

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Dementia is a complex and multifaceted disorder that manifests in the form of behavioral, cognitive and emotional impairments. As dementia becomes a more prevalent disorder posing significant burdens on public health, individual livelihood and economic security, health professionals must gain a better understanding of how to manage and even treat this condition.
The prevailing school of thought holds that memory loss associated with dementia is related to reduced cholinergic function induced by the development of beta amyloid plaques and tau protein tangles [1]; eventually accompanied by the damage and death of neurons [2]. The cholinergic hypothesis points to pathways in the cerebral cortex and basal forebrain that become compromised; subsequently leading to cognitive impairment [3]. Cognitive impairment may also be induced by heavy alcohol use - which has risen to epidemic proportions. The effects of alcohol on the brain include, brain tissue atrophy, reduction in number of cortical neurons in the superior frontal cortex, hypothalamus and cerebellum, and inhibition of N-methyl-D-aspartate (NMDA) [4]-linked to neurotoxicity, increased oxidative stress and the loss of cholinergic muscarinic receptors [5]. Once progressed, this condition leads to alcohol-related dementia.
There are no drugs that slow or stop the progression of Alzheimer's disease (AD), but six FDA-approved drugs are prescribed to alleviate symptoms [2] - donepezil being one of them. These muscarinic agonist drugs act through the inhibition of acetylcholinesterase (AChE), responsible for breaking down acetylcholine (ACh) in the neural synapse. However, the effectiveness of these drugs differs from patient to patient, and adverse effects have been reported including, gastrointestinal symptoms [6], night time disturbances [7] and cardiovascular complications [8]. Thus, the administration of a naturally occurring substance, which potentially has the same alleviative potency as a synthetic psychotropic drug, would be of great interest to the medical field.
Fermented Laminaria japonica (FL) is at the forefront of investigation into the utilization of sea tangle as health foods and even treatments for numerous disease states. Our laboratory previously demonstrated the effectiveness of γ-aminobutyric acid (GABA)-enriched FL in stimulating increased serum brain-derived neurotrophic factor (BDNF) [9] - implicated in the pathogenesis of dementia [10] and AD [1112]. Higher serum BDNF levels were also associated with lower risk for dementia and AD in a ten-year prospective cohort study in 60 + -year-olds [13]. Park et al. [14] investigated the cognitive improving efficacy of Saccharina japonica (also known as Laminaria japonica) in response to trimethyltin chloride-induced memory deficiency and reported recovered choline acetyltransferase, cAMP response element binding protein (CREB) and BDNF immunopositive neurons in the hippocampus. Considering the recent findings revealing potential neurogenic improvements as a result of FL supplementation, it is feasible to suggest a role for dietary intervention in providing a protective effect on the brain in neurodegenerative conditions.
In the laboratory setting, scientific research commonly utilizes scopolamine (an anticholinergic agent) to induce amnesia as a model for dementia. Similarly to alcohol, scopolamine causes oxidative stress in various brain regions and specifically acts as a nonselective muscarinic acetylcholine antagonist - related to a marked deficit in memory and cognitive abilities. Therefore, the purpose of the present study was to evaluate the effects of FL on scopolamine and alcohol-induced memory impairment, in comparison to current drug therapy intervention. The implementation of hippocampus-dependent memory assessment tests and analysis of brain tissue expression and cholinergic activity were utilized to examine the underlying mechanisms providing a protective effect on memory-impaired mice.
MATERIALS AND METHODS
Preparation of Laminaria japonica
FL was prepared using a modified version of a previous procedure [9]. The sea tangle was added to water at a ratio of 1:15 (w/v) with the addition of 3% yeast extract and 1% glucose, based on the amount of sea tangle added to the mixture. After autoclaving at 121℃ for 30 minutes (min), a sea tangle solution was obtained and the Lactobacillus brevis BJ20 (accession No. KCTC 11377BP) culture broth was added to the solution at a concentration of 5% (v/v) (Table 1), which was mixed and incubated at 37℃ for 2 days. During the preparation process, the GABA content had been controlled to a range between 40–60 mg within 1,000 mg of FL. Based on high performance liquid chromatography analysis, it was confirmed that the mean content of GABA was 54.5 ± 0.071 mg g−1 in the FL used in this study. A spray-drying method was used to prepare the FL to be powered prior to ingestion. Of the FL50 solution amino acid concentration, glutamic acid constituted 58.0 g/100g AA (amino acid) and aspartic acid 33.4 g/100g, and GABA was not detected before the fermentation process. However, GABA content was greatly increased (49.5 g/100g) during fermentation with Lactobacillus brevis BJ-20. Alanine was also increased after fermentation. 50 mg/kg was the chosen dosage based on evidence corroborating its effectiveness in improving cognitive function, compared to 100 and 200 mg/kg [14].
