

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Obesity is a major health concern and can lead to metabolic disorders including type 2 diabetes, non-alcoholic fatty liver disease (NAFLD), dyslipidemia, hypertension, coronary heart disease, and certain cancers, as well as psychological problems [123]. Obesity, a complex disease resulting from imbalanced energy intake and expenditure, is characterized by the accumulation of abnormal and/or excessive body fat, which causes harmful effects in humans [4]. There are a variety of underlying causes, which include genetic, social, and environmental factors, as well as diet [5]. Recommended therapeutic strategies in obese individuals include the restriction of simple carbohydrate and saturated fatty acid intake, increased physical activity, and administration of anti-obesity drugs [6]. Despite various attempts to reduce obesity, which have included the development of anti-obesity products, it remains a challenging problem. Synthetic drugs have drawbacks in that they are relatively costly, and that prolonged use may have negative long-term side effects [7]. Therefore, alternative therapies using naturally-occurring compounds have been proposed.
Brassica vegetables, which belong to the Cruciferous family and include cabbage, Brussels sprouts, cauliflower, and broccoli, among others, are consumed in different regions of Asia and central Africa [8]. Brassica juncea L. has been used as an Ayurvedic medicine for centuries [9]. Its leaves contain abundant vitamins, polyphenolics, sterols, triterpene alcohols, and other structurally diverse phytochemicals [1011]. They also contain diverse glucosinolates and isothiocyanate precursors, with the glucosinolate content being particularly high in Brassica juncea L. leaves [1213]. Glucosinolates are a unique group of sulfur-containing secondary plant metabolites, and their hydrolysis products (isothiocyanate, sulforaphane) have been shown to modulate lipid metabolism in vivo and in vitro [1415]. Thakur et al. [16] demonstrated that Brassica juncea L. leaf extract (BLE) contains kaemferol and isorhamnetin. Brassica juncea L. leaves display antioxidant and free radical scavenging activities [1117]. BLE has also been shown to protect against diabetes-associated oxidative stress [18]. Additionally, a previous study showed that BLE has potential antihyperglycemic and anticataractous effects against streptozotocin (STZ)-induced diabetic cataracts in rats [19], while another study showed that BLE has anticancer effects [20]. However, no studies have investigated the anti-obesity and hypolipidemic properties of BLE in high-fat, high-cholesterol (HFC) diet-fed rats. Thus, we investigated the effect of BLE supplementation on obesity in HFC-induced rats and examined the mechanism of its anti-obesity action.
MATERIALS AND METHODS
Preparation of Brassica juncea L. leaf extract (BLE)
Brassica juncea L. was purchased from a local market in Gwangju, Korea. Rinsed Brassica juncea L. leaves were freeze-dried for 72 h and extracted with 80% (v/v) ethanol at 70℃ for 3 h with constant stirring, following which the extract was filtered through Whatman filter paper. Ethanolic leaf extracts were concentrated using a rotary evaporator (Eyela Vacuum NVC-1100, Tokyo, Japan) and stored at −80℃ until analysis. The yield of BLE was 34.52% (w/w).
Animals and treatment
Five-week-old Sprague-Dawley (SD) rats were purchased from Orient Bio, Inc. (Daejeon, Korea). All animal care and experimental protocols were approved by the Animal Care and Use Committee of Chosun University, Korea (CIACUC2017-S0045). Rats were housed under standardized conditions for animal facilities: a 12 h light/dark cycle, 22 ± 2℃ room temperature, and 55 ± 5% humidity. All rats had free access to standard laboratory chow and water. The diet composition is shown in Table 1. After 7 days of adaptation, the animals were randomly divided into four groups (n = 8) and fed normal or experimental diets for 6 weeks as follows: (1) a normal diet group (ND); (2) high-fat, high-cholesterol diet group (HFC; 10% fat based on AIN-93 diet plus 1% cholesterol, w/w); (3) high-fat, high-cholesterol diet with 3% BLE diet group (HFC-A1); (4) high-fat, high-cholesterol diet with 5% BLE diet group (HFC-A2). Body weight and food intake were monitored once per week.
