

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Our recent reports showed a close interaction between inflammation and reactive oxygen species (ROS) / reactive nitrogen species (RNS) induced oxidative damage,123 namely, the inflammatory process is activated by diverse reactive species (RS), e.g., ROS and RNS. The inflammation process triggers a chain of events that forms up-regulating RS, and pro-inflammatory cyclooxygenase (COX) is a key enzyme in the prostaglandin (PG) synthesis pathway that is a major RS-producing factor. Additionally, leukotrienes, key mediators of the inflammatory process, are catalyzed by 5-lipoxygenase (5-LO) during the first two crucial steps of leukotriene biosynthesis from arachidonic acid.4
Numerous reports reveal that various RS are causally related with cellular oxidative insults.1 The primary ROS/RNS species generated in a cell are superoxide (•O2), hydrogen peroxide (H2O2), and nitric oxide (NO•); and •O2 is able to rapidly react with NO• to form toxic peroxynitrite (ONOO–) or dismutate to form H2O2.15 To counteract ROS/RNS induced oxidative damage, cells have enzymatic and non-enzymatic defense mechanisms.5
Lipopolysaccharide (LPS) is known as a powerful macrophage activator, and LPS treated RAW264.7 cells are a well-defined cellular model of macrophage activation at sites of inflammation, which generating various RS including ROS and RNS.67
Flavonoids have gained much attention in the food and pharmaceutical industries due to their various health promoting effects,89 which are beneficial for human diet and food nutrition.1011 One of these flavonoids, rhamnazin is revealed to have anti-oxidative, anti-bacterial, antiangiogenic, and anti-tumor capacities.1213 In addition, previous study showed that rhamnazin revealed powerful anti-oxidative properties either by directly acting as a potent antioxidant or by enhancing anti-oxidative defenses,14 and rhamnazin also has been known to have a potent anti-inflammatory activity.15 However, the cellular action underlying its protective properties against inflammation under a murine macrophage RAW264.7 cells have not been well discovered. Moreover, the anti-oxidative effects of rhamnazin were not thoroughly elucidated in murine macrophage RAW264.7 cells.
In the present study, evidence was gathered that rhamnazin may exert its protective role against LPS-induced inflammation by regulating cellular oxidative insults in macrophages. To this end, the protective property of rhamnazin on cell viability in murine macrophage RAW264.7 cells was first determined. To examine the status of anti-inflammatory and anti-oxidative conditions, levels of crucial inflammatory markers and oxidative indices including total RS, •O2, NO•, ONOO–, lipid peroxidation, catalase activity, pro-inflammatory COX as well as 5-LO activities were assessed.
MATERIALS AND METHODS
Chemicals and reagents
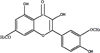
Rhamnazin (see the chemical structure in Fig. 1), 2,7-dichlorofluorescein diacetate (DCFH-DA) was purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Dulbecco's modified Eagle medium/nutrient mixture F-12 (DMEM/F-12) was obtained from GIBCO™ (Grand Island, NY, USA), and fetal bovine serum (FBS) was obtained from Life Technologies Inc. (Lenexa, KS, USA). 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) and dihydrorhodamine 123 (DHR 123) were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA), and all other reagents were of the highest commercial grade available.
Cell culture
RAW264.7 cells, a macrophage like cell line, were purchased from the American Type Culture Collection (Rockville, MD, USA). The cells were cultivated in DMEM supplemented with 10% FBS, at 37℃ in a humidified atmosphere of 5% CO2 in air. Treatment with rhamnazin and / or LPS (100 ng/mL) was carried out under serum-free conditions. All subsequent procedures were carried out under these conditions, and all the experiments were carried out in triplicate and repeated three times to ensure reproducibility.
Assessment of total RS production
Total RS generation was determined in culture supernatant according to the method described by Paraidathathu et al.18 In short, following rhamnazin treatment for 24 h, 25 µM DCFH-DA was added to the cells that were incubated at 37℃ for 15 min, and changes in fluorescence were determined for 30 min at an excitation wavelength of 486 nm and emission wavelength of 530 nm. N-acetyl-L-cystein (NAC, 10 µM) was used as a positive control.
Assay to determine ONOO– level
ONOO–-dependent oxidation of DHR 123 to rhodamine 123 was assayed, according to the method described by Kooy et al.19 Samples were added to the rhodamine buffer (pH 7.4) consisting of 6.25 µM DHR 123 and 125 µM diethylenetriaminepentaacetic acid and incubated 5 min at 37℃. The absorbance was assessed at 500 nm, which is the absorbance of rhodamine 123. Trolox (10 µM), curcumin (10 µM) and penicillamine (10 µM) were used as positive controls.