Animals
5-week old, male C57BL/6J mice weighing 25–28 g were purchased from Samtako BIOKOREA (Gyeonggi-do, Osan, Korea) and housed in a regulated environment (temperature, 22 ± 3℃; relative humidity, 50 ± 10%; 12-h light/dark cycle beginning at 07:00). All experiments were conducted in accordance with the guidelines of the Southeast Medi-Chem Institute (SEMI, Institutional Animal Care and Use committee) (ethical approval number: SEMI-16-05).
Experimental design and drug administration
Initially, the animals were divided into three main group categories: a control group (CON), scopolamine-induced dementia group (Sco) and ethanol-induced dementia group (EtOH) (Table 2). For the induction of dementia, one group received 5 mg/kg of scopolamine via intraperitoneal injection (Sco, Sco + Dpz, Sco + FL50), three days before dissection. The other experimental group orally consumed 2 g/kg of ethanol (conc % W/V) (Deoksan, Korea), daily, for four weeks. Treatment involved a positive control administered with 4 mg/kg Donepezil (Sco + Dpz and EtOH + Dpz) or 50 mg/kg FL50 extract (SEMID 1604-06) (Sco + FL50 and EtOH + FL50), daily, over the 4-week experimental period by oral gavage and orally consumed, respectively (Fig. 1). The CON group orally received a 0.9% saline solution, which was also included in the treatments of the aforementioned Sco and EtOH groups. All animals were fed on standard laboratory chow (Samtako BIOKOREA, Korea), with free access to water. A fasting period took place 24 hours (h) before dissection of the animals, when only water was allowed to be consumed. The experimental animals were treated at a specific time (10:00–12:00 am) in consideration of diurnal variation in enzyme activity.
Morris water maze test
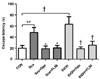
The Morris water maze (MWM) test design utilized in this study was modified from Morris [15], used to assess spatial learning ability and memory. A white circular tank (90 cm diameter and 40 cm high) with a featureless inner surface was used. The pool was filled with opaque water (water mixed with skimmed milk) maintained at a temperature of 22 ± 1℃. The height of the water was adjusted so that the escape platform (8 cm diameter) was 1.5 cm above the water level. The experimental animals were placed in different starting positions, away from the escape platform, and escape latency time was measured using a video tracking system. A 5-day training period was implemented, in which day 1 involved the experimental animals (C57BL/6J mice) being placed in the water bath and allowed to freely swim for 90 seconds (s), without the escape platform. Day 2 to day 5 consisted of a cognitive adaptation period performed 9 times, at intervals of 30 s, twice a day. Mice were placed in the pool facing the wall, at different starting points and allowed to freely swim to the escape platform. If mice were unable to locate the platform within the 90 s allocated, they would be aided to the platform by the experimenter. Mice were kept on the platform for 15 s in order that the escape platform location may be remembered. Training trial memory was tested in scopolamine-induced mice, 30 min after 5 mg/kg i.p. administered scopolamine, and ethanol-induced mice, after the training period. The test trial measured the time taken to reach the quadrant where the platform was located after removing the platform from the round tank. The test trial was performed over 3 days, with the same method as the training trial and a maximum time of 90 s allocated to find the platform location.
Passive avoidance test
The apparatus used for the passive avoidance (PA) test was an acrylic shuttle box (53 × 44 × 33 cm, electric grid floor, Ugo, Italy) with two internal compartments and a guillotine door dividing the two compartments (10 × 10 cm). Stainless steel grids were installed at the base of the dark compartment to produce a scrambled 3 mA footshock to the mice. The light compartment was illuminated by a 20 W bulb and the noise control was limited to less than 60 dB throughout the experiment.