Sampling procedures
At the end of the experiment, fasted rats (12 h) were anesthetized and sacrificed. Blood samples were centrifuged at 1,150 × g for 20 min at 4℃ to collect serum. The liver and adipose tissue (mesenteric, epididymal, and retroperitoneal depots) were harvested, weighed immediately, and stored at −80℃ until analysis.
Biochemical analyses
Serum levels of triglyceride (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), alanine transaminase (ALT), aspartate transaminase (AST), alkaline phosphatase (ALP), and lactate dehydrogenase (LDH) were measured using a FUJI DRI-CHEM 3500 analyzer (Fujifilm, Tokyo, Japan). Low-density lipoprotein cholesterol (LDL-C) was calculated using the Friedewald formula [21], and the atherogenic index (AI) and cardiac risk factor (CRF) were calculated using the Rosenfeld formula [22].
RNA extraction and reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was isolated from frozen liver tissue using an RNeasy kit (Qiagen, Hilden, Germany). To synthesize cDNA, 1 µg of RNA was reverse-transcribed using Oligo-dT primers and SuperScript III Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA). The sequences of the primers were as follows: glucose 6-phosphate dehydrogenase (G6pdh): forward 5′-GTTTGGCAGCGGCAACTAA-3′, reverse 5′-GGCATCACCCTGGTACAACTC-3′; acetyl-coA carboxylase (Acc): forward 5′-CAACGCCTTCACACCACCTT-3′, reverse 5′-AGCCCATTACTTCATCAAAGATCCT-3′; fatty acid synthase (Fas): forward 5′-GGACTGAACGGCATTACTCG-3′, reverse 5′-CATGCCGTTATCAACTTGTCC-3′; cholesterol 7α-hydroxylase (Cyp7a1): forward 5′-GCCGTCCAAGAAATCAAGCAGT-3′, reverse 5′-TGTGGGCAGCGAGAACAAAGT-3′; low-density lipoprotein-receptor (Ldlr): forward 5′-ATTTGGAGGATGAGAAGCAG-3′, reverse 5′-CAGGGCGGGGAGGTGTGAGAA-3′; β-actin: forward 5′-GTGGGGCGCCCCAGGCACCAGGGC-3′, reverse 5′-CTCCTTAATGTCACGCACGATTTC-3′. The reaction conditions were as follows: for G6pdh, Acc, and β-actin: 30 cycles of 95℃ for 15 s, 60℃ for 60 s, and 72℃ for 30 s; for Cyp7a1 and Ldlr: 30 cycles of 94℃ for 3 min, 60℃ for 1 min, and 72℃ for 2 min; for Fas: 27 cycles of 94℃ for 30 s, 55℃ for 30 s, and 68℃ for 30 s. PCR amplification employed β-actin as an internal control. PCR products were separated on a 1.5% agarose gel and visualized by ethidium bromide staining and UV transillumination.
Western blot analysis
Liver protein was extracted with lysis solution containing 1 mM EDTA, 1% Triton X-100, 150 mM HEPES, protease inhibitors, and phosphatase inhibitors (GenDEPOT, Barker, TX, USA). Equal amounts of protein were separated by 8–12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes. The membranes were blocked with 3% bovine serum albumin and then incubated overnight with primary antibodies at 4℃. Antibodies to peroxisome proliferator-activated receptor α (PPARα) and β-actin were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). After incubation with primary antibodies, the membranes were incubated with the corresponding horseradish peroxidase-conjugated secondary antibodies (Santa Cruz) for 1 h at room temperature. The membranes were then treated with ECL detection reagent (Amersham Bioscience, Little Chalfont, UK). The relative band density was determined by densitometry with Image J software (National Institutes of Health, Bethesda, MD, USA).