Assay of NO•
NO• levels were estimated by assessing the accumulation of nitrite in the conditioned medium by the Griess assay. Shortly after, 100 mL culture supernatant was allowed to react with 100 mL of the Griess reagent,20 and after that, it was incubated at room temperature for 5 min. The optical density of the samples at 540 nm was determined using a microplate reader. Trolox (10 µM), curcumin (10 µM) and penicillamine (10 µM) were used as positive controls.
Assessment of •O2
The •O2-scavenging activity was estimated by assessing the reduction in the ratio of the decrease of nitro blue tetrazolium (NBT), as described by Ewing and Janero.21 The culture supernatant was added to the reaction buffer [50 µM PBS with 125 µM EDTA, 62 µM NBT and 98 mM NADH] containing 33 µm 5-methylphenazium methyl sulfate. The absorbance at 540 nm, as an index of NBT reduction, was determined after 5 min. Trolox (10 µM), curcumin (10 µM) and penicillamine (10 µM) were used as positive controls.
Evaluation of lipid peroxidation by thiobarbituric acid reactive substances (TBARS)
To estimate the lipid peroxidation, the levels of TBARS was determined using slightly modified assay according to Buege and Aust.22 This method entailed the creation of a colored complex between the products of lipid peroxidation and thiobarbituric acid at a temperature of 100℃ in an acidic environment. The maximum absorption of this complex was estimated at a wavelength of 532 nm. NAC (10 µM) was used as a positive control.
Estimation of catalase
Cells were cultivated in 100-mm culture dishes containing DMEM supplemented with 10% FBS in a humidified atmosphere of 5% CO2 in air at 37℃. The cells were detached from the dish by 0.05% v/v trypsin-EDTA in PBS and subcultured in culture plates. After incubation for 24 h, cells were treated with rhmanazin, and then the cells were collected and sonicated for 3 min at 30-s intervals. Cell debris was pelleted, and the resulting supernatant was used for the enzyme activity assays. In short, catalase activity was assessed spectrophotometrically by direct estimation of the reduction of light absorption at 240 nm due to the decomposition of H2O2 by catalase.23 Catalase activity was calculated as nmole/min/g protein, and the enzyme activities were calculated by normalizing the results to the total protein concentration of the whole extract, as estimated by the bicinchoninic acid protein assay23 with NAC, (10 µM) as a positive control.
Assessment of COX and 5-LO activity
PGE2 and LTB4 levels were evaluated according to the manufacturer's instructions (R&D Systems, Minneapolis, MN, USA). Cells were transferred to a 48-well plate and allowed to adhere overnight. Cells were incubated for each time in an FBS-free medium in the presence or absence of rhamnazin, and the PGE2 and LTB4 levels present in the media were assayed utilizing an enzyme immunoassay kit.
Statistical analysis
All the experiments were performed in triplicate and repeated three times to ensure reproducibility. Data are presented as mean ± SEM. The effect of each parameter was assessed by the one-way analysis of variance. Individual differences between groups were determined using the Dunnett's test, and those at p < 0.05 were considered statistically significant.
RESULTS
Effect of rhamnazin on cell viability
To determine the protective effect of rhamnazin against LPS-induced cellular damage, cell viability was examined. As Fig. 1 shows, the LPS-treatment decreased cell viability. However, the rhamnazin treatment boosted cell proliferation, indicating that rhamnazin has a protective effect against LPS-induced cell damage.
Rhamnazin suppressed total RS production
As shown in MTT assay, rhamnazin showed cytoprotective effect. In order to investigate the underlying action of cytoprotective action by rhamnazin, total RS production was examined. As shown in Table 1, total RS generation was up-regulated after treatment with LPS, while the rhamnazin treatment decreased total RS generation.
Suppression of ONOO–, NO• and •O2 levels by rhamnazin
To investigate the contributing factors of the cytoprotective effect against LPS exposure in greater detail, we estimated various RS, including ONOO– , NO• and •O2, and found that rhamnazin reduced · ONOO–, NO• and •O2 levels (Table 2) in LPS-treated macrophages. These results indicate that various scavenging activities by rhamnazin are one of the potent, contributing underlying actions that protect against LPS-induced cellular damage.
Rhamnazin reduced lipid peroxidation levels
As Fig. 3A shows, LPS enhanced TBARS levels, while rhamnazin reduced TBARS levels. These data confirm that rhamnazin achieves its anti-oxidative properties by not only scavenging various RS such as •O2, NO•, and ONOO–, but also by suppressing lipid peroxidation.
Rhamnazin boosted catalase activity
To determine the effect of rhamnazin on the regulation of an antioxidant enzyme during LPS-induced oxidative insult, catalase activity was assessed. As shown in Fig. 3B, catalase activity decreased after treatment with LPS. However, rhamnazin treatment enhanced catalase activity. These data indicate that rhamnazin possesses a protective effect against LPS-induced oxidative insult by up-regulating antioxidant enzyme activity.
