.
Treatment with palmitic acid
HepG2 cells were seeded in 6- well plates and treated with three concentrations of PA (250, 500, and 750 µM). After a 24 h incubation, the mRNA expression of ER stress related genes were evaluated to confirm the concentration of PA needed to induce ER stress. The concentration of PA used for further experiments was selected based on the results.
Treatment with emodin
HepG2 cells were treated with 750 µM of PA with or without emodin (1, 2 and 4 µM) and incubated for 24 h. The mRNA expression for various ER stress related genes and sirtuins were checked using PCR to examine the effect of emodin on PA- induced ER stress.
RNA extraction and RT-PCR
Total RNA was isolated from HepG2 cells using Trizol reagent (Invitrogen, Carlsbad, USA). Purity was determined using a Biodrop Duo instrument (Biochrom, Holliston, MA, USA) and cDNA synthesis was performed using cDNA reverse transcription kit (Applied Biosystems, Thermo Fisher, Waltham, MA, USA). This cDNA was then used to perform RT-PCR using a 7,500 Real-Time PCR system (Applied Biosystems, Foster City, CA, USA). The primers are given in Table 1.
Immunoblot analysis
The protein levels of phospho- IRE1α, IRE1α, phosphoeukaryotic initiation factor alpha (phospho-eIFα), eIFα, CHOP, and β-actin (Cell Signaling Technology, Beverly, MA) were determined using immunoblotting. For protein extracts, cells were homogenized in RIPA lysis buffer containing 1% protease inhibitor cocktail and 1% phosphatase inhibitor cocktail. After lysate clarification by centrifugation at 15,000 rpm (15 min, 4℃), the supernatants were collected and used for immunoblotting. Protein concentrations were measured using a BCA protein assay kit (Pierce Biotechnology, Rockford, IL, USA). The proteins were resolved by electrophoresis on a 10~12% sodium dodecyl sulphate-polyacrylamide gel and then transferred to an Immuno-Blot polyvinylidene difluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA) for immunoblotting.
Results
Effect of PA and emodin treatment on cell viability
There were no significant changes in cell viability at treatment concentrations of PA up to 750 µM 24 h after treatment (Fig. 1 (A)). However, the MTT assay showed that PA reduced cell viability 24 h after treatment with 1,000 µM, compared to the non-treated cells. Other studies have also reported that PA reduced cell viability compared to non-treated cells at a concentration of 1,000 µM [14]. Therefore, ER stress was induced by using working concentrations of PA from 250 to 750 µM.
The cells treated with 5 µM and 10 µM emodin showed reduced cell viability of about 8% and 18%, respectively (Fig. 1 (B)). High concentrations of emodin (> 10 µM) have been reported to reduce cell viability of liver cells [1516]. In our study, 5 µM of emodin showed an 8% reduction in cell viability, compared to non-treated cells. Therefore, concentrations less than 5 µM were selected for further experiments.
Induction of ER stress using different concentrations of PA
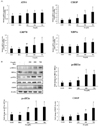
Chronic exposure of liver cells to high concentrations of fatty acids, particularly saturated fatty acids, leads to ER stress-mediated UPR activation. The changes in ER stress-mediated UPR associated genes were examined to determine the PA concentration needed for effective induction of ER stress. Consistent with previous reports, the mRNA expression levels of ER stress-related genes increased following treatment with PA at the concentration of 750 µM, compared to concentrations of 250 and 500 µM (Fig. 2 (A)). The mRNA expression of CHOP and spliced XBP1 (XBP1s) was increased significantly at the PA concentrations of 500 µM and 750 µM, compared to non-treated cells. Activating transcriptional factor 4 (ATF4) levels increased following treatment of PA at all concentrations, while 78-kDa glucose-regulated protein (GRP78) levels showed a dose- dependent increase.