Histological analyses
The liver tissue was fixed in 4% formalin, and liver sections were stained with Oil red O (Sigma-Aldrich, St. Louis, MO, USA) (100 × magnification). The areas of randomly selected adipocytes were measured using Image J software (National Institutes of Health).
Statistical analyses
All data are presented as means ± standard deviations (SD). Statistical analyses were performed using Prism GraphPad 6 software (GraphPad, Inc., La Jolla, CA, USA). All results were compared by one-way analysis of variance with Tukey post hoc analysis. Differences were considered significantly different at P < 0.05.
RESULTS
Effect of BLE diet on body composition
Table 2 shows the effect of BLE on the final body weight, body weight gain, food intake, and food efficiency ratio (FER) of experimental rats following 6 weeks of treatment. HFC administration caused a significant increase in the final body weight of rats when compared to that of rats in the ND group. The body weight gain was also significantly higher in the HFC group than in the ND group, but was significantly lower in the HFC-A2 groups than in the HFC group. Food intake was not significantly changed by BLE supplementation. The FER of rats fed the HFC was significantly higher than that of rats fed the ND, whereas HFC-fed rats supplemented with 5% BLE had significantly lower FERs than rats in the HFC group.
Effect of BLE diet on organ weight
To determine whether BLE-linked weight loss effects were due to a decrease in the fat mass, organ tissue weights were examined. As shown in Table 3, the amounts of liver, mesenteric and total adipose tissues in the HFC group were significantly higher than in the ND group. Additionally, 5% BLE supplementation in rats fed the HFC significantly decreased the masses of mesenteric, epididymal, and total adipose tissues by 30.3%, 27.2%, and 22.5%, respectively, compared to rats fed just the HFC. Liver weight did not significantly change in the BLE-treated HFC group. The mesenteric and total adipose tissue weights were significantly different between the HFC-A1 and HFC-A2 groups.
Effect of BLE diet on serum lipid parameters
We examined the effect of BLE supplementation on serum lipid profiles in the HFC group. Table 4 shows that the rats fed the HFC had significantly higher TG, TC, and LDL-C levels, and lower HDL-C levels, than rats from the ND group. However, the levels of TG, TC, and LDL-C in the HFC group decreased by 35.8%, 19.6%, and 32.5%, respectively, following HFC supplementation with 5% BLE. HDL-C levels were significantly higher (19.1%) in the group given the HFC-A2 diet compared to levels in the HFC group, but there were no significant differences between the HFC-A1 and HFC-A2 groups. Next, we measured the AI and CRF as predictors of cardiovascular risk [26]. AI and CRF of rats from the HFC group was significantly higher than those in the ND group. However, administration of 3% and 5% BLE resulted in significant lowering of AI and CRF relative to the HFC group.
Effect of BLE diet on hepatic function
Liver functions were measured by changes in hepatic markers (ALT, AST, ALP, and LDH) after BLE supplementation in HFC-fed rats for 6 weeks. As shown in Fig. 1, the activities of ALT, AST, ALP, and LDH in the HFC group were significantly higher than those in the ND group. The increased activities of ALT, AST, and LDH in the HFC group were significantly lowered following HFC-A1 and HFC-A2 administration. There was, however, no significant difference between BLE treatments with respect to serum ALT, ALP and LDH activities.
Effect of BLE diet on the lipid profile in tissues
The contents of hepatic, mesenteric, and fecal lipids are shown in Table 5. Rats fed HFC diets had significantly higher hepatic total lipid, TG, and TC levels than rats from the ND group. However, TG and TC levels decreased significantly when HFC diets were supplemented with 3% and 5% BLE. The levels of mesenteric total lipid, TG, and TC were significantly lower in HFC-A1 and HFC-A2 rats than in HFC-fed rats. The total lipid and mesenteric TC levels were significantly different between the HFC-A1 and HFC-A2 groups. Furthermore, the level of fecal TC in the HFC group was 3.9-fold higher than in the ND group. In rats treated with 5% BLE, the levels of TC in fecal lipids were significantly higher (by 20.7%) than levels in the HFC group. Fecal TG levels did not differ significantly between groups.