The PA test was used to measure the working memory ability of memory-impaired C57BL/6N mice according to a modified method used by Oklejewicz et al. [16]. The test involved the animal being placed in the lighted compartment (1,500 lux) for 30 s. After 30 s, the compartment guillotine door was opened and the time spent in the light room was measured. Once the animal had fully crossed over to the dark compartment the guillotine door would automatically close and a 3 mA foot shock for 3 s would be given. A training trial was repeated 5 times to habituate the animals to the relationship between the foot shock and entering the darkened compartment. Mice were repeatedly returned to the lightened side until they didn't move for up to 120 s. 24 h after the training trial was completed the test trial was performed, in which the step-through latency times were measured up to a cut-off time of 300 s.
Biochemical analysis of blood
After 4 weeks of treatment, all experimental animals were fasted for 12 h, anesthetized with 75% CO2 and 25% O2, followed by blood sample collection. Blood was centrifuged at room temperature for 30 min and then centrifuged at 3,000 rpm for 15 min at 4℃. Aspartate aminotransferase (AST), alanine aminotransferase (ALT), triglyceride (TG) and total cholesterol (TC) were analyzed by a serum biochemical analyzer (Roche Ltd., Basel, Switzerland).
Brain tissue sampling and Western blot analysis
Following blood sampling, the guillotine method of decapitation was carried out, under trained personnel, to remove the brain from the skull. To carry out protein analysis, the hippocampal cells were homogenized in cell lysis buffer for protein extraction and then centrifuged at 800 rpm for 10 s (4℃) to allowed supernatant protein concentration to be used. Before 24 h of ice incubation, one round of centrifugation at 14,000 rpm for 20 min (4℃) followed by protein quantification, using the Bradford assay kit, was carried out. The proteins were then resolved by SDS-PAGE (10%) (Polyacrylamide Gel Electrophoresis), transferred to a polyvinylidene fluoride membrane using the semi-dry transfer system (Bio-Rad, USA) and treated with a blocking solution (5% skim milk, 1 × TBST buffer) for 1 h. The membrane was then washed three times for 10 min in TBST, incubated overnight (4℃) with primary antibodies against mAChR, CREB, p-CREB, ERK1/2, p-ERK1/2, and then washed again with the aforementioned procedure. After treatment with the buffer, protein band intensity was detected using enhanced chemiluminescence (Chemi-Doc, BIORAD XRS system) and compared with GAPDH, CREB, and ERK1/2after activation using a Western blot detection kit (Abfrontier, WEST SAVE GOLD).
Immunohistochemistry staining
Brain tissue was harvested and fixed for 24 h in a fixed solution (post-fixation), and then sequentially exchanged for 12 h in 10% and 20% sucrose solution, respectively, and then stored in 30% sucrose solution for 3 days. Frozen sections were cut at 30 µm thickness using a cryotome (Leica CM1850 cryostat, Germany) and stored at −80℃ until immunohistochemical staining. The cut tissues were washed twice with PBS for 10 min, and then incubated with 2% Triton X-100 for 5 min and 0.6% H2O2 for 5 min followed by washing with PBS twice for 10 min. They were then transferred to a pre-incubation solution containing 2% goat serum for 30 min to block non-specific IgG binding, and then incubated overnight (4℃) in a solution containing a primary antibody for p-CREB (ser133) diluted 1:250 (Abcam, U.K.). The biotinylated secondary antibody was stained for 1 h at room temperature, then washed three times with PBS for 10 min and incubated with avidin-biotin peroxidase complex for 1 h at room temperature. After that, the cells were stained with 0.02% 3.3′-diamino benzidine tetrahydrochloride (DAB) solution containing 0.003% hydrogen peroxide for 10 min at room temperature and observed under an optical microscope (Nikon, Japan). P-CREB (ser133) antibody was purchased from Abcam (UK) and photographed at a magnification of 100 × under a microscope.
Acetylcholinesterase activity
In order to determine the inhibitory effects of donepezil and FL50 on AChE activity in memory-impaired mice, AChE was assayed using a modified method of Ellman [17]. The mixture was prepared by mixing 0.1 M phosphate buffer (pH 8.0) 2.6 mL, 10 mM Ellman's solution (10 mM 5,5′-dithio-bis-2-nitrobenzoic acid, 15 mM sodium bicarbonate) 100 mL and zymogen 0.2 mL, transferred into separate wells, and the product of thiocholine reaction with 5,5′-dithio-bis-2-nitrobenzoic acid determined at 410 nm. Following this, 20 uL acetylcholine iodide (75 mM) (Sigma-Aldrich Co., USA) was added to the reaction mixture and change in absorbance was measured at 2-minute intervals. The enzyme activity was determined by repeating the protocol 3 times.
