

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The quantity and quality of sleep are related with positive influences of life scores, are a common public health issue globally, and are closely affected by internal and external factors [1]. Studies have shown that the quality and quantity of adequate sleep (adequate sleep is defined as 6–8 hours per night regularly) are closely related to future health and well-being [2]. Sleep disorder is the most commonly reported sleep-associated problem and can be characterized in terms of duration (acute or chronic) [3]. The process of sleep and sleep-related behavior are modulated by a variety of neurochemicals operating through distributed neuronal circuits. Studies have concentrated on the pharmacological and physiological implications of specific receptor binding sites to design new drugs to treat sleep disorders during the past few decades [4]. Sleep-wake regulatory mechanisms of the central nervous system are also targets for the treatment of sleep disorder. Sleep disorder patients depend on pharmacological treatments even though these chemical agents have various side effects such as difficulty keeping balance and overdosing. For this reason, sleep disorder researches have focused on sleep-promoting properties of natural substances and biomolecule compounds to avoid adverse side effects [56].
Animal models have been used to understand the most common human sleep disorders such as insomnia, narcolepsy, restless legs syndrome, and sleep apnea [7]. In addition, evaluation of non-pharmacological treatments for sleep disorder will require various applied methods in animals for the development and preliminary testing of new therapeutic approaches, and for the identification and validation of sleep-inducing mechanisms. The fruit fly, Drosophila melanogaster, is a model insect with a wealth of information on circadian rhythm and sleep-related neuronal systems. The brain levels of acetylcholine, octopamine, dopamine, glutamate, and receptors for γ-aminobutyric acid (GABA) and serotonin (5-HT) are expressed in the mushroom bodies of the fly and modulate sleep/wake behavior changes [89]. Moreover, the fly shares an evolutionary conservation and many features with mammalian sleep, and fly sleep is controlled homeostatically by the mechanisms of sleep regulation [10]. To record sleep in flies and test sleep disorder treatment requires measuring the circadian behavior of locomotor activity of the flies from the field [11]. Moreover, preclinical studies are recommended for the reliable assessment and the validation phase of candidate treatment, with compounds generally administered to rodents, such as mice and rats. Mice and rats are the most commonly used model system for studying sleep-related behaviors, and the function of sleep and molecular processes [12]. In addition to electrophysiological measures in freely moving mice and rats under controlled laboratory conditions, sleep quantity and quality can be quantified behaviorally by measuring movement and sleep architecture. In humans and animals, the assessment of sleep architecture depends on both non-rapid eye movement (NREM) sleep (light sleep and slow-wave sleep, SWS) and rapid eye movement (REM) sleep, as well as electroencephalogram (EEG) wave patterns [13].
Although data obtained from invertebrate and vertebrate model systems have assessed the treatment properties that underlie sleep-wake cycles and neurochemical regulation, additional experimental models are needed to evaluate the interaction of behavioral changes and candidate hypnotic compounds with neuronal system. Caffeine (1,3,7-trimethylxanthine) is a psychoactive substance in many popular beverages and has been used with the purpose of maintaining physical activity and mental alertness. Even though caffeine overdose known to be associated with a variety of side effects such as tachycardia, hypertension, nausea, vomiting and gastritis, administration of caffeine has been proposed as an effective agent to model sleep-onset insomnia by antagonizing the function of all adenosine receptors (Ars: A1, A2A, A3, and A2B) [1415]. Targeting the adenosine system with caffeine treatment is an effective tool for controlling individual weakness to the long-term effects of sleep deprivation on cognitive ability and sleep [16]. Also, previous studies have described the effects of caffeine on the cellular and behavioral levels in animal species, and chronic administration of caffeine causes a wide range of biochemical alterations in the central nervous system (CNS), including serotoergic, nicotinic, and GABAA receptors [17]. Caffeine is also known to influence the onset of sleep, total sleep time, sleep quality and efficiency, such as stage 3–4 NREM sleep [18]. In this study, caffeine was used as a stimulant to promote wakefulness and simulate the insomnia condition in invertebrate and vertebrate models to evaluate the sleep-promoting effects of two combined amino acids.