Furthermore, the protein expression levels of ER stress-mediated UPRs were augmented by treatment with PA (Fig. 2 (B)). The levels of p-IRE1α and p-elF2α increased significantly after treatment with PA (500 and 750 µM), while their total protein levels remained constant. In addition, the levels of CHOP protein increased in HepG2 cells treated with PA (750 µM), compared with BSA alone. These results demonstrate the induction of ER stress in PA-treated HepG2 cells, so the PA concentration of 750 µM was selected as the working concentration for determining the effect of emodin on ER stress.
Effect of emodin on PA-induced ER stress
As shown in Fig. 3 (A), emodin treatment significantly reduced the expression of XBP1s and GRP78 irrespective of the dose. The mRNA expression of ATF4 decreased in a dose- dependent manner. However, there were no significant differences in the expression of CHOP mRNA expression between the emodin treated and non-treated cells.
These characteristics were also observed in the protein expression levels of ER stress-mediated UPRs (Fig. 3 (B)). The increase in levels of phosphorylated IRE1α and elF2α, and CHOP after treatment of HepG2 cells with PA were significantly decreased following emodin treatment (4 µM).
Effect of emodin on mRNA expression of sirtuins
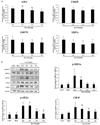
Previous studies have shown that increased expression of sirtuins attenuate ER stress in different cell lines. SIRT1 is known to protect cardiomyocytes from ER stress-induced apoptosis and SIRT3 is known to exert a protective effect against palmitate-induced apoptosis and dysfunction in pancreatic β-cells [1718]. Therefore, we evaluated the mRNA expression of all sirtuins in cells treated with PA alone and PA along with emodin. As shown in Fig. 4, the mRNA expression of SIRT1, SIRT2, SIRT3, SIRT5, and SIRT6, did not change significantly. However, the mRNA expression of SIRT4 and SIRT7 increased in the emodin and PA-treated HepG2 cells, compared to PA only treatment group.
Discussion
Prolonged ER stress can result in cell death. ER stress is one of the causes of neurodegenerative diseases, such as Alzheimer's and Parkinson's disease, as well as other diseases including diabetes mellitus, atherosclerosis, ischemia, heart diseases, kidney disease, and liver disease [192021]. Thus, protecting cells through preventive or therapeutic methods targeting the attenuation of ER stress could be an effective approach to maintaining health [22]. In this study, the effect of emodin on palmitic acid-induced ER stress in HepG2 cells was analyzed. Based on our results of MTT assay and mRNA expression, the PA concentration of 750 µM was used to determine the effect of emodin on ER stress.
There are three major proteins that regulate the ER stress, IRE1α, PERK, and ATF6. IRE1α is activated when unfolded proteins bind directly to it. It then activates the production of XBP1, which binds to the promoters of several genes that are involved in UPR and ERAD (ER-assisted degradation). This promoter activation helps to restore the protein homeostasis and cell protection. IRE1α also regulates the expression of several other proteins related to cell homeostasis [23]. The results of our study show that emodin reduces mRNA expression of spliced XBP1, suggesting that emodin prevents PA-induced ER stress. Recently, Wu et al. reported that emodin decreased the expression of ER stress markers, such as BiP, and IRE1α in rats with induced acute pancreatitis [13]. Our study showed a similar effect of emodin on HepG2 cells.
PERK activation leads to increased phosphorylation of the transcription factor eIF2α, which reduces protein synthesis and thereby decreases the protein load in the ER. However, ATF4 mRNA is one of the preferentially translated mRNAs when eIF2α is phosphorylated. ATF4 acts as an activator for CHOP and DNA damage inducible 34 (GADD34). CHOP triggers apoptosis related genes and GADD34 activates genes that dephosphorylate eIF2α under chronic ER stress conditions [24]. In this study, treatment with emodin reduced mRNA expression of ATF4, indicating that emodin may suppress PA-induced ER stress by downregulating ATF4. The expression of CHOP mRNA decreased, but not in a statistically significant way. Interestingly, the analysis of proteins using immunoblotting revealed that emodin significantly reduced the levels of CHOP. Therefore, we can conclude that emodin modifies CHOP at the protein level to exert its protective effect against ER stress.