Acetylcholine concentration
Acetylcholine content was measured based on the σ-acyl induced reaction of water with alkaline hydroxylamine by the method of Hestrin [18]. After mixing 50 µL of 1% hydroxylamine with 50 µL of brain homogenate, 500 µL of FeCl3 (10% in 0.1N HCl) was added, and the content of acetylcholine was measured at 490 nm.
Statistical analysis
Data presented are mean ± standard deviation (SD). All analyses were performed using Statview software. The effect of treatment on behavioral, biochemical and immunohistochemical data were analyzed by one-way analysis of variance (ANOVA) followed by post hoc analysis using the Fisher's PLSD test.
RESULTS
Biochemical analysis of blood
The effects of FL50 administration on liver toxicity, and triglyceride and total cholesterol levels are illustrated in Fig. 2. The results of liver toxicity measurements following a 4-week administration of EtOH demonstrated a significant increase in ALT and AST concentration (Fig. 2A and B). Treatment with 50 mg/kg of FL50 extract significantly reversed the effects of EtOH in the EtOH + FL50 group (P < 0.05). Conversely, in the case of the treatment groups, TG and TC levels remained in the normal range (Fig. 2C and D).
Morris water maze test
The effects of FL50 administration on scopolamine and EtOH-induced memory damage, following a 4-week treatment course, on hippocampus-dependent spatial learning ability was assessed by the MWM test. These results are illustrated in Fig. 3. The Sco group took 46.58 ± 23.77 s to escape the water maze test, while the EtOH group took a comparable 50.67 ± 8.14 s to escape. The normal CON group recorded an escape time of 19.92 ± 7.24 s, which is significantly less time than the Sco and EtOH induced dementia groups (P < 0.05 and P < 0.001, respectively). Donepezil and FL50 treatments both improved the escape times of memory-impaired mice. Compared to the Sco- and EtOH-induced memory impairment groups, FL50 administration significantly reduced escape times (P < 0.05 and P < 0.001, respectively), with the same effectiveness of donepezil treatment.
Passive avoidance test
The PA test was used to assess the impact of FL50 on short-term memory impairment induced by both scopolamine and EtOH administration. Fig. 4 illustrates the time taken for memory-impaired mice to inhibit behavior in order to avoid a 3 mA shock. The time spent remaining in the white compartment in the Sco and EtOH-induced dementia groups were 49.00 ± 19.52 s and 51.06 ± 13.30 s, respectively; which was significantly less time than the normal CON group (P < 0.001). Treatment with donepezil significantly increased the step-through latency time in both the Sco + Dpz and EtOH + Dpz groups (148.87 ± 25.45 s and 134.80 ± 20.96 s, respectively). Administration of FL50 also significantly improved step-through latency time, but to a much greater extent in the EtOH + FL50 than Sco + FL50 group (157.73 ± 27.80 s and 72.87 ± 25.08 s, respectively).
Acetylcholine and acetylcholinesterase activity
Scopolamine induced dementia significantly reduced ACh activity, in comparison to the CON group (0.493 ± 0.007 and 0.922 ± 0.057 µ mol/mg protein, respectively). 4-week, daily consumption of 2 g/kg of ethanol also reduced ACh activity by almost 50% (0.540 ± 0.041 µ mol/mg protein), compared to CON (P < 0.001). However, FL50 administration significantly reversed the damage caused by scopolamine-(P < 0.05) and EtOH-induced (P < 0.05) dementia, demonstrating similar effectiveness to donepezil treatment (Fig. 5A). Elevated acetylcholinesterase activity is important in characterizing the effects of dementia on the brain; as it is responsible for catalyzing the breakdown of acetylcholine neurotransmitter. Donepezil is a representative AChE inhibitor - reversing cognitive deficit by increasing cholinergic activity degenerated by Alzheimer's disease and dementia. FL50 administration notably reduced AChE activity in both the Sco + FL50 (P < 0.001) and EtOH + FL50 (P < 0.001) dementia groups (Fig. 5B).