MATERIALS AND METHODS
Animals
D. melanogaster Canton-S strain was obtained from the Bloomington Drosophila Stock Center (Bloomington, IN, USA). The flies were reared on Jazz-Mix Drosophila food (Fisher Scientific, Waltham, MA, USA) at 25 ± 1℃ on a 12:12 h light:dark cycle at 60% relative humidity (RH). Prior to treatment, each sample of flies (3-5 day-old male) were anesthetized for transfer by using CO2. ICR mice (4-weekold male) and SD rats (8-week old male) were obtained from Orient Bio Inc. (Gyeonggi-do, Korea). All rodents were maintained in acrylic cages with food and water at 24 ± 2℃ in 50-60% atmospheric humidity with a 12:12 h light: dark cycle. Following the 1week adaptation and recovery period vertebrate models were used for pentobarbital-induced sleep test and measurements of EEG wave patterns. The Korea University Animal Care Committee (Seoul, Korea) approved the use of animals in this protocol (KUIACUC-2015-102).
Behavioral assays
Caffeine, GABA, and 5-HTP (Sigma Aldrich, St. Louis, MO, USA) were mixed in sucrose-agar media (Normal diet: 5% sucrose and 1% agar) for the behavioral assay. For a video-tracking system, flies (10/group) were treated with GABA (10 mg/mL), 5-HTP (1 mg/mL), or combined GABA/5-HTP (10 mg/mL and 1 mg/mL each) with caffeine (1 mg/mL). A 1% caffeine concentration was used as a stimulant in the awake condition [19]. The flies acclimated to the chamber for 1 min, followed by video recording for 5 min 1 h before subjective nighttime. The chamber consisted of a circular arena (8 mm in diameter and 0.1 mm in height) which was laid down a white background. Analysis of fly activity was performed by the Noldus EthoVision-XT system (Noldus Information Technology, Netherlands).
Pentobarbital-induced sleep test
The experiments were analyzed between 13:00 and 17:00, and all mice were fasted for 24 h prior to testing. During oral administration, the individual treatments were not revealed to the observers. GABA (14/group, 60 mg/kg), 5-HTP (14/group, 6 mg/kg), and combined GABA/5-HTP (15/group, 60 mg/kg and 6 mg/kg each) were dissolved in 0.5% physiological saline. All samples were administered (post-oral injection, p.o.) with caffeine (10 mg/kg) and pentobarbital (sub-hypnotic dose: 30 mg/kg, intraperitoneal injection [i.p.]; hypnotic dose: 42 mg/kg, i.p.) was injected 1 h later to induce sleep. Following the pentobarbital injection, mice were laid down on their back in individual cages and sleep latency was recorded from this time-point. In addition, sleeping duration was analyzed, which were defined by loss of righting reflex, to recovery and beginning to move. After injection, mice that did not sleep within 15 min were excluded from the experiment.
Surgical procedures and electrophysiological analysis
Under isoflurane (Troikaa Pharmaceuticals Ltd, Gujarat, India) anesthesia in the mixture of oxygen and nitrous oxide, rats were placed to a stereotaxic instrument frame (Stoelting Inc., Wood Dale, IL, USA). The body hair of the head was removed using a shaver and cleaned to ensure disinfection with 70% ethanol before a dorsal midline incision was made in the scalp to expose the skull. Hemostasis was achieved with sterile cotton-tip applicators and bregma was marked and four holes bored through the skull +1 mm lateral to midline/+1 mm to bregma, and −1 mm lateral to midline/−3 mm to bregma for the hippocampus, frontal cortex, and striatum, respectively [20]. The EEG electrodes with a mounting screw and socket were fixed to the skull surface of rat with dental cement (AgnThós AB, Lidingö, Sweden). Seven-day post-surgery (recovery period), rats were divided into normal and control and treatment groups. Total experiments periods were recorded 1 h than with later administration of caffeine (8/group, 10 mg/kg) and the two amino acids (8/group, GABA: 60 mg/kg, 5-HTP: 6 mg/kg) and combined GABA/5-HTP (8/group, 66 mg/kg) and performed between 10:00 and 18:00. The EEG activity was acquired by Iox2 data acquisition software (version 2.8.0.13, emka Technologies, Paris, France) and EEG spectra were analyzed in 1 Hz bins. Setting for standard bands were the following: δ wave, 0.5–4; θ wave, 4–9; α wave, 9–12; β wave, 12–30; and gamma: 30–60 Hz. Analysis of EEG was calculated wave patterns, wake time, and sleep time on the recording data at 2 sec intervals by fast Fourier transform of ecgAUTO3 software (version. 3.3.0.20, emka Technologies).