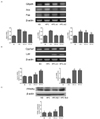
Effect of BLE diet on hepatic mRNA and protein expression
G6pdh, Acc and Fas are key enzymes in fatty acid (FA) synthesis. Fig. 2A shows that HFC induced significantly higher G6pdh, Acc, and Fas mRNA expression levels than were detected in the ND group. The mRNA levels of G6pdh and Acc were significantly lower in the HFC-A1 and HFC-A2 groups than in the HFC group. The mRNA expression of Fas in the HFC-A2 group was significantly lower than in the HFC group, but there was no difference between the HFC-A1 and HFC-A2 treated groups. Furthermore, we examined the effect of cholesterol synthesis in the HFC group after BLE supplementation. Our data showed that Cyp7a1 mRNA levels did not change significantly between the ND and HFC groups. After treatment of the HFC group with 3% and 5% BLE, mRNA levels of Cyp7a1 increased significantly (Fig. 2B). In addition, Ldlr mRNA expression was significantly higher in HFC-fed rats supplemented with 3% and 5% BLE than in untreated HFC-fed rats (Fig. 2B). We also observed higher levels of PPARα protein expression in the HFC-A2 group than in the HFC group (Fig. 2C).
Effects of BLE supplementation on liver histology
To examine the effect of BLE on liver histology, we performed histological analyses of HFC-fed rats. The liver of ND rats did not contain lipid droplets, while hepatic lipid droplet accumulation was observed in HFC-fed rats. Livers from HFC-fed rats supplemented with 3% and 5% BLE showed fewer lipid droplets than were observed in the HFC rat livers (Fig. 3).
DISCUSSION
Obesity impairs personal lifestyle and is the main contributing factor in several chronic diseases, including type 2 diabetes, cardiovascular disease, and cancer. While there are several types of obesity-treatment drugs, these may cause severe side effects, including mortality. Therefore, the identification of alternative phytonutrients which lack side effects is a worthwhile goal in the prevention and treatment of obesity. In the present study, we investigated the anti-obesity effects of dietary supplementation with 3% and 5% BLE on HFC-induced obesity in SD rats for 6 weeks. The administration dose of 3% and 5% BLE extract were determined based on a previous study [2728]. We found that, while food intake did not differ between the HFC-A1 and HFC-A2 groups, FER was significantly lowered in the HFC group after administration of 5% BLE.
High fat, high cholesterol diets increase the risk of dyslipidemia [29] and nonalcoholic steatohepatitis (NASH), which is associated with diabetes, hypertension, and hepatocarcinoma [30]. In this animal study, we established an HFC diet-induced obesity model which succeeded in increasing the body weight of rats and levels of TG, TC, and LDL-C, and decreasing the level of HDL-C. According to our results, dietary supplementation with BLE reversed the abnormal serum lipid profiles induced by dyslipidemia in HFC-fed rats, restoring healthy AI. We also found that fecal lipid contents were significantly higher in the HFC-A1 and HFC-A2 groups than in the HFC group. These results, along with the observed decreases in serum TG and TC levels, suggest that BLE is beneficial for improving hyperlipidemia and hypercholesterolemia caused by a HFC diet.
Dietary high fat can lead to hepatic fat accumulation and liver dysfunction [31], conditions indicated by leakage of cellular enzymes into the bloodstream. Serum ALT, AST, ALP, and LDH are biochemical markers for the functional diagnosis of liver damage or cardiovascular disease [32]. Our results showed that increased levels of ALT, AST, and LDH in the HFC group were significantly lowered through supplementation of HFC diets with 3% and 5% BLE, indicating that Brassica juncea L. leaf extracts have a protective effect against HFC-induced liver injury.