Rhamnazin modulated COX and 5-LO activities
To examine whether rhamnazin's cytoprotective property is associated with the modulation of inflammation, key pro-inflammatory enzymes, COX and 5-LO activities were estimated. As Fig. 3C and Fig. 3D display, rhamnazin reversed both COX and 5-LO activities with LPS treatment. These results indicate that rhamnazin suppresses inflammation through inhibition of both COX and 5-LO, implying that the dual modulation of COX and 5-LO might be a crucial underlying action in rhamnazin's protection against LPS-induced cytotoxicity.
DISCUSSION
Recently, rhamnazin is known to posses in vitro anti-inflammatory activity acts as inhibitors against dihydroorotate dehydrogenase,24 and rhamnazin from Callicarpa kwangtungensis Chun possessed antioxidant property.14 More recently, rhamnazin from Caragana leucophloea Pojark revealed to have strong antioxidant capacity.25 The present work was undertaken to establish the beneficial effects of rhamnazin on cellular insults, particularly its anti-inflammatory property in LPS-treated macrophage cells. To the best of my knowledge, the current study is the first report showing that rhamnazin possesses cytoprotective effects in its ability to protect the macrophage cells against LPS induced inflammation and ROS/RNS related oxidative status.
The initial approach was to assess the protective effect of rhamnazin on cell proliferation in LPS-exposed macrophages, finding that rhamnazin ameliorated LPS-induced cytotoxicity as shown in Fig. 2. To find the possible underlying action by which rhamnazin acts, several key oxidative markers were assessed and obtained evidence showing that rhamnazin can reduce total RS production (Table 1). These data are parallel with a previous study,26 indicating that an important underlying action against LPS-induced cytotoxicity by rhamnazin is the modulation of total RS generation.
To investigate the anti-oxidative actions of rhamnazin on various RS, its effect on •O2, NO•, and ONOO– levels were monitored. As shown in Table 2, the results reveal that rhamnazin has broad RS scavenging actions that simultaneously ameliorate •O2, NO•, and ONOO–, indicating that these potent and broad RS-suppressing properties are significant contributing factors in rhamnazin's cytoprotective effect against LPS.
Because oxidative damage due to RS is known to cause lipid peroxidation in the cell membrane,27 the ability of rhamnazin to suppress lipid peroxidation was also evaluated. As expected, LPS-mediated lipid peroxidation was effectively suppressed by rhamnazin (Fig. 3A).
One intriguing and important piece of evidence obtained in the present study is rhamnazin's up-regulation of catalase activity, which may play an important role in protecting various cellular components.
We previously reported that oxidative damage caused by excessive ROS/RNS and lipid peroxidation is casually connected with inflammation.3 As enhanced PG biosynthesis catalyzed by COX is a significant and important source of RS, and COX is a crucial enzyme in the inflammatory process,28 the action of rhamnazin on COX activity was assessed. As Fig. 3C shows, rhamnazin suppressed COX activity. Also, because 5-LO is another major pro-inflammatory enzyme,5 5-LO activity was examined. The results show that rhamnazin inhibited 5-LO activity, (Fig. 3D) indicating that controlling 5-LO activity is also a key underlying action that counteracts the cytotoxicity induced by LPS.
Chronic inflammatory disease is known to possess various causative actions, and one such action is oxidative damage from RS such as ROS/RNS and lipid peroxidation. LPS has been experimentally used as an macrophage activator and an oxidative stressor,29 and the LPS treated macrophage cell model has been utilized for investigating chronic inflammatory disease30 and oxidative stress related.31
One major finding from this study is the inhibition of both COX and 5-LO by rhamnazin, thereby showing a crucial underlying action in the protection against LPS-induced inflammation, and therefore, it is highly likely that rhamnazin possesses anti-inflammatory properties through the dual inhibition of COX and 5-LO. Thus, novel and interesting findings from this current work are the multifarious protective effects of rhamnazin, including those defending against pro-inflammatory COX and 5-LO.
Other significant findings from this study include the broad efficacies of rhamnazin with respect to its anti-inflammatory and anti-oxidative actions on total RS production, lipid peroxidation, •O2, NO•, ONOO–, enhanced catalase activity, and against pro-inflammatory COX-2 and 5-LO activities. It is important to highlight that rhamnazin may protect cells directly not only by scavenging various ROS/RNS through its anti-oxidative action, but also by boosting antioxidant enzymes, like catalase, and anti-inflammatory actions in LPS treated macrophage cells.
SUMMARY
The results from current study demonstrate that rhamnazin protects against LPS-induced inflammation and ROS/RNS in macrophage cells. The plausible underlying protective action might be rhamnazin's ability to possess anti-inflammatory and anti-oxidative properties through the modulation of NO•, ONOO–, pro-inflammatory COX-2 and 5-LO activities, total RS generation, •O2, lipid peroxidation and catalase activity.




 XML Download
XML Download