Statistical Analyses
The Statistical Package for Social Sciences version 12.0 (SPSS Inc., Chicago, IL, USA) was used to analyze the data. Data from the caffeine-induced sleepless model systems are represented as mean ± standard error (SE). Differences between groups were evaluated by one-way analysis of variance (ANOVA) with Tukey's and Dunnett's multiple comparison test.
RESULTS
Effects of two amino acids and combined GABA/5-HTP on the distance moved, turn angle, and mobility
The effects of two amino acids and combined GABA/5-HTP on the distance moved, turn angle, and mobility in the caffeine-induced sleepless fly are presented in Fig. 1. The control group (flies treated with the caffeine) traveled a significantly greater distance (1.61-fold, F(4,85) = 5.57, P < 0.01) compared to the normal (untreated) group (Fig. 1A). Administration of GABA (0.69-fold, P < 0.05) and 5-HTP (0.69-fold, P < 0.05) significantly decreased the distance moved compared to the control group. Furthermore, the distance moved was significantly decreased by combined GABA/5-HTP (0.52-fold, P < 0.001) compared to control group. As shown in Fig. 1B, the turn angle provides the angle formed by the change in the direction of the individual fly movement. However, there was no significant difference in the turn angle between groups. In Fig. 1C, mobility is defined on the basis of changes in the pixels caused by the spatial displacement of individual flies. The control group showed significant increased mobility (1.40-fold, F(4,85) = 5.34, P < 0.01) compared to the normal group. Two amino acids and combined GABA/5-HTP groups showed significantly reduced mobility compared to the control group (P < 0.01).
Effects of two amino acids and combined GABA/5-HTP on meander and velocity
The meander measure represents the change in the direction of the individual fly movement of a subject relative to the distance moved by the subject (Fig. 2A). Administration of caffeine, and caffeine with two amino acids and combined GABA/5-HTP did not significantly change the meander of individual flies compared to the normal group. Fig. 2B shows the walking speed (velocity) during the 5-min observation period; this is a measure of the activity of the individual flies. Caffeine-treated flies showed significantly higher walking speeds compared to normal flies (1.32-fold, F(4,85) = 4.27, P < 0.05). Although GABA treatment did not affect velocity, administration of 5-HTP and combined GABA/5-HTP caused a significantly lower velocity compared to the control group (P < 0.01).
Effects of two amino acids and combined GABA/5-HTP on pentobarbital-induced sleeping behaviors
The effects of two amino acids and combined GABA/5-HTP on sleep latency and duration of caffeine-induced sleepless mice induced by a sub-hypnotic and hypnotic dosage of pentobarbital are shown in Table 1 and Fig. 3, respectively. Oral administration of caffeine significantly decreased the sleep onset ratio and duration compared to the normal group (F(4,66) = 21.96, P < 0.01). The sleep onset ratio and duration were significantly increased by Two amino acids and combined GABA/5-HTP compared to the control group (Table 1). Particularly, the administration of combined GABA/5-HTP increased the change of the sleep onset ratio and duration when compared to the control group. In mice treated with the hypnotic dose of pentobarbital (Fig. 3), caffeine administration significantly increased sleep latency (F(4,45) = 6.46, P < 0.001) and decreased sleep duration (F(4,45) = 21.54, P < 0.001). In addition, sleep latency was significantly decreased (GABA and combined GABA/5-HTTP: P < 0.001, 5-HTP: P < 0.01) and the duration of sleep was prolonged by administration of two amino acids and combined GABA/5-HTP compared to the control group (P < 0.001).