ATF6 acts as a transcription factor that regulates genes related to ERAD and protein folding in the ER. ATF6 activates ER stress response genes, such as CHOP and BiP can also control the transcription of XBP1 [24]. In this study, the expression of GRP78/BiP was significantly reduced in the emodin treatment group compared to the PA alone treatment group.
Sirtuins are deacetylases that require NAD+ for their catalytic activity and was first discovered in yeast as Sir2. In mammals, sirtuins are linked to various cellular and metabolic processes. Seven sirtuins have been identified so far in mammals [25]. Since sirtuins specifically depend on NAD+ for their deacetylase activity, they are thought to play significant roles in responding to cellular stress and changes in cell homeostasis [26]. Several previous studies have shown that ER stress is directly linked to sirtuins, the most studied being SIRT1. SIRT1 exerts a protective effect on ER stress-induced cardiac injury by deacetylating eIF2α [17]. Furthermore, exendin-4, a GLP-1 receptor agonist, attenuates ER stress through a SIRT1 dependent mechanism [27]. In the light of previous studies showing that emodin exhibits an antifibrotic effect and antiangiogenic effect in cancer cells by enhancing SIRT1 expression, we evaluated whether the protective effect of emodin against ER stress is through sirtuin activation [2829]. Therefore, we examined the mRNA expression of all sirtuins (1 to 7) in the cells. Surprisingly, the mRNA expression of SIRT1 did not change in the emodin treated cells compared to the palmitic acid treated cells. Similar results were observed for the mRNA expression of SIRT2, SIRT3, SIRT5 and SIRT6. Interestingly, mRNA levels of SIRT4 and SIRT7 increased following emodin treatment. One previous study showed that overexpression of SIRT7 reduces phosphorylation of eIF2α and target gene expression, thereby reducing ER stress in a tunicamycin-induced ER stress animal model. The results of our study were similar. The liver tissue had the highest level of SIRT7 amongst the tissues of the body. SIRT7 acts as a cofactor for the transcription factor Myc to suppress protein expression in ribosomes by interacting with promoters of ribosomal protein genes and thus attenuates ER stress [30]. While no other studies have demonstrated the effect of SIRT4 on ER stress, our study showed that emodin may attenuate ER stress by the action of SIRT7 and/or SIRT4 on the ER stress gene transcription factors.
In the present study we did not demonstrate the induction of SIRT4 and/or SIRT7 protein by emodin. Therefore, further studies such as modulating the expression or activity of SIRT4 and SIRT7 are needed to elucidate a precise mechanism of emodin against ER stress. In conclusion, our results suggest that emodin attenuates ER stress in HepG2 cells. Emodin reduces the expression of ER transcription factors and their target genes. Since ER stress is associated with many metabolic diseases, such as non-alcoholic fatty liver, emodin could be used as an agent to attenuate the ER stress and reduce cellular damage.
Summary
The possible protective effect of emodin against PA induced-ER stress was evaluated in this study. 750 µM of PA was effective in inducing ER stress, as evidenced by the increased expression of proteins, such as IRE1α, eIF2α and CHOP. Treatment of cells with emodin decreased the mRNA expression and protein expression of the genes that were upregulated by PA treatment, thereby showing that it has a protective effect against PA-induced ER stress. Emodin also increased the mRNA expression of SIRT4 and SIRT7 suggesting that SIRT4 and/or SIRT7 may regulate the expression of UPR-associated transcription factors to reduce ER stress. Therefore, emodin could be used as an agent to reduce ER stress and thereby ameliorate several metabolic conditions, such as non-alcoholic fatty liver and type 2 diabetes.
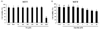


 XML Download
XML Download