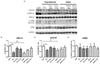
Brain tissue protein expression: mAChR, ERK1/2, CREB
Western blot was used to determine the effects of FL50 administration over 4 weeks on memory-related protein expression in brain tissue in dementia induced mice. The results from mAChR, ERK1/2 and CREB protein expression are shown in Fig. 6. Western blot analysis of the Sco- and EtOH-induced dementia groups showed a significant decrease in the expression of mAChR (P < 0.05) and ERK1/2 (P < 0.001) in the brain, but had no significant effect on CREB protein expression (Fig. 6A). In the case of ERK1/2, both donepezil and FL50 treatment significantly promoted hippocampal expression (P < 0.05) (Fig. 6B), whereas mAChR expression, was improved, but without significance (Fig. 6C), and CREB protein expression was unchanged as a result of either intervention (Fig. 6D).
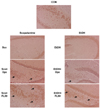
Brain tissue p-CREB (Ser133) immunohistochemistry
Fig. 7 illustrates the results of FL50 administration on the expression of p-CREB (Ser133) in the hippocampus of the brain, associated with memory. As a result of scopolamine and EtOH-induced dementia, protein expression of p-CREB (Ser133) in the hippocampus was decreased. However, treatments of donepezil and FL50 both increased the expression of p-CREB, indicated by the arrows in Fig. 7.
DISCUSSION
Laminaria japonica is a type of sea tangle rich in vitamins, minerals, essential fatty acids, carotenoids and bioactive compounds. Supplementation of fermented Laminaria japonica has been reported to provide a variety prophylactic benefits, including antimutagenic [19] hepatoprotective [20] and antioxidant properties [21], and improved cognitive function [14], among others. A specialized fermentation process, using Lactobacillus brevis BJ20, enriches the GABA content and enhances the bioactive properties of Laminaria japonica. We investigated whether FL50 can be a viable natural intervention for Sco- and/or EtOH-induced dementia model mice, in comparison with the current standard treatment for dementia. The results of the present study firstly demonstrate that liver damage induced by EtOH consumption can be reversed with administration of FL50, as shown by a significant reduction in ALT and AST after treatment. Regarding short-term memory impairment and learning ability, 4 weeks of FL50 treatment reduced the step-through latency time in the PA test, with a bias toward EtOH-induced dementia. Hippocampal-dependent spatial learning ability in C57BL/6J mice was also improved by FL50 - to an equivalent degree as conventional donepezil administration - demonstrated by reduced escape latency time in the MWM test. The anticholinergic effects of Sco and EtOH on the brain were notably reversed by FL50, through the attenuation of AChE activity and elevation of ACh concentration - providing protection against central cholinergic pathway dysfunction and potential memory decay. FL50 also showed promise as a defense against neuronal signaling pathway compromise, by elevating ERK1/2 protein expression (Western blot analysis) and increasing the expression of p-CREB (ser133) (immunohistochemical analysis) in hippocampus brain tissue.
AST and ALT levels were used in the present study as indicators of liver damage or injury. It is widely known that excessive alcohol consumption can lead to alcohol-induced liver disease, and remains the leading cause of death and mortality in individuals in mid-life [22]. The hepatoprotective properties of FL have been demonstrated in previous studies, with the focus being on ethanol-induced oxidative stress and the antioxidant properties FL possesses in preventing/inhibiting membrane peroxidation, while increasing antioxidant species in liver tissue [202324]. Initially, our results can confirm the damage associated with alcohol consumption, but also the viability of FL as a safe and effective intervention for the attenuation of hepatotoxicity.
Passive avoidance memory (PAM) involves associative learning (explicit), operant-like conditioning to the shock, and a spatial memory component, where the mouse remembers the location in which it received the foot shock [25]. This inhibitory avoidance depends on the activation of the cholinergic system, which originates in the basal forebrain and projects to the hippocampus, releasing ACh, which impinges on the hippocampal mAChRs and modifies neuronal activity - confirmed by PAM impairment in response to pre- and post-training administration of mAChR antagonists, and promotion following mAChR agonists administration [262728]. Several studies have convincingly demonstrated the recall impairment (1 h or 24 h after training) associated with Sco (a non-selective mAChR antagonist) administration [293031]. Furthermore, ethanol neurotoxicity has been related to loss of hippocampal pyramidal neurons, cholinergic neurons in the basal forebrain and inhibition of NMDA (involved in long-term potentiation) [4] that coincide with impairments to memory and learning [5]. Hence, the upregulation of ACh neurotransmitter and decrease in AChE activity, along with improved step-through latency time in the EtOH + FL50 group, supports a mechanism for FL50 action on the cholinergic system involved in the formation of short-term memory - particularly, impaired by EtOH exposure. In addition, previous studies have highlighted an integrated mechanism between the stimulation of mAChRs and ERK1/2 transduction pathway activation in PAM-type tasks [323334]. Giovannini et al. [29] demonstrated that, during and immediately after PAM acquisition an increase in ACh release by the activation of ERK in CA1 hippocampal neurons, and memory formation occur. It is possible that FL50 acted on the cholinergic system via this mechanism (discussed below), and from the immunohistochemistry results, potentially activated CREB transcription factor leading to memory encoding [2535], and even neuronal protection [3637]. In accordance with Park et al. [14], the present study also demonstrated that FL50 reduced escape latency time in the MWM test following Sco- and EtOH-dementia, indicating an improvement in short-term hippocampus-dependent spatial memory - particularly impaired in AD [2]. We can, therefore, confirm that FL50 is an efficacious intervention for Sco- and EtOH-induced AD or dementia-related impairments in learning and memory, and likely related to hippocampal synaptic plasticity.