Effects two amino acids and combined GABA/5-HTP on sleep/wake architecture
The processed recording per 8 h and sleep/wake architectures of two amino acids and the combined GABA/5-HTP administration in a caffeine-induced awake model are presented in Fig. 4. Doses for the caffeine-induced awake model were chosen based on previous literature [21]. Caffeine administration (10 mg/kg) to rats caused significant reductions in sleep time (F(4,35) = 10.68, P < 0.05) and parallel increases in wake time (F(4,35) = 11.36, P < 0.01) compared to normal rats. Caffeine combined with GABA/5-HTP administration caused a significant reduction in the wake time (P < 0.05) and a parallel increase in the sleep time compared to caffeine administration (P < 0.05). There was no significant change in REM sleep time in response to the combined GABA/5-HTP treatment. The only significant effect of the combined GABA/5-HTP administration was an increase in the NREM sleep time compared to caffeine administration (F(4,35) = 15.41, P < 0.05).
DISCUSSION
Although questions at the behavioral and electrophysiological levels regarding the caffeine-induced sleepless model system remain unaddressed, the present study, based on behavior investigations and EEG wave patterns observations, provides a convincing explanation for the inhibitory effects of two amino acids and combined GABA/5-HTP on sleep disorder in animals. Currently, most studies of hypnotic drugs and candidate treatments focus on GABAergic inhibitory synaptic transmission, the GABAA receptor-binding site in the brain and selective 5-HT2A receptor inverse agonist remains a target of sleep disorder treatments [2223]. In the fruit fly, downregulation of slow metabotropic GABAB receptors in the pigment dispersing factor-positive ventrolateral clock neurons using RNAi reduced sleep maintenance in the second half of the night [24]. In addition, GABAB receptor agonists are potentially useful in schizophrenia and insomnia related to schizophrenia, and are involved in inhibiting dopamine release and regulating glutamatergic systems of dopamine [252627]. Arnaud et al. [28] assessed sleep-waking regulation of GABAC receptors, which have a higher sensitivity than other GABA receptors, using (1,2,5,6,-tetrahydropyridine)-methylphosphinic acid, a specific antagonist, in rats. Five-HTP is a serotonin precursor and easily crosses the blood brain barrier without a transport molecule [29]. Moreover, 5-HTP regulates sleep-wake behavior and 5-HT receptors interact with GABAergic inhibitory synaptic transmission in flies and mammals [3031]. The polysomnography data support an increase in sleep maintenance following treatment with several selective 5-HT2A receptor antagonists [32]. In a previous study, we also evaluated the dose-dependent sleep-promoting effect of two amino acids and combined GABA/5-HTP on locomotor activity and the quantity and quality of sleep in animal models [3334].
In the present study, we utilized the caffeine-induced sleepless Drosophila to investigate the inhibitory effect of two amino acids and combined GABA/5-HTP and regulated significant difference in locomotion such as distance moved, turn angle, mobility, meander, and velocity (Fig. 1 and 2). Following an action potential, GABA is released to terminate GABA-stimulated responses from presynaptic neurons into the synaptic cleft in both vertebrate and invertebrate nervous systems [35]. The GABAergic system is the target of pharmacological approaches and GABA transport inhibitors resulted in diminished locomotor activity [36]. D. melanogaster locomotor activity follows circadian regulation and biochemical alterations [11]. In addition, previous reports suggested that 5-HTP increases serotonin levels in the CNS of both insects and mammals, and doses of 5-HTP treatment from 1 mg/mL to 5 mg/mL and 2 mg/kg to 200 mg/kg have been found to significantly increase sleep time [937383940].