HFC-fed rats to which BLE was administered showed significantly decreased organ weights and lowered masses of mesenteric, epididymal, and total adipose tissues. Increased adipose tissue mass, particularly the amount of visceral fat, is associated with an increased risk of chronic diseases, and many factors secreted by preadipocytes are involved in the inflammatory response to infection [33]. The body and organ weight reductions suggest that supplementation with Brassica juncea L. can prevent fat accumulation and help treat obesity.
Accumulation of lipid droplets in hepatocytes causes hepatic steatosis, including an increase in peripheral lipolysis, upregulation of FA transporters, and an increase in de novo lipogenesis which can lead to inflammation, fibrosis, and hepatocellular carcinoma [3435]. To further investigate the regulation of hepatic lipid metabolism, we examined the mRNA levels of the lipogenic enzymes G6pdh, Acc, and Fas. Brassica juncea L. leaf extract-treated rats in the HFC group showed significant reductions in G6pdh, Acc, and Fas mRNA expression levels. Our results revealed an improvement in hepatic and serum lipid profiles after BLE treatment, which appeared to be related to the inhibition of G6pdh, Acc, and Fas gene expression. Additionally, we observed that PPARα protein expression was significantly higher in the HFC-A2 group than in the HFC group. PPARα is predominantly expressed in tissues such as the liver, heart, brown adipose tissue, and muscle, and regulates target genes involved in lipid metabolism and obesity [36]. PPARα increases fatty acid oxidation and decreases the expression of fatty acid synthesis target genes such as Acc and Fas, thus decreasing dyslipidemia-related metabolic syndrome in vivo and in vitro [37]. Stimulation of β-oxidation by PPARα activation regulates a mechanism for decreasing tissue lipid content, therefore preventing lipid accumulation [37]. It has been reported that obese patients with impaired fat oxidation failed to lose weight [38]. In line with this, PPARα-deficient mice showed abnormal TG and cholesterol metabolism and became obese [37]. Thus, our findings suggest that Brassica juncea L. leaves may be clinically useful for treating dyslipidemia. Further studies are needed to determine the mechanisms underlying these effects.
Next, we investigated the effect of BLE supplementation on the expression of genes responsible for cholesterol metabolism. We found that the mRNA level of Ldlr was higher in the HFC-A1 and HFC-A2 groups than in the HFC group. Ldlr on cell surfaces plays a major role in the uptake of circulating LDL-C into cells [39]. Increased Ldlr expression in the liver is negatively correlated with lower circulating LDL-C resulting from enhanced uptake and clearance of LDL-C [40]. BLE in the HFC group also showed significantly higher expression of Cyp7a1 mRNA than was observed in the HFC group. Cyp7a1 is the first and rate-limiting enzyme involved in the biosynthetic pathway that converts secreted cholesterol into bile acids [41]. Bile acid synthesis plays a major role in hepatic regulation of cholesterol homeostasis as well as in the catabolism of cholesterol [42]. Li et al. [43] demonstrated that transgenic overexpression of Cyp7a1 in the liver induced resistance to high-fat diet-induced obesity and decreased inflammation in mice. In addition, constitutive transgenic expression of Cyp7a1 in C57BL/6J mice prevented diet-induced hypercholesterolemia and blocked the development of atherosclerosis and gallstones [44]. Thus, our results suggest that Brassica juncea L. leaf promotes cholesterol uptake and conversion into bile acids in the liver, and increased cholesterol in the feces. In the present study, we observed that supplementation of a HFC diet with 3% and 5% BLE improved serum and organ lipid parameters and had a cholesterol-lowering effect in rats. Thus, BLE may be a useful natural reagent for treating obesity.
Several research questions were not addressed in our study. For example, the functional molecules responsible for the anti-obesity effects of BLE extracts were not identified. Additionally, in order to study the detailed molecular mechanisms underlying the described effects, a comprehensive in vitro study would need to be conducted. Further research would reveal the identities and mechanisms of action of the active molecules of BLE extracts responsible for the observed anti-obesity effects.







 XML Download
XML Download