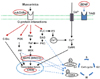
The present study investigated hippocampal mAChR, ERK1/2 and CREB expression in response to Sco- and EtOH-induced dementia, to evaluate the effect of FL50 on protein factors in brain tissue and the subsequent impact on cognitive ability. The mAChR is a G protein-coupled muscarinic acetylcholine receptor in the cell membrane of neurons [38], known to promote long-term potentiation of the postsynaptic membrane and induce Ca2+ mobilization and inositol trisphosphate production. Of its 5 subtypes (M1–M5), the M1 muscarinic receptor subtype, predominantly expressed in the central nervous system, has been implicated in the impairment of cognitive ability. M1 mAChRs have been demonstrated to potentiate NMDA receptor signaling in the hippocampus and cortex [3940], while M1-preferring mAChR antagonist pirenzepine impaired accuracy and acquisition of tasks in PA, Morris water maze, and visual discrimination in rats [4142]. On the other hand, CREB is a cAMP response element-binding transcription factor that is regulated by BDNF and tyrosine hydroxylase and is known to be involved in long-term memory formation [354344]. The action of mAChRs also regulates CREB by modulating MAPK (ERK1/2), triggering a cascade signaling pathway, and creating conditions favorable for neuronal plasticity - ERK1/2 is thought to trigger a highly important signal transduction pathway in short-term memory formation [2545]. We have previously shown FL to induce a rise in BDNF levels [9], while Park et al. [14] reported recovered choline acetyltransferase, CREB and BDNF immunopositive neurons in the hippocampusas a result of Saccharina japonica (also known as Laminaria japonica) administration. Taken together, FL50 administration may regulate, in part, the promotion of ERK1/2 via a BDNF-dependent TrkB transduction mechanism [4446], and/or by targeting the dysregulation of cholinergic signaling in the brain, via ACh neurotransmitter elevation and AChE inhibition (Fig. 8). This would suggest a similar mechanism to that of current primary treatments for AD patients, including donepezil, tacrine, rivastigmine and galantamine - nonselective AChE inhibitors. These drugs have been tested and show efficacy in treating AD-related cognitive symptoms, however not without the side effects including, night time disturbances [7], gastrointestinal [6] and cardiovascular complications [8].
The implications of this study are meaningful in contributing to the development of cholinergic therapeutics that can provide clinical efficacy without the unwanted side effects. It is worth noting that although 4 weeks of EtOH administration caused damage to cognitive ability, it may not have been sufficient to cause inhibition of transcription factor (CREB) expression, potentially resulting in a strong central nervous system disorder. Nonetheless, neuronal damage was incurred following EtOH consumption. This limitation to our study may be redressed in future research into the effects of FL50 on long-term alcohol-related disorders.
In conclusion, treatment of FL50 reduced Sco- and EtOH-induced spatial learning and memory impairments as demonstrated by the MWM test, however, aversive stimulus avoidance in the PAM test was specifically improved in EtOH-induced dementia. This poses a question for further study: which memory types may be improved by FL50 administration? FL50 treatment had a neuroprotective effect against Sco- and EtOH-induced inhibition of ACh, ERK1/2 and p-CREB (ser133), and upregulation of AChE. Based on these findings, FL50 may be considered an effective intervention for scopolamine-induced dementia model animals, and alcohol-related amnesia and dementia, concerning the reverse of cognitive impairment and neuroplastic dysfunction.






 XML Download
XML Download