This study was designed to validate the inhibitory and sleep-promoting effects of combined GABA/5-HTP and demonstrate that the caffeine-induced sleepless model is necessary to evaluate sleep at both the behavioral and electrophysiological levels. Caffeine (10 mg/kg) was used to simulate the insomnia condition in pentobarbital-induced sleep test and electrophysiological analysis [41]. To understand the inhibitory effect of combined GABA/5-HTP on behavioral changes, we used the pentobarbital-induced sleep test for evaluating caffeine-induced sleepless mice (Table 1 and Fig. 3). Caffeine (control group) significantly decreased sleep onset and duration relative to the normal group at the sub-hypnotic dose pentobarbital. However, although sleep behavior depends on caffeine administration, the combined GABA/5-HTP treatment plus caffeine group showed significant differences compared to the control group. In addition, administration of caffeine alone with the hypnotic dose showed a significant arousal effect compared to the normal group, while two amino acids and combined GABA/5-HTP significantly increased total sleeping time compared to mice treated with caffeine only (Fig. 3B). Other evidence from the pentobarbital-induced sleep test has shown that caffeine produces a dose-dependent increase in sleep latency and decrease in sleep duration. Kuribara et al. [42] assessed the interaction of caffeine with ethanol, diazepam, and pentobarbital, and classified them as CNS depressants or stimulants by observing the ambulatory activity of mice. In addition, the hypnotic effect, and the interaction of caffeine with pentobarbital, was studied in 42 medical and surgical patients [43]. In male NIH Swiss strain mice, chronic ingestion of caffeine increased the densities of cortical 5 HT1 and 5 HT2 serotonergic receptors (26–30%) and benzodiazepine-binding sites associated with GABAA receptors (65%) [44]. Taken together, administration of caffeine produced arousal effects and a wide range of biochemical alterations in the CNS and the combined GABA/5-HTP treatment produced inhibitory effects by binding to upregulated GABAergic and serotonergic receptors.
We also investigated the inhibitory effect of the combined GABA/5-HTP on sleep/wake architecture in the caffeine-induced sleepless rats (Fig. 4). Oral administration of combined GABA/5-HTP increased total sleep time and NREM sleep compared to the control group. Paterson et al. [21] assessed the dose-dependent effect of caffeine on all sleep parameters using EEG analysis in SD rats and discussed the merits of using caffeine as an insomnia model. In addition, according to the report by Lazarus et al. [45] caffeine acts to produce motivational and motor responses identified using focal RNA interference to silence the expression of adenosine A2A receptors in the rat brain. Caffeine administration led to increased wakefulness and reduced EEG slow wave activity during subsequent sleep periods [4647]. In contrast, the GABAA receptor agonist THIP (4,5,6,7,-tetrahydroisoxazole [5,4-c] pyridine-3-ol), a muscimol derivative, enhanced spectral power in the low-frequency range of EEG and affected EEG slow wave activity patterns in both waking and NREM sleep periods in rodents and humans [48]. In our previous study, GABA showed ‘temporary but unstable’ sedation and 5-HTP had a ‘delayed but stabilized’ effect on flies. The combination of GABA and 5-HTP can support stabilization and delayed effects [3334]. The GABA re-uptake inhibitor tiagabine dose-dependently elevated the lower frequency waves during NREM sleep [49]. In addition, studies have shown that the serotonergic system and 5-HT level are linked to slow wave activity generation and that 5-HT receptors are involved in GABAergic interneuron and motoneuron modulation [505152].
In conclusion, using video tracking to measure behavior, as well as electrophysiological approaches, we identify that two neuronal substances and their mixture enhance the quantity and quality of sleep in the caffeine-induced sleepless model system. The combined evidence suggests that the caffeine-induced sleepless model is a valid system for investigating the inhibitory and sleep-promoting effects of non-pharmacological remedies, and the combined GABA and 5-HT precursors can be helpful in patients with sleep disorder who have mild or moderate insomnia. The combined GABA/5-HTP could be a potential medication that acts at low doses with fewer side effects on behavioral and neurochemical responses.
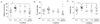

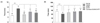


 XML Download
XML Download